
Figure 2.
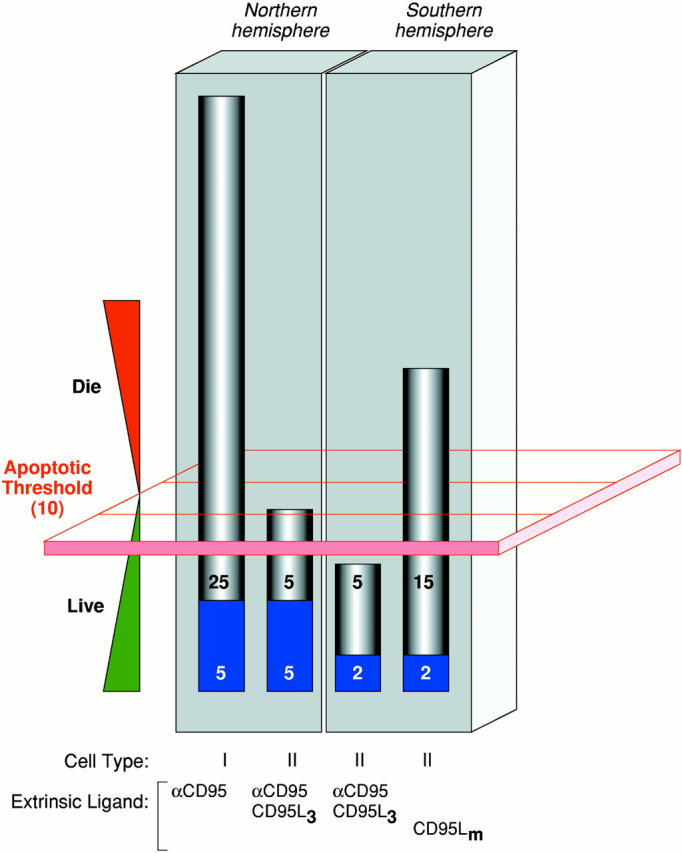
How apoptotic thresholding might influence experimental outcome. Many possible explanations may account for the differential sensitivity of cells from the ‘Northern hemisphere’ labs (e.g., Krammer/Peter) versus those from the ‘Southern hemisphere’ (e.g., Strasser) to CD95 stimulation (the Krammer/Peter labs are located in Germany and the Strasser lab is in Australia, hence the Northern and Southern hemisphere references.). Herein is illustrated one hypothesis. It assumes that most cell types have an apoptotic threshold that, when breached, results in apoptotic cell death. This apoptotic threshold (represented by the red plane) requires, say, 10 arbitrary apoptotic units (which could be, among other things, a critical mass of caspase catalytic activity). The ability of a cell to cross this threshold depends on the combined contributions of the extrinsic cell death pathway (gray) and the fraction of the intrinsic pathway (blue) that can be engaged through pathway cross-talk. Type I cells (first bar), in both the Northern and Southern hemispheres, harbor a very efficient DISC mechanism that alone can contribute the necessary apoptotic units to breach the threshold. Thus, regardless of the status of the intrinsic pathway, and even if it is completely blocked by the presence of Bcl-2, the type I cell can muster the necessary currency to engage the apoptotic machinery and die. Therefore, extrinsic and intrinsic pathways in type I cells appear independent (assuming intrinsic does not talk to extrinsic). In type II cells, DISC formation is weaker and the extrinsic pathway depends more on contributions from the intrinsic pathway. These cells behave differently in the Northern hemisphere versus the Southern hemisphere. In the Northern hemisphere, the combination of the extrinsic contribution and the fraction of the intrinsic pathway that it is able to engage is sufficient for apoptosis to proceed (second bar). Attenuation of the intrinsic component, by Bcl-2 overexpression for example, reduces the total number of apoptotic units to below the apoptotic threshold and the cells survive. The extrinsic pathway, therefore, appears to be modulated by Bcl-2. In the Southern hemisphere, the number of available apoptotic units that can be contributed by the intrinsic pathway may be lower. Thus, engagement of the extrinsic pathway with ligating CD95 (Fas/Apo1) antibody or trimerized ligand is not sufficient to mediate passage across the apoptotic threshold on its own (third bar). With a more efficient stimulus, mediated by multimerized CD95L, for example (fourth bar), a critical mass of the extrinsic pathway is enlisted to breach the apoptotic threshold, regardless of the contribution from the intrinsic pathway. Cell death in this scenario is therefore insensitive to Bcl-2 overexpression. The differences observed in the Northern and Southern hemispheres might therefore be explained by a combination of (a) the number of apoptotic units within the intrinsic pathway that can be recruited by cross-talk from the extrinsic pathway (lower in the South) and (b) the strength of the apoptotic stimulus (ligating antibodies and trimerized CD95L being equivalent but weaker than multimerized CD95L).