
Figure 8.
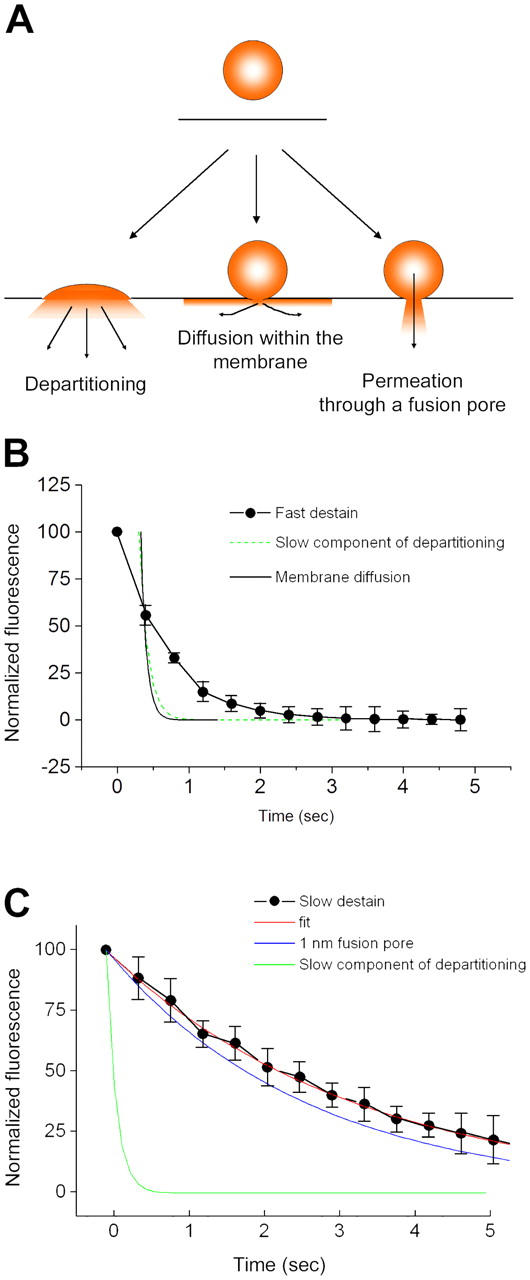
Mechanisms of FM1-43 efflux from exocytosing SVs. (A) Schematic diagram illustrating possible modes by which FM1-43 might leave SVs during exocytosis. If vesicles fully collapse into the plasma membrane, FM1-43 could escape by direct departitioning into the bulk solution. Alternatively, dye could escape through even a tiny fusion pore if the pore was lipidic and allowed mixing of vesicular and plasma membrane components, as a result of diffusion within the bilayer. The final possibility is that a small aqueous pore opens, the constituents of which form a barrier to diffusion of FM1-43 within the membrane. (B) Examination of the kinetics of fast destaining events, using the averaged values from Fig. 5 D. Because the fastest events occur at a rate similar to the sampling frequency, we cannot be certain when the drop in fluorescence actually begins. Consequently, the data do not fall all that far from a τ of ∼120 ms, which is the rate of the slow component of departitioning we observed in our stopped-flow experiments. As a result of the limitations of the imaging frequency, we cannot readily distinguish between FM1-43 leaving vesicles by diffusion within the membrane and leaving by direct departitioning. (C) Examination of the kinetics of slow destaining events. The averaged curves are replotted from Fig. 5 D. The slow component of membrane departitioning is much too slow to account for the kinetics by which FM1-43 escapes from vesicles. Instead, we have compared the rate of efflux of FM1-43 with simple diffusion-limited permeation through an aqueous pore with an opening of 1 nm. There is a fairly close agreement between this and the experimental data, although this model does not take into account any electrostatic interactions or geometric constraints, which might tend to slow the dye efflux. Error bars are SEM.