

Abstract
Despite considerable interest in using stable-hydrogen isotope ratio (δD) measurements in ecological research, it was previously unknown whether hydrogen derived from drinking water, in addition to that derived from diet, contributed to the nonexchangeable hydrogen in animal tissues. We raised four experimental groups of quail (Coturnix coturnix japonica) from hatch on two isotopically distinct diets (mean nonexchangeable δD: −146 and −60‰, Vienna Standard Mean Ocean Water Standard) and drinking waters (mean δD: −130 and +196‰, Vienna Standard Mean Ocean Water Standard). Here we show that both dietary and drinking water hydrogen are incorporated into nonexchangeable hydrogen in both metabolically active (i.e., muscle, liver, blood, fat) and inactive (i.e., feather, nail) tissues. Approximately 20% of hydrogen in metabolically active quail tissues and 26–32% of feathers and nail was derived from drinking water. Our findings suggest environmental interpretations of δD values from modern and fossil animal tissues may need to account for potentially large isotopic differences between drinking water and food and require a good understanding of the physiological ecology of study organisms.
Previous studies have shown that the natural stable-isotope ratios of hydrogen (δD) in various animal tissues are a powerful tool in food web delineation (1–3), paleoclimatic reconstruction (4–6), and animal migration tracking (7, 8). In particular, body tissues that are biochemically inert following synthesis in living organisms (e.g., hair, feathers) or archaeological and fossil tissues (e.g., bone collagen, chitin) that have not been subject to diagenetic alteration can provide key environmental and paleoclimatic information. Despite the potential of this area of research, little attention has been paid to the relative roles of diet and drinking water and their effect on the overall hydrogen isotopic composition of animal tissues. In addition, the δD values of faunal tissues may be controlled by a variety of factors, including metabolic and biosynthetic isotope fractionation, the δD values of food, and hydrogen isotope exchange with ambient water and body water (9). For δD measurements to be used effectively in food web or dietary reconstruction, it is necessary to understand how hydrogen isotopes fractionate between dietary hydrogen and consumer tissue and the relative contribution of drinking water to the δD value of nonexchangeable hydrogen in that tissue. Despite previous studies investigating hydrogen isotope fractionation between diet and tissues of animals (1, 2, 9, 10), the relative contributions of food and drinking-water to various tissue δD values has not been established previously, and the isotopic fractionation of deuterium between diet and consumer tissues remains unclear (2, 3, 11).
DeNiro and Epstein (9) first showed that exposure to atmospheric water vapor of different δD values changed the overall δD value of mouse food and the brain and liver tissue of mice. Hydrogen atoms are held weakly by oxygen and nitrogen, so ambient water vapor readily exchanges hydrogen with available oxygen- or nitrogen-bound hydrogen in samples of organic material (12). The extent of hydrogen isotopic exchange is temperature-dependent, but it has been determined empirically that exchangeable hydrogen can account for up to ≈20% of the total hydrogen of dehydrated organic materials (7, 8). Only the nonexchangeable hydrogen portion of a tissue reveals information on dietary sources of hydrogen, but this could in turn be complicated by the incorporation drinking water hydrogen into the nonexchangeable hydrogen pool within animals. Using captive-reared Japanese quail (Coturnix coturnix japonica), we determined relative contributions of drinking water versus food to the nonexchangeable hydrogen in animal tissues under controlled conditions and ascertained whether single diet-tissue δD fractionation factors for various tissue types could be generally applied in ecological studies involving animals.
METHODS
Newly hatched Japanese quail were randomly assigned to each of four treatment groups. These groups had access to one of two batches of isotopically distinct diet and one of two sources of isotopically distinct drinking water (Table 1). We used a single homogenized batch of Texas-grown commercial turkey starter composed of 28% protein, 4% carbohydrate, and 3% fat for diet 1, and a single homogenized batch of Saskatchewan-grown commercial turkey starter composed of 16% protein, 6% carbohydrate, and 3% fat for diet 2. Both diets were formulated to meet nutritional requirements of quail. The drinking water sources were two 10-liter batches of tap water (water 1), one of which had been spiked with 300 μl of 99.9% D2O (water 2). δD values of water were measured (n = 24) throughout the experiment.
Table 1.
Tissue δD in quail
| Tissue | Diet 1
|
Diet 2
|
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Water 1 | n | Water 2 | n | Water-derived hydrogen, % | Water 1 | n | Water 2 | n | Water-derived hydrogen, % | |
| Blood | −142 ± 6 | 7* | −69 ± 3 | 7* | 22 | −162 ± 3 | 3* | −94 ± 5 | 3* | 21 |
| Muscle | −137 ± 2 | 7* | −68 ± 3 | 7* | 21 | −158 ± 1 | 3*† | −99 ± 3 | 2* | 18 |
| Liver | −127 ± 2 | 7† | −50 ± 2 | 7† | 24 | −151 ± 4 | 4† | −84 ± 1 | 3† | 21 |
| Lipids | −198 ± 3 | 7‡ | −141 ± 3 | 7‡ | 18 | −256 ± 5 | 3‡ | −194 ± 5 | 3‡ | 19 |
| Feather | −134 ± 2 | 5* | −29 ± 2 | 3§ | 32 | −152 ± 1 | 3* | −72 ± 1 | 5§ | 26 |
| Nail | −118 ± 3 | 7¶ | −31 ± 3 | 7§ | 27 | — | — | — | — | — |
Mean δD ± SD (n) of tissues from quail raised with exclusive access to one of two dietary (diet 1 = −56 ± 2%, n = 7; diet 2 = −146 ± 8%, n = 8) and drinking water (water 1 = −130 ± 1%, n = 6; water 1 = +196 ± 2%, n = 6) treatments. Calculated percent contribution of drinking water to tissue δD values for quail raised on each diet was calculated by using ΔδDtissue/ΔδDwater × 100%. For each tissue, all between-group δD values were significantly different (ANOVA, P < 0.05). Within groups, δD values with common symbols indicate no significant difference (Tukey’s test). All δD values refer to measurements of nonexchangeable hydrogen.
Birds were raised in the laboratory under identical conditions from hatch with ad libitum access to food and water until at least 23 days of age, when flight feathers were fully grown. At that time, a single outer primary feather was removed for stable-isotope analysis of nonexchangeable hydrogen. The quail were euthanized (13) at 63 days of age, and samples of blood, liver, and pectoral muscle from each bird were stored frozen and later freeze-dried and powdered with a mortar and pestle. Lipids were removed from liver and muscle by using a solution of 2:1 chloroform/methanol (14). Abdominal fat was sonicated for 1 hour in this solvent. The dissolved lipids were then filtered through a GF/C glass fiber filter paper, the solvent evaporated at room temperature for approximately 48 hours, and the filtered lipid sample frozen before analysis. Feathers were cleaned of surface oils with the chloroform/methanol solvent. We measured δD values of blood, pectoral muscle, liver, feather, and abdominal fat for all four treatments to assess the influence of both food and water on consumer tissues. Because the δD values of experimental drinking waters changed slightly through time, waters for the feather experiment (−126 and +191‰) differed slightly from those used for the longer experiment (−130 and +196‰) involving other tissues.
Although cellulose and some other materials can be nitrated to eliminate the exchangeable hydrogen fraction (15), in complex organic material nitration is not possible; thus, the exchangeable hydrogen is impossible to separate from the overall bulk hydrogen isotopic composition (12). Uncontrolled exchange of labile hydrogen in organic material with ambient water vapor necessitates that researchers account for this through equilibration procedures (6, 8, 9, 16). Samples can be standardized by exposure to water vapor of known δD value and calibrated according to the proportion of exchangeable hydrogen for each tissue type. In this way, only the differences in nonexchangeable hydrogen δD values are considered (12). The difference between the amount of theoretically exchangeable hydrogen (i.e., nitrogen- and oxygen-bound hydrogen) and the amount of hydrogen that actually exchanges in a tissue can be considerable. The fraction of nitrogen- and oxygen-bound hydrogen that undergoes exchange depends on temperature and on the tertiary and quaternary molecular structure that can render some hydrogen sterically inaccessible for exchange (10). Actual exchangeability must be determined experimentally for complex organic materials but does not permit accurate calculation of the δD of nonexchangeable hydrogen. For the purposes of this study, we refer to the fraction of hydrogen that does not exchange under our standard experimental conditions as nonexchangeable hydrogen.
Hydrogen-isotope exchangeability for all tissue types was first quantified by using a static equilibration technique with steam having a wide range of hydrogen-isotopic values (−135 to +525‰) at constant temperature (130 ± 0.1°C) for 2 hours and then measuring the total hydrogen δD values (refs. 16 and 17; Appendix). After equilibration in Vycor breakseal tubes, all water vapor was removed cryogenically. Samples were then sealed under vacuum and combusted at 850°C in the presence of cupric oxide, followed by cryogenic separation of CO2 from H2O. Waters of combustion were reduced to H2 gas by using hot zinc, and 2H/H ratios were measured on a Micromass Optima dual inlet isotope-ratio mass spectrometer. Stable-hydrogen isotope results are reported in parts per thousand deviation from the Vienna Standard Mean Ocean Water Standard (VSMOW), with a sample reproducibility better than ±2.0‰. All results in Table 1 have been corrected to account for differences in exchangeability among tissue types.
RESULTS AND DISCUSSION
Our results indicate unequivocally that both food and drinking water ultimately influenced the nonexchangeable δD values of all tissues measured (Table 1, Fig. 1). For diet 1, a δD enrichment of 326‰ in drinking water resulted in a positive shift of 57–105‰ in the mean δD value of quail tissues. Similarly, for diet 2, the same isotopic difference in drinking water caused a corresponding positive shift of 59–85‰. For food substrate, a positive difference of 86‰ caused a corresponding positive shift in tissue δD of 21–58‰ for birds raised on water 1, and 25–53‰ change in tissue δD for those raised on water 2. Identical tissues from the two drinking-water groups for each diet had significantly different δD values (t > 12, P < 0.0001 in all cases; Table 1, Fig. 1). For quail, both diets showed differences in δD values because of different drinking waters, obtaining approximately 18–24% of their nonexchangeable hydrogen from drinking water during biosynthesis of internal tissue biomass and 26–32% during biosynthesis of nail and feathers.
Figure 1.

Nonexchangeable δD values of proteinaceous tissues (A) (●, liver; ▴, blood; ▾, feather; ●, muscle) and lipids (B), from quail raised on two isotopically distinct diets and drinking-water sources (see Table 1 for specific diet and water δD values).
Similar to findings by Smith and Epstein (1), DeNiro and Epstein (9), and Estep and Hoering (10), lipids were more depleted in deuterium than proteinaceous tissues in all treatment groups (Table 1, Fig. 1b). Although hydrogen atoms in lipids are bound to carbon and therefore are not available for exchange with ambient metabolic or drinking water, the mean δD values of abdominal fat shifted by 57‰ (diet 1) and 62‰ (diet 2) with the different drinking water sources. Thus, drinking water contributed to the hydrogen composition of both nonlipid and lipid tissues in quail. Despite differences in protein content between diet 1 and diet 2, the influence of water on consumer tissue δD values was remarkably consistent for both diets.
In addition to mechanisms previously suspected of influencing δD values of the exchangeable portion of hydrogen (9), our measurement of the nonexchangeable hydrogen in both food and metabolically active and inactive tissues conclusively demonstrates that hydrogen from drinking water is incorporated into nonexchangeable sites in body proteins during or before protein synthesis. The hydrogen metabolic pathway responsible for this process is unknown but may involve exchange of hydrogen in drinking water with food macromolecules (particularly proteins and carbohydrates), both in the stomach and with body fluids before protein synthesis. Bird stomachs are particularly acidic, and this may promote greater enzymatic breakdown of larger molecules, thus providing increased steric access to H exchange sites.
Interestingly, the drinking water effect was most pronounced for feathers and hair, both metabolically inactive keratinous tissues. This suggests more opportunity for hydrogen exchange with body water during keratin or keratin precursor synthesis compared with the other body tissues we measured. Hydrogen in carbohydrates can also potentially exchange with drinking-water hydrogen and ultimately be fixed in nonexchangeable C—H bonds during conversion from carbohydrate to lipid. Experimental evidence for such a process was indicated by Jungas (18), who synthesized fatty acids in tritiated water and found hydrogen exchange between water and carbohydrate precursors to fatty acids. Incorporation of 3H from injected tritiated water into lipids has since been widely used as a method of measuring de novo synthesis of lipid (e.g., refs. 19–21). Evidence from these studies show that the incorporation of water elements into lipids is related to diet and nutritional status of an animal. This relationship further supports the suggestion that physiological ecology of organisms will be important when interpreting δD ratios in animal tissues.
Researchers using the doubly labeled water technique for metabolic studies (22) should be aware that the δD value of drinking water influences tissue δD values, including those in the blood, because an assumption of that technique is that injected labeled water is not incorporated into tissues (23). The time frame of doubly labeled water studies may be brief enough that the amount of water-derived hydrogen incorporated into tissues has only a negligible effect on the results; however, rapid exchange with water may occur at certain stages of fatty acid synthesis (18).
Our findings emphasizes that those studies using δD measurements of modern or fossil animal tissues for environmental interpretations may need to account for hydrogen isotopic differences that can occur between diet and drinking water. These findings are of particular concern to research using deuterium in animal tissues as an isotopic tracer in environmental studies. Animals that obtain drinking water from an isotopically different biome than where they obtain their food will have tissues with different δD values than species feeding and drinking in the same food web. For example, animals that live in marine habitats but drink freshwater (24), or animals living in arid regions where open water has undergone extensive evaporation and enrichment in deuterium, may show a departure from expected tissue δD values if only the δD values of their food is considered. Thus, comparison of isotopic data from animal remains may require an understanding of the local feeding and drinking habits of the organism, because isotopic differences over time may reflect a change in feeding and drinking behavior rather than climate. However, the recent application of δD measurements to trace natal origins of migrant insects (17, 25) illustrates one of many applications where drinking water is not a complicating variable, because the insect wings were formed ultimately only from a single species of plant material and were metabolically inactive after formation.
Our findings show that the application of simple diet-tissue δD fractionation factors for various tissue types cannot be generally assumed or applied in ecological studies aiming to reconstruct diet or climate. However, although water did contribute to tissue hydrogen isotopic composition, its influence was remarkably consistent among tissue types and was largely independent of diet. Further work aimed at modeling the empirical relationship between naturally occurring isotopic values for diet, water, and tissue is needed.
Acknowledgments
Laboratory assistance was provided by Brigitte Boldt-Leppin, Steve VanWilgenburg, and Randy George. We thank three anonymous reviewers for their valuable comments on an earlier draft of this paper. Funding was provided by the Canadian Wildlife Service and the National water Research Institute of Environment Canada. Financial assistance (L.A.) was also provided by a University of Saskatchewan Graduate Scholarship.
ABBREVIATION
- δD
stable-hydrogen isotope ratio
Appendix
A significant proportion, fe, of the total hydrogen in organic tissues may be subject to isotopic exchange with ambient moisture. This process involves a temperature-dependent isotopic fractionation factor (α) between the exchangeable organic hydrogen and ambient water (δDw) whereby the stable isotope value for the tissue, δDt, can be expressed as:
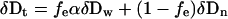 |
1 |
and δDn represents the isotopic value of the nonexchangeable hydrogen (26). A series of isothermal isotopic equilibrations with steam yield a straight line relationship between δDw and δDt, the slope being a function of fe and the equilibrium fractionation factor α (16, 26). Unfortunately, α cannot be easily determined for complex organic matrices, and so, in our calculations of exchangeability, we allowed it to range between 1.060 and 1.100, typical of many complex organics (ref. 16; A. Schimmelmann, personal communication).
A simple two end-member isotopic equilibration procedure was used to calculate the proportion of exchangeability for tissues based on tissue δD values equilibrated with various waters.
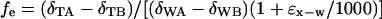 |
2 |
where the A and B subscripts refer to widely separated equilibration waters (A. Schimmelmann, personal communication). We used three pairs of static equilibration experiments for each tissue type to determine the average fe (e.g., −135‰ vs. +525‰, +312‰ vs. +525‰, −135‰ vs. +312‰), and, corresponding to the range of values considered for α above, ɛx−w, the equilibrium isotopic fractionation factor, expressed in parts per thousand, was allowed to range between 60 and 100. This provided a series of estimates for fe that were ultimately averaged for our use in deriving δDn. For example, using this experimental approach and Eq. 2, we derived the following estimates of exchangeability: whole freeze-dried blood, 17.0 ± 0.5%; muscle, 19.5 ± 0.4%; liver, 18.6 ± 1.5%; lipid, 0%; feather, 18.6 ± 3.9%; and quail food, 20.7 ± 0.5%. All results in Table 1 are reported as δDn from Eq. 1.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
References
- 1.Smith B N, Epstein S. Plant Physiol. 1970;46:738–742. doi: 10.1104/pp.46.5.738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Estep M F, Dabrowski H. Science. 1980;209:1537–1538. doi: 10.1126/science.209.4464.1537. [DOI] [PubMed] [Google Scholar]
- 3.Schimmelmann A, DeNiro M J. Geochim Cosmochim Acta. 1986;50:1485–1496. [Google Scholar]
- 4.Miller R F, Fritz P, Morgan A V. Palaeogeogr Palaeoclimatol Palaeoecol. 1988;66:277–288. [Google Scholar]
- 5.Cormie A B, Schwarcz H P, Gray J. Geochim Cosmochim Acta. 1994;58:377–391. [Google Scholar]
- 6.Cormie A B, Luz B, Schwarcz H P. Geochim Cosmochim Acta. 1994;58:3439–3449. [Google Scholar]
- 7.Chamberlain C P, Blum J D, Holmes R T, Feng X, Sherry T W, Graves G R. Oecologia. 1997;109:132–141. doi: 10.1007/s004420050067. [DOI] [PubMed] [Google Scholar]
- 8.Hobson K A, Wassenaar L I. Oecologia. 1997;109:142–148. doi: 10.1007/s004420050068. [DOI] [PubMed] [Google Scholar]
- 9.DeNiro M, Epstein S. Science. 1981;214:1374–1376. doi: 10.1126/science.7313700. [DOI] [PubMed] [Google Scholar]
- 10.Estep M F, Hoering T C. Geochim Cosmochim Acta. 1980;44:1197–1206. [Google Scholar]
- 11.Schoeller D A, Minagawa M, Slater R, Kaplan I R. Ecol Food Nutr. 1986;18:159–170. [Google Scholar]
- 12.Schimmelmann A, Miller R F, Leavitt S W. Geophys Monogr. 1993;78:367–374. [Google Scholar]
- 13.Canadian Council on Animal Care. Guide to the Care and Use of Experimental Animals. Vol. 2. Canadian Council on Animal Care; 1984. [Google Scholar]
- 14.Dobush G R, Ankney C D, Krementz D G. Can J Zool. 1985;63:1917–1920. [Google Scholar]
- 15.Epstein S, Yapp C J, Hall J H. Earth Planet Sci Lett. 1976;30:241–251. [Google Scholar]
- 16.Schimmelmann A. Anal Chem. 1991;63:2456–2459. [Google Scholar]
- 17.Wassenaar L I, Hobson K A. Proc Nat Acad Sci USA. 1998;95:15436–15439. doi: 10.1073/pnas.95.26.15436. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Jungas R L. Biochemistry. 1968;7:3708–3717. doi: 10.1021/bi00850a050. [DOI] [PubMed] [Google Scholar]
- 19.Goldman J K, Bernardis L L, Kodis M. Metabolism. 1978;27:2006–2008. doi: 10.1016/s0026-0495(78)80016-x. [DOI] [PubMed] [Google Scholar]
- 20.Cascales C, Benito M, Cascales M, Caldes T, Santos-Ruiz A. Br J Nutr. 1983;50:549–553. doi: 10.1079/bjn19830126. [DOI] [PubMed] [Google Scholar]
- 21.Dunshea F R, Bell A W, Trigg T E. Br J Nutr. 1988;60:633–644. doi: 10.1079/bjn19880133. [DOI] [PubMed] [Google Scholar]
- 22.Nagy K A. In: Stable Isotopes in Ecological Research. Rundel P W, Ehleringer J R, Nagy K A, editors. New York: Springer; 1989. pp. 270–287. [Google Scholar]
- 23.Tatner P, Bryant D M. In: Techniques in Comparative Respiratory Physiology. Bridges C R, Butler P J, editors. New York: Cambridge Univ. Press; 1989. [Google Scholar]
- 24.Woodin M C. Hydrobiologia. 1994;279–280:279–287. [Google Scholar]
- 25.Hobson, K. A., Wassenaar, L. I. & Taylor, O. R. (1999) Oecologia, in press. [DOI] [PubMed]
- 26.Grinsted M J, Wilson A T. N Z J Sci. 1979;22:281–287. [Google Scholar]