

Abstract
Endoplasmic reticulum (ER)-associated degradation (ERAD) is the process by which aberrant proteins in the ER lumen are exported back to the cytosol and degraded by the proteasome. Although ER molecular chaperones are required for ERAD, their specific role(s) in this process have been ill defined. To understand how one group of interacting lumenal chaperones facilitates ERAD, the fates of pro–α-factor and a mutant form of carboxypeptidase Y were examined both in vivo and in vitro. We found that these ERAD substrates are stabilized and aggregate in the ER at elevated temperatures when BiP, the lumenal Hsp70 molecular chaperone, is mutated, or when the genes encoding the J domain–containing proteins Jem1p and Scj1p are deleted. In contrast, deletion of JEM1 and SCJ1 had little effect on the ERAD of a membrane protein. These results suggest that one role of the BiP, Jem1p, and Scj1p chaperones is to maintain lumenal ERAD substrates in a retrotranslocation-competent state.
Keywords: Jem1p, BiP, ERAD, molecular chaperone, protein translocation
Introduction
Newly synthesized secretory proteins translocate into the ER, and a quality control system ensures that only correctly folded and/or assembled proteins are transported from the ER to their ultimate destinations. However, misfolded or unassembled multimeric proteins are retained in the ER and may aggregate or be degraded (Ellgaard et al. 1999). A process that we termed ER-associated protein degradation (ERAD) (McCracken and Brodsky 1996; Werner et al. 1996) plays a central role in quality control by selectively degrading terminally misfolded or unassembled proteins (for review see Brodsky and McCracken 1999; Plemper and Wolf 1999). The ERAD of many soluble and membrane proteins requires the 26S proteasome located in the cytosol, suggesting that these proteins are retrotranslocated from the ER to the cytosol. In fact, much evidence indicates that the protein translocation channel, composed of the Sec61p complex in the ER membrane, mediates the retrotranslocation of ERAD substrates (for review see Römisch 1999). This hypothesis was first proposed because an ERAD substrate in mammalian cells coprecipitates with Sec61p (Wiertz et al. 1996). In accordance with this result, CPY*, a mutated form of vacuolar carboxypeptidase Y (CPY) that is degraded by ERAD (Hiller et al. 1996), was stabilized in sec61 mutant yeast (Plemper et al. 1997), and the degradation of ΔGpαF, an unglycosylated mutant form of the yeast α mating pheromone pro–α-factor (pαF), was inhibited in microsomes prepared from sec61 strains (Pilon et al. 1997, Pilon et al. 1998). Strains containing mutant alleles of SEC61 that are proficient for protein import but are defective in the retrotranslocation process were isolated (Zhou and Schekman 1999; Wilkinson et al. 2000), suggesting that these processes are mechanistically distinct.
BiP, an Hsp70 molecular chaperone in the yeast ER lumen, has also been shown to facilitate ERAD. BiP recognizes unfolded proteins and facilitates protein folding in the yeast ER (Simons et al. 1995). Stabilization of soluble ERAD substrates, such as CPY* and a variant of the α1-protease inhibitor, A1PiZ, was observed in yeast cells containing temperature-sensitive mutations in KAR2, the gene encoding yeast BiP, and the degradation of ΔGpαF was inhibited in microsomes prepared from temperature-sensitive kar2 mutants (Plemper et al. 1997; Brodsky et al. 1999). BiP also drives posttranslational import into the ER by acting as a molecular ratchet (Matlack et al. 1999). Because some temperature-sensitive kar2 mutants that are proficient for import show ERAD defects, the role of BiP in ERAD differs from its role in protein import (Brodsky et al. 1999). However, the molecular mechanisms underlying the function of BiP in ERAD are not clear.
Interestingly, the yeast ER contains a chaperone that is ∼24% identical to BiP, known variably as Lhs1p (Craven et al. 1996), Cer1p (Hamilton and Flynn 1996), or Ssi1p (Baxter et al. 1996). The translocation of some preproteins in cells lacking LHS1 is impaired, and genetic interactions have been observed between this chaperone and BiP. Although a role for Lhs1p in stabilizing and refolding thermally denatured proteins in the ER has been demonstrated (Saris et al. 1997), it is unknown whether this chaperone plays a role in ERAD.
DnaJ-like proteins regulate the ATP-dependent reaction cycle of Hsp70 by interacting with Hsp70 via their conserved J domains (Kelley 1998). The yeast ER contains three DnaJ-like proteins: Sec63p, Jem1p, and Scj1p (Sadler et al. 1989; Schlenstedt et al. 1995; Nishikawa and Endo 1997). Sec63p is an essential membrane protein and interacts with BiP to drive the translocation of proteins into the ER lumen (Rothblatt et al. 1989; Brodsky and Schekman 1993; Lyman and Schekman 1995; Corsi and Schekman 1997; Matlack et al. 1999). Jem1p and Scj1p are ER lumenal proteins that are not essential for cell viability, but cells lacking both Jem1p and Scj1p are temperature sensitive, suggesting that their functions overlap (Nishikawa and Endo 1997). The Δjem1Δscj1 strain is proficient for protein import into the ER, but is defective for the transport of CPY*. Because the quality control system prevents the transport of defective proteins through the secretory pathway, Jem1p and Scj1p are likely involved in protein folding and assembly in the ER lumen (Silberstein et al. 1998). Scj1p interacts with BiP via its J domain (Schlenstedt et al. 1995) and in cooperation with BiP, Jem1p mediates nuclear membrane fusion during mating (Nishikawa and Endo 1997; Brizzio et al. 1999). Thus, BiP may engage each of the three J domain–containing chaperones in the yeast ER to drive several cellular processes.
Because DnaJ-like proteins regulate Hsp70s, it is natural to suspect that Sec63p, Jem1p, and Scj1p are involved in ERAD. Although an increase in the half-life of CPY* in the ER was observed using a sec63-1 mutant, this effect is marginal when compared with the defect observed in sec61 mutants (Plemper et al. 1997). Similar results were observed for the in vitro degradation of ΔGpαF using microsomes prepared from the sec63-1 mutant (Pilon et al. 1997).
In this study, we report that Jem1p and Scj1p, the soluble DnaJ-like proteins in the ER lumen, are involved in the ERAD of CPY* and ΔGpαF. CPY* and ΔGpαF aggregate in a temperature-sensitive kar2 mutant and the Δjem1Δscj1 strain at elevated temperature, and once aggregation occurs, the ERAD of these proteins is significantly impaired. We propose that BiP cooperates with the soluble J domain proteins in the ER, Jem1p and Scj1p, to facilitate the export of ERAD substrates to the cytosol by preventing their aggregation in the ER lumen.
Materials and Methods
Yeast Strains and Culture Conditions
Yeast strains used in this study were constructed from SEY6210 (MATα ura3 leu2 trp1 his3 lys2 suc2) (a gift from S. Emr, University of California, San Diego, San Diego, CA; Robinson et al. 1988), MS10 (MATα ura3 leu2 ade2; a gift from M. Rose (Princeton University, Princeton, NJ), RSY579 (MATα kar2-159 ura3 leu2 trp1 ade2) (a gift from R. Schekman, University of Calfornia, Berkeley, Berkeley, CA; Sanders et al. 1992), RDM15-5B (MATα sec61-2 ura3 leu2 ade2 pep4) (a gift ofrom R. Schekman; Deshaies and Schekman 1987), JN516 (MATα SSA1 ssa2-1 ssa3-1 ssa4-1 his3 leu2 ura3 trp1 lys2), or a1-45Δu (MATα ssa1-45 ssa2-1 ssa3-1 ssa4-1 his3 leu2 ura3 trp1 lys2) (Becker et al. 1996). SNY1084-1D (MATα ura3 leu2 ade2) was constructed by crossing MS10 and RSY579. To construct the prc1-1 mutant, a 3.7-kb EcoRI-SalI fragment of pTSY1000 (a gift from T.M. Stevens, Univeristy of Oregon, Eugene, OR; Stevens et al. 1986) containing the PRC1 gene was subcloned into the EcoRI and SalI sites of pJJ244 (Jones and Prakash 1990). The prc1-1 mutation was introduced by oligonucleotide-directed mutagenesis (Kunkel et al. 1987) us-ing an oligonucleotide 5′-CCACATCGCTAGGGAATCCTAC-3′. The sec63-1 and sec61-2 mutant genes were cloned by PCR using the genomic DNA of RSY151 (MATα sec63-1 ura3 leu2 pep4) (a gift from R. Schekman; Rothblatt et al. 1989) and RDM15-5B as a template, respectively. The amplified DNA fragments were introduced into the polylinker site of pRS306 (Sikorski and Hieter 1989). The resulting plasmids were linearized and used for introduction of the prc1-1, the sec63-1, or the sec61-2 allele by the two step gene replacement method (Rose et al. 1990). Introduction of the mutant alleles into the yeast genome was confirmed by DNA sequencing. The Δjem1::LEU2 and Δscj1::TRP1 alleles were constructed as described previously (Nishikawa and Endo 1997). The Δscj1::URA3 gene was constructed by replacing the 0.84-kb MunI-XbaI fragment of pBS-SCJ1 (Nishikawa and Endo 1997) with the 1.2-kb HindIII fragment of pJJ242 containing the URA3 gene. The LHS1 gene was cloned by PCR using primers 5′-GCGCTCGAGCGCCATTGAATTTAGAAC-3′ and 5′-GCGGAGCTCGCCAAAGGAGACAGGTGG-3′, and the amplified DNA fragment was introduced into the XhoI and SacI sites of pBluescriptII SK+ to give pBSLHS1. The Δlhs1::TRP1 allele was constructed by replacing the 2.5-kb EcoRV fragment of pBSLHS1 with the 0.85-kb SmaI-StuI fragment of pJJ281 (Jones and Prakash 1990) containing the TRP1 gene. The DOA4 gene was cloned by PCR using primers 5′-CGCGAGCTCCATAAGGGATGAGTGTA-3′ and 5′-GCGCTCGAGCAAACACCGTAGACGCGG-3′, and the amplified DNA fragment was introduced into the XhoI and the SacI sites of pBluescriptII SK+ to give pBSDOA4. The Δdoa4::LEU2 allele was constructed by replacing the 1-kb BglII-SalI fragment of pBSDOA4 (Papa and Hochstrasser 1993) with the 2-kb BamHI-SalI fragment of pJJ282 (Jones and Prakash 1990) containing the LEU2 gene. A 3.5-kb DNA fragment containing the Δerg6::LEU2 gene was amplified from the genomic DNA of KUY161 (MATα erg6::LEU2 ura3 lys2 ade2 his3 leu2) (a gift from K. Umebayashi, RIKEN, Wako, Japan) by PCR using primers 5′-AATCAACCAAACAGCTAGGGCTGG-3′ and 5′-AAGGCCTGCTAGCAATGAACGTGC-3′. Null alleles were introduced into yeast cells by the one step gene disruption method (Rose et al. 1990). Multiple mutants were generated by a second round of transformation. The constructed strains were: KYSC1 (MATα prc1-1 ura3 leu2 trp1 his3 lys2 suc2), KYSC2 (MATα prc1-1 Δjem1::LEU2 ura3 leu2 trp1 his3 lys2 suc2), KYSC3 (MATα prc1-1 Δscj1::TRP1 ura3 leu2 trp1 his3 lys2 suc2), KYSC4 (MATα prc1-1 Δjem1::LEU2 Δscj1::TRP1 ura3 leu2 trp1 his3 lys2 suc2), KYSC5 (MATα prc1-1 sec63-1 ura3 leu2 trp1 his3 lys2 suc2), KYSC7 (MATα sec63-1 ura3 leu2 trp1 his3 lys2 suc2), KYSC11 (MATα sec61-2 Δjem1::LEU2 ura3 leu2 ade2 pep4), KYSC13 (MATα prc1-1 kar2-159 ura3 leu2 trp1 ade2), KYSC14 (MATα prc1-1 Δlhs1::TRP1 ura3 leu2 trp1 his3 lys2 suc2), SNY1026-7A (MAT α Δjem1::LEU2 Δscj1::TRP1 ura3 leu2 trp1 his3 lys2 suc2), SNY1070 (MATα sec61-2 Δscj1::URA3 ura3 leu2 ade2 pep4), SNY1071 (MATα sec61-2 Δjem1::LEU2 Δscj1::URA3 ura3 leu2 ade2 pep4), SNY1077 (MATα sec61-2 SSA1 ssa2-1 ssa3-1 ssa4-1 his3 leu2 ura3 trp1 lys2), SNY1078 (MATα sec61-2 ssa1-45 ssa2-1 ssa3-1 ssa4-1 his3 leu2 ura3 trp1 lys2), SNY1081 (MATα erg6::LEU2 sec61-2 ura3 leu2 ade2 pep4), SNY1082 (MATα doa4::LEU2 sec61-2 ura3 leu2 ade2 pep4), and SNY1083 (MATα prc1-1 ura3 leu2 ade2). BJ3505 (MATα pep4::HIS3 prb1 lys2 trp1 ura3 gal2 can1) (a gift from Y. Onya, University of Tokyo, Tokyo, Japan) was used for cytsol preparation. Cells were grown in YPD medium containing 1% yeast extract, 2% polypeptone, and 2% glucose. A sulfate-free synthetic minimal medium (Rothblatt and Schekman 1989) was used for metabolic labeling of yeast cells.
Pulse–Chase Experiments
Metabolic labeling of yeast cells with Tran35S-label (ICN Biomedicals) and preparation of cell extracts were performed as described previously (Nishikawa and Nakano 1991). When indicated, carbobenzoxyl-leucinyl-leucinyl-leucinal (MG132; Calbiochem) was added to the labeling medium at 200 μM for 60 min before the labeling. For the pulse–chase analysis of pαF, cells were pretreated with tunicamycin (10 μg/ml) for 20 min at 23°C and pulse labeled in the presence of tunicamycin. Immunoprecipitation with anti-CPY or anti–prepro–α-factor (ppαF) antiserum (gifts from A Nakano, RIKEN, Wako, Japan; Jones 1991) was performed as described by Nishikawa et al. 1994. In the case of immunoprecipitation using an anti-Sec61p antiserum, cell extracts were prepared by incubating at 37°C for 30 min instead of 100°C, and 2.5 μl of anti-Sec61p antiserum (Stirling et al. 1992) was added to the cell extracts derived from 0.5 OD600 cells.
Sucrose Density Gradient Centrifugation
60 OD600 units of cells were collected by centrifugation and washed once with ice cold 10 mM NaN3. Cells were suspended in 600 μl of 20 mM Hepes-KOH, pH 7.4, 50 mM potassium acetate, 2 mM EDTA, 1 mM PMSF, 5 μg/ml pepstatin A, 2 μg/ml chymostatin, 5 μg/ml E-62, 100 μg/ml N-tosyl-l-phenylmethylchloromethyl ketone, 200 μg/ml p-aminobenzamidine hydrochloride, 1 mM ε-amino capronic acid, 2 μg/ml aprotinin, 2 μg/ml antipain, 2 μg/ml leupeptin, and 20 μg/ml 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochloride and disrupted by agitation with glass beads. After removing cell debris by centrifugation, 1% Triton X-100 was added to the cell lysate, which was subsequently centrifuged at 12,000 g for 10 min at 4°C. The cleared lysate was layered onto linear sucrose gradients (4 ml, 5–40% sucrose in 20 mM Hepes-KOH, pH 7.4, 50 mM potassium acetate, 2 mM EDTA, 0.1% Triton X-100, 1 mM PMSF, 5 μg/ml pepstatin A, 2 μg/ml chymostatin, 5 μg/ml E-62, 100 μg/ml N-tosyl-l-phenylmethylchrolomethyl ketone, 200 μg/ml p-aminobenzamidine hydrochloride, 1 mM ε-amino capronic acid, 2 μg/ml aprotinin, 2 μg/ml antipain, 2 μg/ml leupeptin, and 20 μg/ml 4-(2-aminoethyl) benzenesulfonyl fluoride hydrochrolide), and centrifuged at 145,000 g for 20 h at 4°C. After centrifugation, fractions were collected from the bottom and analyzed by immunoblotting using anti-CPY antiserum.
In Vitro Translocation and ERAD Assays
Preparations of the yeast microsome and cytosol fractions were performed as described previously (McCracken and Brodsky 1996). 35S–labeled ppαF and ΔGppαF were prepared in vitro as described previously, and purified over a Sephadex G-25 column (McCracken and Brodsky 1996; Werner et al. 1996). ER-derived microsomes were incubated with 35S–labeled ppαF or ΔGppαF in the presence of an ATP-regenerating system. Microsomes containing GpαF and ΔGpαF were collected by centrifugation, washed once, and resuspended in buffer containing an ATP-regenerating system. Microsomes were preincubated at 23 or 37°C for the indicated duration in the absence of the cytosol, but in the presence of ATP. For the analysis of ΔGpαF degradation, preincubated microsomes were further incubated in the presence of an ATP-regenerating system and cytosol at 37°C. At the indicated time points, proteins were precipitated with trichloroacetic acid and subjected to SDS-PAGE with 18% polyacrylamide gel containing SDS and 4 M urea. For the analysis by sucrose density gradient centrifugation, proteins were extracted from the preincubated suspension by 1% Triton X-100, and after centrifugation at 12,000 g for 10 min at 4°C, cleared extracts were subjected to sucrose density gradient centrifugation as described above. After centrifugation, fractions were collected from the bottom and analyzed by SDS-PAGE after trichloroacetic acid precipitation.
Results
Jem1p and Scj1p, but Not Sec63p, Cooperate with BiP in the ERAD of CPY*
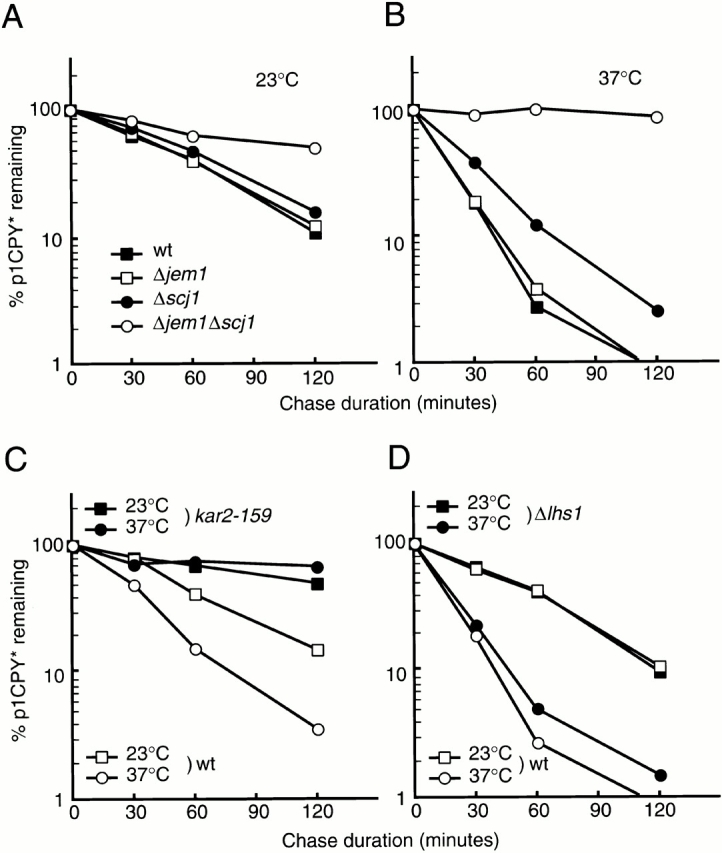
Several lines of evidence show that Jem1p, Scj1p, and Sec63p interact with BiP through their J domains (Brodsky and Schekman 1993; Scidmore et al. 1993; Schlenstedt et al. 1995; Corsi and Schekman 1997; Nishikawa and Endo 1997). To reveal which, if any, of these J domain–containing proteins are involved in ERAD, we tested the effects of functional defects in these proteins on the degradation of CPY*. Although CPY is a vacuole-targeted protein, CPY* does not reach the vacuole but, instead, is exported from the ER to the cytoplasm and rapidly degraded by the proteasome (Finger et al. 1993; Hiller et al. 1996). When we examined the fate of CPY* in a wild-type strain by pulse–chase analysis, CPY* was found to be degraded with half-lives of 43 min at 23°C, and 12 min at 37°C (Fig. 1A and Fig. B, filled squares). The half-life of CPY* increased 2.7-fold at 23°C in temperature-sensitive BiP (kar2-159) mutant cells (Fig. 1 C), suggesting BiP-dependent export of lumenal CPY* to the cytosol, as observed previously (Plemper et al. 1997). This defect was exacerbated when kar2-159 mutant cells were metabolically labeled at 23°C for 10 min and subsequently chased at 37°C. In contrast, cells containing a disruption of the LHS1 gene degraded CPY* at similar rates to those in wild-type cells at 23 and 37°C (Fig. 1 D). To confirm this result, we measured the degradation of ΔGpαF, another soluble ERAD substrate (McCracken and Brodsky 1996) in microsomes prepared from wild-type or the LHS1-deleted strain and found that ERAD was unaffected (data not shown). These results indicate that BiP, but not Lhs1p, is involved in the degradation of CPY* in the ER lumen.
Figure 1.
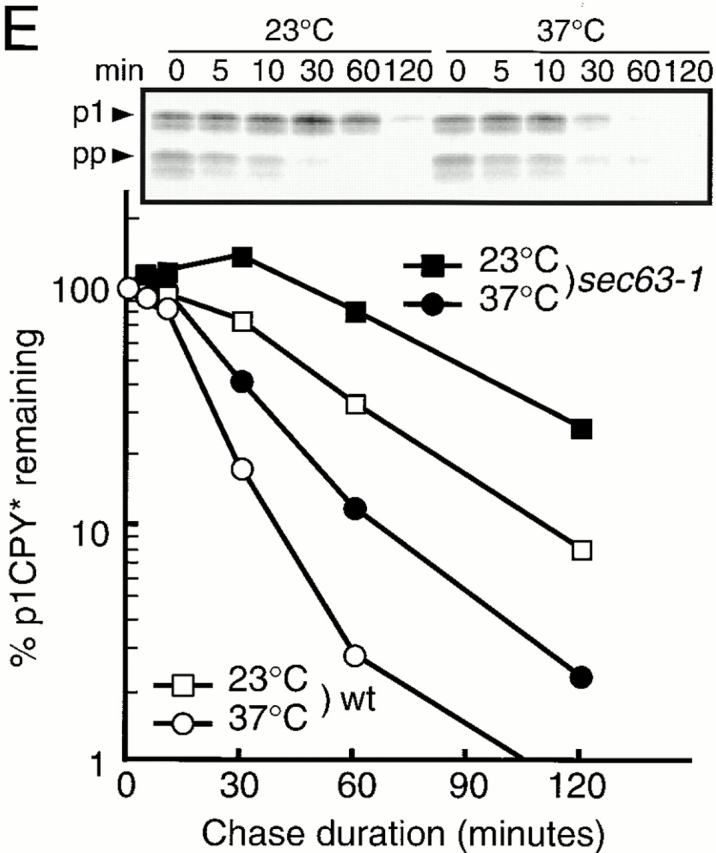
Degradation of CPY* in ER chaperone mutants. Wild-type (KYSC1, wt), Δjem1 (KYSC2, Δjem1), Δscj1 (KYSC3, Δscj1), and Δjem1 Δscj1 (KYSC4, Δjem1 Δscj1) cells were pulse labeled with 35S–amino acids for 10 min and chased for the indicated times either at 23°C (A) or at 37°C (B). CPY* was recovered from cell extracts by immunoprecipitation using an anti-CPY antiserum and analyzed by SDS-PAGE. Relative amounts of CPY* were quantified by radioimaging, and the means of two independent experiments are shown. The amount of CPY* at 0 min chase was set to 100%. ▪, wild-type; □, Δjem1; •, Δscj1; ○, Δjem1 Δscj1. (C) kar2-159 (KYSC13, kar2-159) and wild-type (SNY1083, wt) cells were pulse labeled with 35S–amino acids for 10 min at 23°C and chased for the times indicated at 23°C or at 37°C. Degradation of CPY* was followed as in A and B. ▪, kar2-159 chased at 23°C; •, kar2-159 chased at 37°C; □, wild-type chased at 23°C; ○, wild-type chased at 37°C. (D) Δlhs1 (KYSC14) cells and wild-type (KYSC1, wt) cells were pulse labeled with 35S–amino acids for 10 min and chased for the times indicated either at 23 or 37°C. Degradation of CPY* was followed as in A and B. ▪, Δlhs + at 23°C; •, Δlhs1 at 37°C; □, wild-type at 23°C; ○, wild-type at 37°C. (E) sec63-1 (KYSC5, sec63-1) and wild-type (KYSC1, wt) cells were pulse labeled with 35S–amino acids for 10 min at 23°C and chased for the indicated times at 23 or 37°C. Degradation of CPY* was followed as in A and B. ▪, sec63-1 chased at 23°C; •, sec63-1 chased at 37°C; □, wild-type at 23°C; ○, wild-type at 37°C; pp, prepro-CPY*; p1, p1CPY*.
Disruption of one of the two genes for the ER lumenal DnaJ-like proteins, Jem1p and Scj1p, did not lead to profound stabilization of CPY* at 23 or 37°C. However, simultaneous disruption of the genes encoding Jem1p and Scj1p stabilized CPY* significantly, so that the half-life of CPY* increased 2.6-fold at 23°C, and only ∼10% of the CPY* was degraded over a 120-min chase at 37°C (Fig. 1A and Fig. B, open circles). Because a precursor form of CPY* was not observed, translocation of CPY* was not delayed in the Δjem1Δscj1 mutant cells in these experiments (data not shown). To reexamine the role of Sec63p in ERAD, we measured the stability of CPY* in sec63-1 mutant cells (Fig. 1 E). The sec63-1 mutant strain exhibited a moderate defect in protein import into the ER, even at 23°C, because a precursor form of CPY* (prepro-CPY*) transiently accumulated at early time points during the chase. However, the level of CPY* in the ER lumen (p1 form) subsequently declined with half-lives of 38 min at 23°C and 17 min at 37°C. These half-lives of lumenal CPY* do not differ significantly from those in wild-type cells (see above). Thus, the 1.5-fold retardation in the rate of CPY* proteolysis in the sec63-1 strain that was previously reported (Plemper et al. 1997) was not apparent in these experiments, perhaps because the genetic backgrounds of the strains used in these studies differ. From these data, we conclude that two of the three ER DnaJ-like proteins, Jem1p and Scj1p, are involved in the ERAD of CPY*.
Defects in BiP or Jem1p and Scj1p Cause Aggregation of CPY* in the ER
It has been established that Hsp70 binds to and dissociates from unfolded proteins concomitant with ATP binding and hydrolysis, and thereby prevents substrate proteins from aggregating; J domain–containing proteins impact upon this cycle by stimulating ATP hydrolysis (Liberek et al. 1991). The aggregation or formation of high molecular weight oligomers of ERAD substrates in the lumen may hinder retrotranslocation to the cytoplasm. Therefore, we decided to test if defects in the functions of BiP, Jem1p, and Scj1p promote the aggregation of CPY* when ERAD is hindered. Wild-type, Δjem1Δscj1, and kar2-159 cells grown at 23°C were incubated at 37°C for 60 min, and proteins were extracted from the cells with 1% Triton X-100 and subjected to sucrose density gradient centrifugation. After centrifugation, ∼50% of CPY* in wild-type cells was recovered in the fractions corresponding to 70–150 kD (fractions 7–10), and only 13% was found in the pellet (Fig. 2). In contrast, in Δjem1Δscj1 and kar2-159 cells, the amount of CPY* recovered in fractions 7–10 was reduced, and 30% (in Δjem1Δscj1 cells) and 60% (in kar2-159 cells) of CPY* were recovered in the pellets, indicating that CPY* aggregates in these strains at 37°C. The distribution patterns of CPY* by sucrose density gradient centrifugation did not differ significantly at 23°C between wild-type cells and kar2-159 or Δjem1Δscj1 cells (data not shown). We suggest that the aggregated CPY* in the Δjem1Δscj1 and in kar2 mutants at 37°C gives rise to the ERAD defect observed in Fig. 1.
Figure 2.

CPY* aggregates in the Δjem1 Δscj1 and the kar2-159 mutants. Wild-type (KYSC1, wt), Δjem1 Δscj1 (KYSC4), and kar2-159 (KYSC13) cells were grown to early logarithmic phase at 23°C and then shifted to 37°C for 60 min. Cell extracts were prepared and subjected to sucrose density gradient centrifugation as described in the Materials and Methods. Fractions were collected and proteins were analyzed by SDS-PAGE followed by immunoblotting using an anti-CPY antiserum. Relative amounts of CPY* were quantified. Numbers indicate fractions (from bottom to top) and P represents the pellet fraction recovered in the bottom of the tube. Vertical arrowheads show the positions of apoferritin (440 kD), alcohol dehydrogenase (150 kD), bovine serum albumin (70 kD), and carbonic anhydrase (30 kD).
BiP and Jem1p/Scj1p Facilitate ERAD of ΔGpαF by Preventing Aggregate Formation in the ER
To confirm these results, we performed in vitro export/degradation assays with microsomes prepared from Δjem1Δscj1, kar2-159, and sec63-1 mutant cells. Radiolabeled ΔGppαF was translocated into microsomes prepared from each of the mutant strains and the isogenic wild-type strain. Upon signal sequence cleavage, the resulting product, ΔGpαF, becomes an ERAD substrate after the washed vesicles are incubated in the presence of cytosol and ATP (McCracken and Brodsky 1996). When this assay was performed using microsomes prepared from the kar2-159, sec63-1, and Δjem1Δscj1 mutant strains, efficient degradation of ΔGpαF was observed at both 23 and 37°C (data not shown). These results demonstrated that ERAD was proficient in the kar2-159 and Δjem1Δscj1 microsomes, in contrast to what was observed using CPY* in vivo (Fig. 1).
To explain this discrepancy, we envisaged that the aggregation of ΔGpαF, and subsequent ERAD defect, might not be apparent unless the translocation and ERAD assays were further uncoupled. Thus, ΔGpαF-loaded microsomes were first preincubated at 37°C for 20 min in the absence of cytosol but in the presence of ATP, and subsequently incubated with both cytosol and ATP. The additional incubation had no effect on ERAD activity in wild-type microsomes (Fig. 3 A, panel a). However, we now observed that the degradation of ΔGpαF in the kar2-159 and Δjem1Δscj1 mutant microsomes was compromised. Only ∼20% of ΔGpαF was degraded after a 60-min chase for both mutant microsomes (Fig. 3 A, panels b and c, filled circles), in contrast to 40–50% of ΔGpαF degraded in wild-type microsomes and in the absence of the additional incubation (open circles). Only a slight decrease was observed when ERAD efficiency was examined at 37°C (filled circles) versus 4°C (open circles) in microsomes prepared from the sec63-1 mutant (Fig. 3 A, panel d). We also found that the extent of the ERAD defect in the kar2-159 and Δscj1Δjem1 microsomes correlated with the length of the incubation at 37°C (Fig. 3 B). In contrast, ERAD activity was robust in the wild-type and sec63-1 microsomes during the same time course at 37°C (Fig. 3 B).
Figure 3.


ERAD defects correlate with aggregation of ΔGpαF. (A) Radiolabeled ΔGppαF was translocated into microsomes prepared from wild-type (a), kar2-159 (b), Δjem1 Δscj1 (c), or sec63-1 (d) strains. After translocation, membranes were incubated on ice (○) or at 37°C (•) for 20 min in the presence of ATP. Cytosol prepared from the wild-type strain was added to a final concentration of 5 mg protein/ml, and posttranslocation chase reactions were performed at 37°C for the indicated times. Proteins were precipitated with trichloroacetic acid and analyzed by SDS-PAGE. Relative amounts of ΔGpαF were quantified by radioimaging. Error bars indicate standard deviation from the means of at least four independent experiments. The amount of ΔGpαF at 0 min chase was set to 100%. (B) Radiolabeled ΔGppαF was translocated into microsomes prepared from wild-type (•), kar2-159 (▪), Δjem1 Δscj1 (□), or sec63-1 (○) strains. Membranes were recovered and incubated at 37°C in the presence of ATP for the indicated times. Cytosol was added to a final concentration of 5 mg protein/ml, and posttranslocation chase reactions were performed at 37°C for 20 min. Degradation activity was represented as the amount of ΔGpαF degraded, and the amount of ΔGpαF with 0 min preincubation was set to 100%. (C) Radiolabeled ΔGppαF and ppαF were translocated into microsomes prepared from wild-type (a and e), kar2-159 (b and f), Δjem1 Δscj1 (c and g), or sec63-1 (d and h) strains. After translocation, membranes were incubated at 23°C (a–d) or 37°C (e–h) for 20 min in the presence of ATP. Membranes were solubilized with 1% Triton X-100, subjected to sucrose density gradient centrifugation, and analyzed as in the legend to Fig. 2. Relative amounts of ΔGpαF and glycosylated pαF were quantified by radioimaging. Black bars, ΔGpαF; white bars, glycosylated pαF.
Next, we tested if ΔGpαF aggregates in kar2-159 and Δjem1Δscj1 mutant microsomes during the 37°C incubation. ΔGppαF and wild-type ppαF were translocated into microsomes prepared from wild-type, kar2-159, Δjem1Δscj1, or sec63-1 strains at 20°C, and microsomes were collected by centrifugation and incubated at 23 or 37°C for 20 min in the presence of ATP. Microsomal protein was solubilized with Triton X-100, analyzed by sucrose density gradient centrifugation, and fractions were examined for the presence of radiolabeled protein by SDS-PAGE and radioimaging. At 23°C in all cases, ΔGpαF showed a broad range distribution around the fraction corresponding to 70 kD (Fig. 3 C, black bars). When microsomes from kar2-159 and Δjem1Δscj1 cells were incubated at 37°C, the amount of ΔGpαF recovered at the bottom of the tube increased significantly. In contrast, the distribution of ΔGpαF in the wild-type or sec63-1 microsomes incubated at 37°C was essentially unchanged from the results obtained when microsomes had been incubated at 23°C. These data indicate that ΔGpαF aggregates at 37°C in kar2-159 and Δjem1Δscj1 microsomes.
When the distribution of wild-type, glycosylated pαF (GpαF) was analyzed under the same conditions, we noted that GpαF was recovered exclusively in fractions corresponding to proteins of 30–70 kD, regardless of the type of microsomes used and whether they had been incubated at 23 or 37°C (Fig. 3 C, white bars). Therefore, aggregate formation due to defects in the functions of BiP, Jem1p, and Scj1p is specific for an ERAD substrate.
In parallel with these in vitro ERAD assays, we followed the degradation of unglycosylated α-factor in the presence of tunicamycin in vivo (Zhou and Schekman 1999). We failed to observe a difference in the degradation rates of ΔGpαF between wild-type cells and Δjem1Δscj1 cells at either 23 or 37°C (Fig. 4 A). We suggest that this arises from the fact that the requirement of Scj1p/Jem1p for the degradation of ΔGpαF in vitro is most evident only after a further uncoupling of the translocation and ERAD processes (Fig. 3). In vivo, the delivery of ΔGpαF from the ER to the cytoplasm and proteasome may be very tightly coupled, obviating the need for these chaperones. In contrast, the degradation of CPY* does require Scj1p/Jem1p (Fig. 1), a phenomenon that may represent the fact that CPY* translocates completely through the pore and into the lumen of the ER before its reexport (Plemper et al. 1999); this may not be the case for ΔGpαF in vivo. On the other hand, we observed a partial stabilization of ΔGpαF in the kar2-159 mutant (Fig. 4 B); after 45 min at 37°C, there was ∼34% of the initial material in the kar2 mutant strain, compared with ∼16% in the wild-type strain. Tunicamycin treatment upregulates the expression of the KAR2 gene, so that the overproduction of the mutant BiP protein may well interfere with the degradation of ΔGpαF by interacting unproductively with ΔGpαF or components required for the ERAD process.
Figure 4.

In vivo ERAD analysis of pαF. (A) Wild-type (SEY6210, wt) and Δjem1 Δscj1 (SNY1026-7A, Δjem1 Δscj1) cells were pulse labeled with 35S–amino acids at 23°C for 10 min in the presence of 10 μg/ml tunicamycin, and chased for the indicated times at either 23 or 37°C. pαF was recovered from cell extracts by immunoprecipitation using an anti-ppαF antiserum and analyzed by SDS-PAGE. Relative amounts of pαF were quantified by radioimaging and the means of two independent experiments are shown. The amount of pαF at 0 min chase was set to 100%. □, wild-type chased at 23°C; ○, wild-type chased at 37°C; ▪, Δjem1 Δscj1 chased at 23°C; •, Δjem1 Δscj1 chased at 37°C. (B) kar2-159 (RSY579, kar2-159) and wild-type (SNY1084-1D, wt) cells were pulse labeled with 35S–amino acids for 10 min at 23°C in the presence of 10 μg/ml tunicamycin, and were chased for the times indicated at 23 or 37°C. Degradation of pαF was followed as in A. ▪, kar2-159 chased at 23°C; •, kar2-159 chased at 37°C; □, wild-type chased at 23°C; ○, wild-type chased at 37°C.
Jem1p and Scj1p Do Not Facilitate the ERAD of a Membrane Protein
To determine whether Jem1p and Scj1p are also required for the degradation of an integral membrane ERAD substrate, we examined the fate of a mutant form of Sec61p encoded by the temperature-sensitive sec61-2 allele. Sec61-2p is selectively degraded by the ubiquitin-proteasome pathway at the restrictive temperature of 38°C, as mutations in ubiqutin-conjugating enzymes or proteasomal subunits inhibit the degradation (Biederer et al. 1996). In agreement with these earlier findings, we found that 56 and 78% of Sec61-2p were degraded after 80 min and 150 min of chase, respectively, in wild-type cells (Fig. 5). We also observed the stabilization of Sec61-2p to a similar extent either when the activity of the ubiquitin-proteasome pathway was compromised by treating cells with MG132 (Fig. 5 A), an inhibitor of the 26S proteasome (Lee and Goldberg 1996), or when the degradation of Sec61-2p was examined in a strain containing a disruption of the DOA4 gene (Fig. 5 B), which encodes a deubiquitination enzyme (Papa and Hochstrasser 1993; Hochstrasser 1996). On the other hand, we failed to observe significant stabilization of Sec61-2p in strains disrupted for JEM1, SCJ1, or both genes in combination (Fig. 5 C), compared with the strong stabilization of CPY* and ΔGpαF under these conditions (Fig. 1 and Fig. 3). It should be noted that the amount of Sec61-2p at 0 min in the chase was higher in the Δscj1 and the Δjem1Δscj1 mutant cells because the SEC61 gene is induced (data not shown). This result is consistent with Scj1p and Jem1p playing a role in ERAD, because conditions that compromise ERAD constitutively upregulate the unfolded protein response in yeast and the SEC61 gene is one target of this response (Travers et al. 2000)
Figure 5.
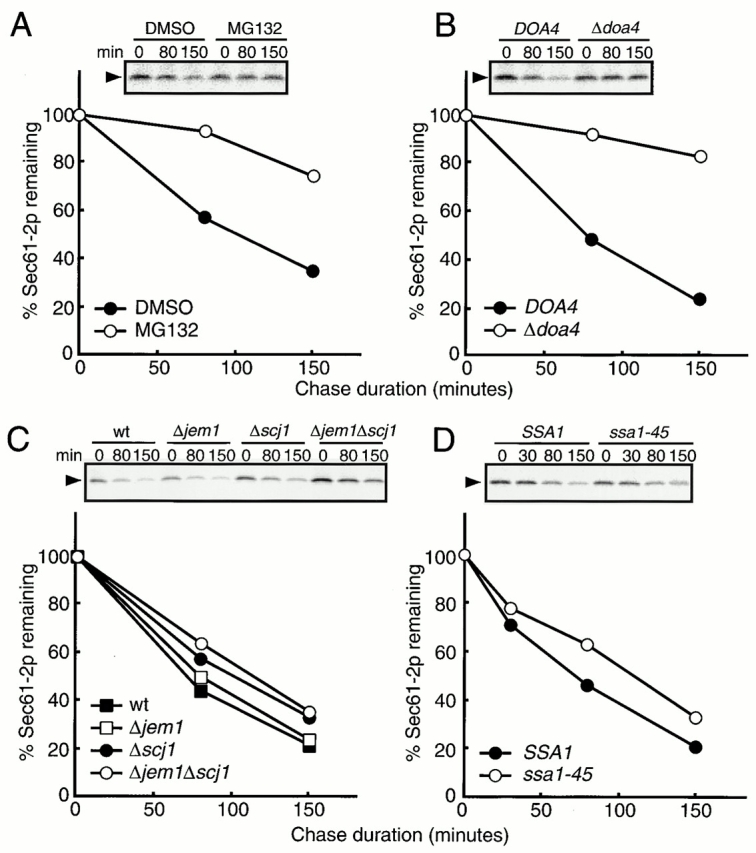
Jem1p and Scj1p are dispensable for the degradation of Sec61-2p. (A) Δerg6 cells with the sec61-2 background (SNY1081) were pulse labeled with 35S–amino acids for 10 min, and chased for the times indicated above the lanes at 38°C in the presence (MG132) or the absence (DMSO) of 200 μM MG132. Sec61-2p was recovered from cell extracts by immunoprecipitation using an anti-Sec61p antiserum and analyzed by SDS-PAGE. Relative amounts of Sec61-2p were quantified by radioimaging and the means of two independent experiments are shown. The amount of Sec61-2p at 0 min chase was set to 100%. •, pulse–chase experiment in the absence of MG132; ○, pulse–chase experiments in the presence of 200 μM MG132. (B) Δdoa4 (SNY1082) and its isogenic wild-type (DOA4, RDM15-5B) with the sec61-2 background were pulse labeled with 35S–amino acids for 10 min, chased for the times indicated above the lanes at 38°C, and analyzed as in A. The means of four independent experiments are shown. The amount of Sec61-2p at 0 min chase was set to 100%. •, wild-type; ○, Δdoa4. (C) Wild-type (RDM15-5B), Δjem1 (KYSC11), Δscj1 (SNY1070), and Δjem1 Δscj1 (SNY1071) cells with the sec61-2 background were pulse labeled with 35S–amino acids for 10 min, chased for the times indicated above the lanes at 38°C, and analyzed as in A. The means of two independent experiments are shown. The amount of Sec61-2p at 0 min chase was set to 100%. ▪, wild-type; □, Δjem1; •, Δscj1; ○, Δjem1 Δscj11. (D) ssa1-45 (SNY1078) and its isogenic wild-type (SSA1, SNY1077) strains with the sec61-2 background were pulse labeled with 35S–amino acids for 10 min, chased for the times indicated above the lanes at 37°C, and analyzed as in A. The means of two independent experiments are shown. The amount of Sec61-2p at 0 min chase was set to 100%. •, wild-type; ○, ssa1-45.
Because the degradation of three integral ER membrane proteins, Vph1p, CFTR, and Ste6p*, is suppressed in the yeast ER by compromising the activity of the Ssa cytosolic Hsp70 chaperones (Loayza et al. 1998; Hill and Cooper 2000; Zhang et al. 2001; Harper, C., J.L. Brodsky, and S. Michaelis, manuscript in preparation), we tested whether the proteolysis of Sec61-2p was reduced in the temperature-sensitive ssa1-45 mutant (Becker et al. 1996) at a nonpermissive temperature by pulse–chase analysis. Although the degradation of Vph1p, CFTR, and Ste6p* is attenuated in this strain under these conditions, the stabilization of Sec61-2p in the ssa1-45 mutant was marginal and comparable to that in the Δjem1Δscj1 mutant (Fig. 5 D). An explanation for this discrepancy is provided in the Discussion. Nevertheless, our combined results do indicate that the Jem1 and Scj1 proteins are dispensable for the degradation of Sec61-2p, a conclusion that is in accordance with previous observations that the ERAD of membrane proteins is BiP independent (Plemper et al. 1998; Brodsky and McCracken 1999).
Discussion
Misfolded and incorrectly assembled proteins in the ER lumen are distinguished from correctly folded and assembled proteins, and are degraded by ERAD. We report here that the degradation of CPY* and ΔGpαF, two soluble ERAD substrates, was delayed both in vivo and in vitro when BiP or Scj1p and Jem1p function is attenuated at elevated temperature. These results suggest that Jem1p and Scj1p function as major partners for BiP in the ERAD process. In contrast, ERAD activity appears only marginally affected when Sec63p function is ablated. The role of Sec63p in the ERAD of soluble proteins is controversial (Pilon et al. 1997; Plemper et al. 1997) and may depend on the conditions and assays employed in these unique studies.
To determine the molecular basis for our observations, we developed methods to biochemically analyze the oligomeric state of ERAD substrates. When degradation was compromised either in vivo or in vitro, we noted that the aberrant immature proteins aggregated and migrated in the densest fraction upon sucrose gradient centrifugation. Thus, one role for the BiP-Jem1p-Scj1p chaperone system is to retain ERAD substrates as lower molecular weight species. This is because the diameter of the Sec61p translocation pore has been estimated at 40–60 Å using fluorescent, actively translocating preproteins (Hamman et al. 1998), or 20 Å using three-dimensional reconstruction (Henein et al. 1996); therefore, high molecular weight oligomers or aggregates would remain trapped in the ER lumen and would be prevented from being exported to the cytoplasm for proteasome-mediated degradation. Such a role for molecular chaperones in protein transport through translocation channels is not without precedent. For example, the cytosolic Hsp70s in yeast facilitate protein translocation into both the ER lumen and mitochondria by preventing protein aggregation (Chirico et al. 1988; Deshaies et al. 1988; Murakami et al. 1988; Endo et al. 1996).
We do not rule out the possibility that the action of the BiP-Jem1p-Scj1p system in the quality control of proteins in the ER lumen may be complex, and envision three, nonmutually exclusive scenarios by which these chaperones may engineer ERAD. First, BiP, Jem1p, and Scj1p likely function together to facilitate protein folding and assembly in the ER (Silberstein et al. 1998). Thus, if proteins fail to fold correctly in the ER lumen, BiP may remain bound to them. In wild-type cells, Scj1p/Jem1p could trigger the release of these substrates from BiP, which are then retrotranslocated from the ER and degraded. However, in the absence of Scj1p/Jem1p action, the substrate remains in the ER and eventually aggregates. Second, misfolded proteins may be recognized by components of a secondary ER export machinery. The BiP-Jem1p-Scj1p complex might transfer ERAD substrates to this system, through which they are ultimately delivered to the translocation channel for export. Candidates for this system are the calnexin-calreticulin chaperones that are known to play a role in ERAD (McCracken and Brodsky 1996; Qu et al. 1996; Liu et al. 1997; for review see Ellgaard et al. 1999). Furthermore, yeast calnexin has been shown to associate both biochemically (Jannatipour et al. 1998) and genetically (Brodsky et al. 1999) with BiP. The future development of an in vitro, ordered, export reaction will address this hypothesis. Third, BiP has been shown to regulate the translocation pore in the mammalian ER (Hamman et al. 1998). It is possible that yeast BiP, in conjunction with Scj1p and Jem1p, performs an analogous role during retrotranslocation. In the event that the channel cannot open, ERAD substrates may aggregate, as observed here.
The Lhs1p chaperone has been shown to aid in the refolding of heat denatured proteins in the ER (Saris et al. 1997). Not surprisingly, an unfolded protein response enhancer resides upstream of the LHS1 gene, and Lhs1p is induced by conditions that lead to the accumulation of unfolded proteins in the ER (Craven et al. 1996; Cassagrande et al. 2000; Travers et al. 2000). However, we show here that Lhs1p is not required for ERAD, indicating that its synthesis is induced to help re-fold denatured proteins, but not to destroy them. Thus, the synthetic lethal interactions observed in cells deleted for LHS1 and containing a thermosensitive KAR2 allele (Baxter et al. 1996; Craven et al. 1996; Hamilton and Flynn 1996) suggest that BiP and Lhs1p cooperate in this refolding process.
Our failure to observe SSA-dependent degradation of Sec61-2p (Fig. 5 B) was initially surprising, as three integral membrane ERAD substrates in yeast are stabilized in the ssa1-45 strain (Hill and Cooper 2000; Zhang et al. 2001; Harper, C., J.L. Brodsky, and S. Michaelis, manuscript in preparation). However, it should be noted that each of these proteins contains large cytoplasmic domains that may have to be retained in a soluble, degradation-competent conformation. In contrast, Sec61-2p lacks such domains, is overall quite hydrophobic, and therefore its proteolysis might occur independently of cytoplasmic chaperone action. One intriguing hypothesis forwarded by Mayer et al. 1998 is that the proteasome is necessary and sufficient to extract membrane proteins for ERAD. The construction of synthetic, integral membrane ERAD substrates, as was used by these investigators, will allow us to better examine this hypothesis.
Upon sequencing the integrated copy of the sec61-2 gene used for the experiments described in the legend to Fig. 5, we discovered that the mutation in this thermosensitive allele of SEC61 alters a glycine to an aspartic acid at position 213 (see Materials and Methods). From the work of Stirling and colleagues 1992, this residue is presumed to lie at the start of transmembrane domain 5 (Wilkinson et al. 1996). This domain contains only 12 amino acids, 11 of which are nonpolar, and has been proposed to require the stabilizing effect of downstream sequences to ensure its proper insertion into the ER membrane. It was also reported that Sec61-2p was stabilized by overexpression of Sss1p, another subunit of the Sec61 complex, and in turn, Sss1p was destabilized in strains containing the sec61-2 allele (Esnault et al. 1994). The mutation could affect insertion into the ER membrane and/or interactions with Sss1p of Sec61-2p at elevated temperatures. We labeled the sec61-2 strain at permissive temperature, then switched to restrictive temperature and observed rapid turnover of Sec61-2p (Fig. 4). Therefore, the latter scenario, suggesting the failure of Sec61-2p to interact with Sss1p at elevated temperatures, appears more likely.
In sum, this study reveals one role for the lumenal Hsp70 chaperone system in the export of aberrant proteins from the ER lumen to the cytosol, but how ERAD substrates are initially identified and targeted for the export to the cytosol remains unanswered. As exemplified in this work, the continued development of novel biochemical assays, using genetic tools, will be imperative to continue to define this pathway.
Acknowledgments
We thank Mark Rose, Randy Schekman, Scott Emr, Yoshinori Ohsumi, Kyohei Umebayashi, Tom Stevens, and Akihiko Nakano for materials, and Tohru Yoshihisa and Colin Stirling for valuable discussions.
This work was supported by Grants-in-aid for Scientific Research from the Ministry of Education (T. Endo and S. Nishikawa), and grant MCB-9722889 from the National Science Foundation (to J.L. Brodsky). S.W. Fewell acknowledges a National Research Service Award from the National Institutes of Health.
Footnotes
Abbreviations used in this paper: CPY, carboxypeptidase Y; ERAD, ER-associated degradation; pαF, pro–α-factor; ppαF, prepro–α-factor.
References
- Baxter B.K., James P., Evans T., Craig E.A. SSI1 encodes a novel Hsp70 of the Saccharomyces cerevisiae endoplasmic reticulum. Mol. Cell. Biol. 1996;16:6444–6456. doi: 10.1128/mcb.16.11.6444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Becker J., Walter W., Yan W., Craig E.A. Functional interaction of cytosolic hsp70 and a DnaJ-related protein, Ydj1p, in protein translocation in vivo. Mol. Cell. Biol. 1996;16:4378–4386. doi: 10.1128/mcb.16.8.4378. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Biederer T., Volkwein C., Sommer T. Degradation of subunits of the Sec61p complex, an integral component of the ER membrane, by the ubiquitin-proteasome pathway. EMBO J. 1996;15:2069–2076. [PMC free article] [PubMed] [Google Scholar]
- Brizzio V., Khalfan W., Huddler D., Beh C.T., Andersen S.S.L., Latterich M., Rose M.D. Genetic interactions between KAR7/SEC71, KAR8/JEM1, KAR5, and KAR2 during nuclear fusion in Saccharomyces cerevisiae . Mol. Biol. Cell. 1999;10:609–626. doi: 10.1091/mbc.10.3.609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodsky J.L., McCracken A.A. ER protein quality control and proteasome-mediated protein degradation. Semin. Cell Dev. Biol. 1999;10:507–513. doi: 10.1006/scdb.1999.0321. [DOI] [PubMed] [Google Scholar]
- Brodsky J.L., Schekman R. A Sec63p–BiP complex from yeast is required for protein translocation in a reconstituted proteoliposome. J. Cell Biol. 1993;123:1355–1363. doi: 10.1083/jcb.123.6.1355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brodsky J.L., Werner E.D., Dubas M.E., Goeckeler J.L., Kruse K.B., McCracken A.A. The requirement for molecular chaperones during endoplasmic reticulum-associated protein degradation demonstrates that protein export and import are mechanistically distinct. J. Biol. Chem. 1999;274:3453–3460. doi: 10.1074/jbc.274.6.3453. [DOI] [PubMed] [Google Scholar]
- Cassagrande R., Stern P., Diehn M., Shamu C., Osario M., Zuniga M., Brown P.O., Ploegh H. Degradation of proteins from the ER of S. cerevisiae requires an intact unfolded protein response pathway. Mol. Cell. 2000;5:729–735. doi: 10.1016/s1097-2765(00)80251-8. [DOI] [PubMed] [Google Scholar]
- Chirico W.J., Waters M.G., Blobel G. 70K heat shock-related proteins stimulate protein translocation into microsomes. Nature. 1988;332:805–810. doi: 10.1038/332805a0. [DOI] [PubMed] [Google Scholar]
- Corsi A.K., Schekman R. The lumenal domain of Sec63p stimulates the ATPase activity of BiP and mediates BiP recruitment to the translocon in Saccharomyces cerevisiae . J. Cell Biol. 1997;137:1483–1493. doi: 10.1083/jcb.137.7.1483. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Craven R.A., Egerton M., Stirling C.J. A novel Hsp70 of the yeast ER lumen is required for the efficient translocation of a number of protein precursors. EMBO J. 1996;15:2640–2650. [PMC free article] [PubMed] [Google Scholar]
- Deshaies R.J., Schekman R. A yeast mutant defective at an early stage in import of secretory protein precursors into the endoplasmic reticulum. J. Cell Biol. 1987;105:633–645. doi: 10.1083/jcb.105.2.633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deshaies R.J., Koch B.D., Werner-Washburne M., Craig E.A., Schekman R. A subfamily of stress proteins facilitates translocation of secretory and mitochondrial precursor polypeptides. Nature. 1988;332:800–805. doi: 10.1038/332800a0. [DOI] [PubMed] [Google Scholar]
- Ellgaard L., Molinari M., Helenius A. Setting the standardsquality control in the secretory pathway. Science. 1999;286:1882–1888. doi: 10.1126/science.286.5446.1882. [DOI] [PubMed] [Google Scholar]
- Endo T., Mitsui S., Nakai M., Roise D. Binding of mitochondrial presequences to yeast cytosolic heat shock protein 70 depends on the amphiphilicity of the presequence. J. Biol. Chem. 1996;271:4161–4167. doi: 10.1074/jbc.271.8.4161. [DOI] [PubMed] [Google Scholar]
- Esnault Y., Feldheim D., Blondel M.-O., Schekman R., Kepes F. SSS1 encodes a stabilizing component of the Sec61 subcomplex of the yeast protein translocation apparatus. J. Biol. Chem. 1994;269:27478–27485. [PubMed] [Google Scholar]
- Finger A., Knop M., Wolf D.H. Analysis of two mutated vacuolar proteins reveals a degradative pathway in the endoplasmic reticulum or a related compartment in yeast. Eur. J. Biochem. 1993;218:565–574. doi: 10.1111/j.1432-1033.1993.tb18410.x. [DOI] [PubMed] [Google Scholar]
- Hamilton T.G., Flynn G.C. Cer1p, a novel Hsp70-related protein required for posttranslational endoplasmic reticulum translocation in yeast. J. Biol. Chem. 1996;271:30610–30613. doi: 10.1074/jbc.271.48.30610. [DOI] [PubMed] [Google Scholar]
- Hamman B.D., Hendershot L.M., Johnson A.E. BiP maintains the permeability barrier of the ER membrane by sealing the lumenal end of the translocon pore before and early in translocation. Cell. 1998;92:747–758. doi: 10.1016/s0092-8674(00)81403-8. [DOI] [PubMed] [Google Scholar]
- Henein D., Matlack K.E.S., Jungnickel B., Plath K., Kalies K.-U., Miller K.R., Rapoport T.A., Akey C.W. Oligomeric rings of the Sec61p complex induced by ligands required for protein translocation. Cell. 1996;87:721–732. doi: 10.1016/s0092-8674(00)81391-4. [DOI] [PubMed] [Google Scholar]
- Hill K., Cooper A.A. Degradation of unassembled Vph1p reveals novel aspects of the yeast ER quality control system. EMBO J. 2000;19:550–561. doi: 10.1093/emboj/19.4.550. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hiller M.M., Finer A., Schweiger M., Wolf D.H. ER degradation of a misfolded lumenal protein by the cytosolic ubiquitin-proteasome pathway. Science. 1996;273:1725–1728. doi: 10.1126/science.273.5282.1725. [DOI] [PubMed] [Google Scholar]
- Hochstrasser M. Ubiquitin-dependent protein degradation. Annu. Rev. Genet. 1996;30:405–439. doi: 10.1146/annurev.genet.30.1.405. [DOI] [PubMed] [Google Scholar]
- Jannatipour M., Callejo M., Parodi A.J., Armstrong J., Rokeach L.A. Calnexin and BiP interact with acid phosphatase independently of glucose trimming and reglucosylation in Schizosaccharomyces pombe . Biochemistry. 1998;37:17253–17261. doi: 10.1021/bi981785c. [DOI] [PubMed] [Google Scholar]
- Jones E.W. Tackling the protease problem in Saccharomyces cerevisiae . Methods Enzymol. 1991;194:428–453. doi: 10.1016/0076-6879(91)94034-a. [DOI] [PubMed] [Google Scholar]
- Jones J.S., Prakash L. Yeast Saccharomyces cerevisiae selectable markers in pUC18 polylinkers. Yeast. 1990;6:363–366. doi: 10.1002/yea.320060502. [DOI] [PubMed] [Google Scholar]
- Kelley W.L. The J-domain family and the recruitment of chaperone power. Trends Biochem. Sci. 1998;23:222–227. doi: 10.1016/s0968-0004(98)01215-8. [DOI] [PubMed] [Google Scholar]
- Kunkel T.A., Roberts J.D., Zakour R.A. Rapid and efficient site-specific mutagenesis without phenotypic selection. Methods Enzymol. 1987;154:367–382. doi: 10.1016/0076-6879(87)54085-x. [DOI] [PubMed] [Google Scholar]
- Lee D.H., Goldberg A.L. Selective inhibitors of the proteasome-dependent and vacuolar pathways of protein degradation in Saccharomyces cerevisiae . J. Biol. Chem. 1996;271:22280–22284. doi: 10.1074/jbc.271.44.27280. [DOI] [PubMed] [Google Scholar]
- Liberek K., Marszalek J., Ang D., Georgopoulos C., Zylicz M. Escherichia coli DnaK and GrpE heat shock proteins jointly stimulate ATPase activity of SnaK. Proc. Natl. Acad. Sci. USA. 1991;88:2874–2878. doi: 10.1073/pnas.88.7.2874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu Y., Choudhury P., Cabral C.M., Sifers R.N. Intracellular disposal of incompletely folded human α1-antitrypsin involves release from calnexin and post-translational trimming of asparagine-linked oligosaccharides. J. Biol. Chem. 1997;272:7946–7951. doi: 10.1074/jbc.272.12.7946. [DOI] [PubMed] [Google Scholar]
- Loayza D., Tam A., Schmidt W.K., Michaelis S. Ste6p mutants defective in exit from the endoplasmic reticulum (ER) reveal aspects of the ER quality control pathway in Saccharomyces cerevisiae . Mol. Biol. Cell. 1998;9:2767–2784. doi: 10.1091/mbc.9.10.2767. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lyman S.K., Schekman R. Interaction between BiP and Sec63p is required for the completion of protein translocation into the ER of Saccharomyces cerevisiae . J. Cell Biol. 1995;131:1163–1171. doi: 10.1083/jcb.131.5.1163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Matlack K.E., Misselwitz B., Plath K., Rapoport T.A. BiP acts as a molecular ratchet during posttranslational transport of prepro-alpha factor across the ER membrane. Cell. 1999;97:553–564. doi: 10.1016/s0092-8674(00)80767-9. [DOI] [PubMed] [Google Scholar]
- Mayer T.U., Braun T., Jentsch S. Role of the proteasome in membrane extraction of a short-lived ER transmembrane protein. EMBO J. 1998;17:3251–3257. doi: 10.1093/emboj/17.12.3251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McCracken A.A., Brodsky J.L. Assembly of ER-associated degradation in vitrodependence on cytosol, calnexin and ATP. J. Cell Biol. 1996;132:291–298. doi: 10.1083/jcb.132.3.291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murakami H., Pain D., Blobel G. 70-kD heat shock-related protein is one of at least two distinct cytosolic factors stimulating protein import into mitochondria. J. Cell Biol. 1988;107:2051–2057. doi: 10.1083/jcb.107.6.2051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nishikawa S., Endo T. The yeast JEM1p is a DnaJ-like protein of the endoplasmic reticulum membrane required for nuclear fusion. J. Biol. Chem. 1997;272:12889–12892. doi: 10.1074/jbc.272.20.12889. [DOI] [PubMed] [Google Scholar]
- Nishikawa S., Nakano A. The GTP-binding Sar1 protein is localized to the early compartment of the yeast secretory pathway. Biochim. Biophys. Acta. 1991;1093:135–143. doi: 10.1016/0167-4889(91)90114-d. [DOI] [PubMed] [Google Scholar]
- Nishikawa S., Hirata A., Nakano A. Inhibition of endoplasmic reticulum (ER)-to-Golgi transport induces relocalization of binding protein (BiP) within the ER to form the BiP bodies. Mol. Biol. Cell. 1994;5:1129–1143. doi: 10.1091/mbc.5.10.1129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Papa F.R., Hochstrasser M. The yeast DOA4 gene encodes a deubiquitinating enzyme related to a product of the human tre-2 oncogene. Nature. 1993;366:313–319. doi: 10.1038/366313a0. [DOI] [PubMed] [Google Scholar]
- Pilon M., Schekman R., Römisch K. Sec61p mediates export of a misfolded secretory protein from the endoplasmic reticulum to the cytosol for degradation. EMBO J. 1997;16:4540–4548. doi: 10.1093/emboj/16.15.4540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pilon M., Schekman R., Romisch K. Sec61p serves multiple roles in secretory precursor binding and translocation into endoplasmic reticulum membrane. Mol. Biol. Cell. 1998;9:3455–3473. doi: 10.1091/mbc.9.12.3455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plemper R.K., Wolf D.H. Retrograde protein translocationERADication of secretory proteins in health and disease. Trends Biochem. Sci. 1999;24:266–270. doi: 10.1016/s0968-0004(99)01420-6. [DOI] [PubMed] [Google Scholar]
- Plemper R.K., Bohmler S., Bordallo J., Sommer T., Wolf D.H. Mutant analysis links the translocon and BiP to retrograde protein transport for ER degradation. Nature. 1997;388:891–895. doi: 10.1038/42276. [DOI] [PubMed] [Google Scholar]
- Plemper R.K., Egner R., Kuchler K., Wolf D.H. Endoplasmic reticulum degradation of a mutated ATP-binding cassette transporter Pdr5 proceeds in a concerted action of Sec61 and the proteasome. J. Biol. Chem. 1998;273:32848–32856. doi: 10.1074/jbc.273.49.32848. [DOI] [PubMed] [Google Scholar]
- Plemper R.K., Deak P.M., Otto R.T., Wolf D.H. Re-entering the translocon from the lumenal side of the endoplasmic reticulum. Studies on the mutated carboxypeptidase yscY species. FEBS Lett. 1999;443:241–245. doi: 10.1016/s0014-5793(98)01724-4. [DOI] [PubMed] [Google Scholar]
- Qu D., Teckman J.H., Omura S., Perlmutter D.H. Degradation of a mutant secretory protein, α1-antitrypsin Z, in the endoplasmic reticulum requires proteasome activity. J. Biol. Chem. 1996;271:22791–22795. doi: 10.1074/jbc.271.37.22791. [DOI] [PubMed] [Google Scholar]
- Robinson J.S., Klionsky D.J., Banta L.M., Emr S.D. Protein sorting in Saccharomyces cerevisiaeisolation of mutants defective in the delivery and processing of multiple vacuolar hydrolases. Mol. Cell. Biol. 1988;8:4936–4948. doi: 10.1128/mcb.8.11.4936. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Römisch K. Surfing the Sec61 channelbidirectional protein translocation across the ER membrane. J. Cell Sci. 1999;112:4185–4191. doi: 10.1242/jcs.112.23.4185. [DOI] [PubMed] [Google Scholar]
- Rose M.D., Winston F., Hieter P. Methods in Yeast GeneticsA Laboratory Course Manual 1990. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: pp. 198 pp [Google Scholar]
- Rothblatt J., Schekman R. A hitchhiker's guide to analysis of the secretory pathway in yeast. Methods Cell Biol. 1989;32:3–36. doi: 10.1016/s0091-679x(08)61165-6. [DOI] [PubMed] [Google Scholar]
- Rothblatt J.A., Deshaies R.J., Sanders S.L., Daum G., Schekman R. Multiple genes are required for proper insertion of secretory proteins into the endoplasmic reticulum in yeast. J. Cell Biol. 1989;109:2641–2652. doi: 10.1083/jcb.109.6.2641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sadler I., Chiang A., Kurihara T., Rothblatt J., Way J., Silver P. A yeast gene important for protein assembly into the endoplasmic reticulum and the nucleus has homology to DnaJ, an Escherichia coli heat shock protein. J. Cell Biol. 1989;109:2665–2675. doi: 10.1083/jcb.109.6.2665. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sanders S.L., Whitfield K.M., Vogel J.P., Rose M.D., Schekman R.W. Sec61p and BiP directly facilitate polypeptide translocation into the ER. Cell. 1992;69:353–365. doi: 10.1016/0092-8674(92)90415-9. [DOI] [PubMed] [Google Scholar]
- Saris N., Holkeri H., Craven R.A., Stirling C.J., Makarow M. The hsp70 homologue Lhs1p is involved in a novel function of the yeast endoplasmic reticulum, refolding and stabilization of heat-denatured protein aggregates. J. Cell Biol. 1997;137:813–824. doi: 10.1083/jcb.137.4.813. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlenstedt G., Harris S., Risse B., Lill R., Silver P.A. A yeast DnaJ homologue, Scj1p, can function in the endoplasmic reticulum with BiP/Kar2p via a conserved domain that specifies interactions with Hsp70s. J. Cell Biol. 1995;129:979–988. doi: 10.1083/jcb.129.4.979. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scidmore M.A., Okamura H.H., Rose M.D. Genetic interactions between KAR2 and SEC63 encoding homologues of DnaK and DnaJ in the endoplasmic reticulum. Mol. Biol. Cell. 1993;4:1145–1159. doi: 10.1091/mbc.4.11.1145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Silberstein S., Schlenstedt G., Silver P.A., Gilmore R. A role for the DnaJ homologue Scj1p in protein folding in the yeast endoplasmic reticulum. J. Cell Biol. 1998;143:921–933. doi: 10.1083/jcb.143.4.921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sikorski R.S., Hieter P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevisiae . Genetics. 1989;122:19–27. doi: 10.1093/genetics/122.1.19. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simons J.F., Ferro-Novick S., Rose M.D., Helenius A. BiP/Kar2p serves as a molecular chaperone during carboxypeptidase Y in yeast. J. Cell Biol. 1995;130:41–49. doi: 10.1083/jcb.130.1.41. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens T.H., Rothman J.H., Payne G.S., Schekman R. Gene dosage-dependent secretion of yeast vacuolar carboxypeptidase Y. J. Cell Biol. 1986;102:1551–1557. doi: 10.1083/jcb.102.5.1551. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stirling T.H., Rothblatt J., Hosobuchi M., Deshaies R., Schekman R. Protein translocation mutants defective in the insertion of integral membrane proteins into the endoplasmic reticulum. Mol. Biol. Cell. 1992;3:129–142. doi: 10.1091/mbc.3.2.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Travers K.J., Patil C.K., Wodicka L., Lockhart D.J., Weissman J.S., Walter P. Functional and genomic analyses reveal an essential coordination between the unfolded protein response and ER-associated degradation. Cell. 2000;101:249–258. doi: 10.1016/s0092-8674(00)80835-1. [DOI] [PubMed] [Google Scholar]
- Werner E.D., Brodsky J.L., McCracken A.A. Proteasome-associated endoplasmic reticulum associated protein degradationan unconventional route to a familiar fate. Proc. Natl. Acad. Sci. USA. 1996;93:13797–13801. doi: 10.1073/pnas.93.24.13797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wiertz E.J., Tortorella D., Bogyo M., Yu J., Mothes W., Jones T.R., Rapoport T.A., Ploegh H.L. Sec61-mediated transfer of a membrane protein from the endoplasmic reticulum to the proteasome for destruction. Nature. 1996;384:432–438. doi: 10.1038/384432a0. [DOI] [PubMed] [Google Scholar]
- Wilkinson B.M., Critchley A.J., Stirling C.J. Determination of the transmembrane topology of yeast Sec61p, an essential component of the endoplasmic reticulum translocation complex. J. Biol. Chem. 1996;271:25590–25597. doi: 10.1074/jbc.271.41.25590. [DOI] [PubMed] [Google Scholar]
- Wilkinson B.M., Tyson J.R., Reid P.J., Stirling C.J. Distinct domains within yeast Sec61p involved in post-translational translocation and protein dislocation. J. Biol. Chem. 2000;275:521–529. doi: 10.1074/jbc.275.1.521. [DOI] [PubMed] [Google Scholar]
- Zhang Y., Nijbroek G., Sullivan M.L., McCracken A.A., Watkins S.C., Michaelis S., Brodsky J.L. The Hsp70 molecular chaperone facilitates the ER associated protein degradation of the cystic fibrosis transmembrane conductance regulator in yeast. Mol. Biol. Cell. 2001;12:1303–1314. doi: 10.1091/mbc.12.5.1303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou M., Schekman R. The engagement of Sec61p in the ER dislocation process. Mol. Cell. 1999;4:925–934. doi: 10.1016/s1097-2765(00)80222-1. [DOI] [PubMed] [Google Scholar]