

Abstract
Endogenous synthesis of oxalate is an important contributor to calcium oxalate stone formation and renal impairment associated with primary hyperoxaluria. Although the principal precursor of oxalate is believed to be glyoxylate, pathways in humans resulting in glyoxylate synthesis are not well defined. Hydroxyproline, a component amino acid of collagen, is a potential glyoxylate precursor. We have investigated the contribution of dietary hydroxyproline derived from gelatin to urinary oxalate and glycolate excretion. Responses to the ingestion of 30 g of gelatin or whey protein were compared on controlled oxalate diets. The time course of metabolism of a 10 g gelatin load was determined as well as the response to varying gelatin loads. Urinary glycolate excretion was 5.3-fold higher on the gelatin diet compared to the whey diet and urinary oxalate excretion was 43% higher. Significant changes in plasma hydroxyproline and urinary oxalate and glycolate were observed with 5 and 10 g gelatin loads, but not 1 and 2 g loads. Extrapolation of these results to daily anticipated collagen turnover and hydroxyproline intake suggests that hydroxyproline metabolism contributes 20−50% of glycolate excreted in urine and 5−20% of urinary oxalate derived from endogenous synthesis. Our results also revealed that the kidney absorbs significant quantities of hydroxyproline and glycolate, and their metabolism to oxalate in this tissue warrants further consideration.
Keywords: hydroxyproline, oxalate, glycolate, urolithiasis, kidney
The most common form of kidney stone is composed of calcium oxalate. Oxalate excreted in urine is derived in approximately equal parts from the diet and endogenous synthesis.1 An increased endogenous production of oxalate in the primary hyperoxalurias (PHs) not only dramatically increases stone formation, but can in some individuals lead to tissue deposition of calcium oxalate and renal failure. The metabolic pathways that result in oxalate synthesis remain unclear despite this clinical significance. In Type 1 and Type 2 PH, deficiencies in the glyoxylate metabolizing enzymes, alanine:glyoxylate aminotransferase (AGT) and glyoxylate reductase, respectively, result in excessive oxalate synthesis.2 The sources of the glyoxylate, important in both Type 1 and Type 2 disease, have not been clearly identified. In Type 1 disease, the glyoxylate derived from glycolate through the activity of glycolate oxidase localized in liver peroxisomes is thought to be the major source of oxalate synthesis. Sources of the glycolate, however, have not been identified. Glyoxylate reductase activity, present in both mitochondria and the cytoplasm, may be an important source as it reduces glyoxylate to glycolate.
Hydroxyproline metabolism occurs principally in mitochondria of hepatocytes and renal proximal tubule cells, and results in the formation of equal amounts of pyruvate and glyoxylate.3,4 The daily turnover of collagen in humans is estimated to be 2−3 g/day,4 leading to the metabolism of 240−420 mg of hydroxyproline and the formation of 140−240 mg of glyoxylate. Additional hydroxyproline may be obtained from the diet, primarily through the ingestion of collagen-containing meat products and gelatin-containing foods. The ingestion of large amounts of hydroxyproline increases the urinary excretion of oxalate and glycolate in rats5–7 and pigs.8 The response of humans to large doses of hydroxyproline is not known. This study was designed to determine how human subjects metabolize hydroxyproline derived from gelatin, as measured by changes in the urinary excretion of oxalate and glycolate. An understanding of the contribution of hydroxyproline metabolism to oxalate is required to determine whether blocking this pathway would be useful in limiting endogenous oxalate synthesis in individuals with PH or calcium oxalate stone disease.
RESULTS
The mean circulating concentration of hydroxyproline in individuals when consuming self-selected diets was 13.2±5.7 μM (Table 1). A broad range of values was detected spanning 4.8−23.8 μM, suggesting inter-individual differences in collagen turnover, hydroxyproline metabolism, and/or hydroxyproline intake. With the ingestion of 30 g of gelatin containing 2.75 g hydroxyproline each day for 3 days, hydroxyproline concentrations increased to a mean value of 117.9±21.1 μM on the second day and 132.1±20.7 μM on the third day. Glycine plasma concentration was significantly higher following ingestion of the gelatin compared to the whey protein (Table 1). In contrast, alanine and serine did not show significant changes compared to whey protein (data not shown).
Table 1.
| Mean 24 h urine oxalate, urine glycolate, and plasma hydroxyproline after consuming either self-selected diets or diets containing either 30 g of gelatin or whey protein per day
|
Diet |
||||
|---|---|---|---|---|
| Self-selected | Whey | Gelatin | P-value (gelatin vs whey) | |
| Oxalate (mg/g creatinine) | 17.6±5.2 (12.2−26.1) | 17.2±3.9 (10.5−23.7) | 24.4±4.6 (17.0−33.0) | <0.001 |
| Glycolate (mg/g creatinine) | 19.9±5.7 (12.2−34.9) | 14.9±3.8 (5.0−21.8) | 78.3±33.1 (35.8−166.8) | <0.001 |
| Plasma hydroxyproline (μM) | 13.2±5.7 (4.8−23.8) | 14.6±4.5 (8.8−24.0) | 125.0±21.5 (97.8−162.2) | <0.001 |
| Plasma glycine (μM) | ND | 217.3±69.6 (85.3−365.1) | 515.8±212.9 (306.1−958.5) | <0.001 |
ND, not determined.
The urine results are the mean 24 h excretions on days 2 and 3 of the protein diets or on 3 days of a self-selected diet. Plasma values are the mean values of samples obtained 4 h after breakfast on days 2 and 3 of the protein diets or on 2 days of a self-selected diet. Ranges are indicated in parentheses.
The mean oxalate and glycolate excretions following ingestion of gelatin were significantly higher than those with whey protein (Table 1). Glycolate excretion increased by 5.3-fold and oxalate excretion increased by 43%. The difference in glycolate and oxalate excretions on gelatin and whey diets indicated that on a molar basis, approximately 5% of the hydroxyproline metabolized was excreted as glycolate and 0.5% as oxalate. The urinary excretion of hydroxyproline increased from 3.6±0.2 mg/day on day 3 of the whey diet to 14.2±5.2 mg/day on day 3 of the gelatin diet.
An analysis of the time course of metabolism of a 10 g gelatin load revealed significant changes in plasma glycolate over time that mirrored those of plasma hydroxyproline. Both parameters peaked at 3 h and were still 33% greater than fasting levels 24 h post load (Figure 1). Plasma oxalate levels were highest at 3 h, but were not significantly different from the fasting values (Figure 1). Urinary glycolate increased significantly post load reaching a peak in the 4−6 h collection, where glycolate levels were approximately eightfold greater than fasting values (Figure 2). Urinary oxalate was significantly higher at all post load collections compared to fasting levels and reached a plateau at the 2−8 h collections (Figure 2). The ratio of glycolate clearance relative to creatinine clearance was calculated for each timed collection so as to provide some insight into how the kidney handles filtered glycolate. The mean clearance ratio for collection periods from 0−8 h post 10 g gelatin load was 0.63±0.21, indicating a net reabsorption of glycolate (Figure 3).
Figure 1. Change in plasma hydroxyproline (inset), glycolate (■), and oxalate (–▲–) after ingestion of 10 g gelatin.
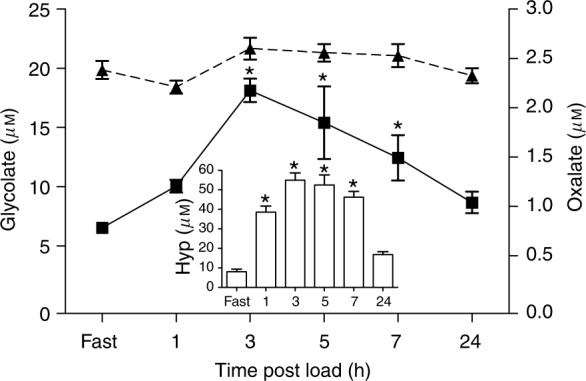
Each time point represents the mean±s.e.m. *P<0.05 for the comparison of plasma glycolate or hydroxyproline at each post load collection time with the corresponding fasting value.
Figure 2. Urinary glycolate (■) and urinary oxalate (–▲–) following ingestion of 10 g gelatin.
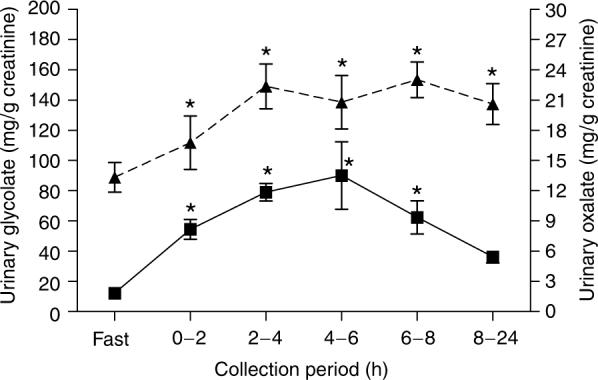
Each collection represents the mean±s.e.m. *P<0.05 for the comparison of urinary glycolate or oxalate at each post load collection period with the corresponding fasting value.
Figure 3. The ratio of glycolate clearance to creatinine clearance following 10 g load of gelatin.
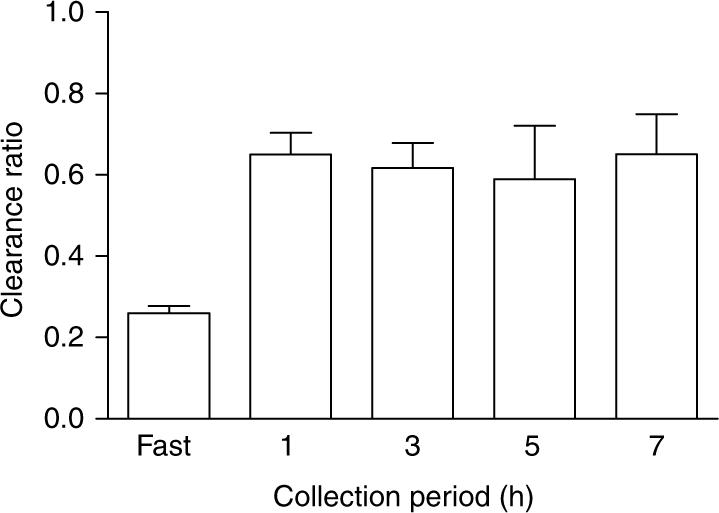
Each collection represents the mean±s.e.m.
The relationship between the amount of hydroxyproline ingested and oxalate and glycolate excretion in a 6-h period was assessed following loads containing 1−10 g of gelatin. The results shown in Figure 4 indicate that there was a proportional relationship between urinary oxalate and glycolate excretion and the amount of gelatin ingested, but significant increases in oxalate and glycolate excretion were not observed at the lower gelatin doses. This finding was in keeping with plasma hydroxyproline measurements, which did not show significant increases at the lower gelatin doses (Figure 4).
Figure 4. Increases in urinary oxalate (■) and glycolate (□) excretion over fasting levels in a 6-h period following ingestion of various loads of gelatin.
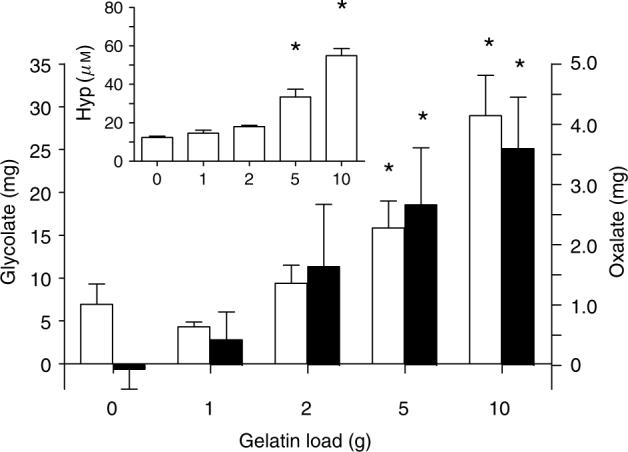
Increases in urinary oxalate and glycolate excretion were calculated by subtracting three times the fasting 2 h levels from the total amount measured in each collection. Plasma hydroxyproline (inset) was measured 3 h after ingestion of each load. The asterisk denotes significantly higher (P<0.05) values compared to the 0 g gelatin load. Each parameter represents the mean±s.e.m.
DISCUSSION
The mean circulating concentration of hydroxyproline we determined in individuals on self-selected diets, 13.2 μM, is similar to other reported mean values in the range of 10−15 μM.9,10 Although collagen turnover in the body is a major source of hydroxyproline, significant amounts of hydroxyproline may also be ingested in meats and gelatin-containing foods. Ground beef may contain up to 7% collagen, for instance.11 Collagen supplements are also promoted for the health of skin, bone, and joints, and as a weight loss agent, and may be a significant source of hydroxyproline for some individuals. If there is a linear relationship between the amount of hydroxyproline metabolized and the amounts of glycolate and oxalate produced, and if 2−5 g of collagen/gelatin are broken down each day, the results in Figure 4 suggest that this will result in the production of approximately 1−3 mg oxalate and 10−20 mg of glycolate per day. This could account for 20−50% of the glycolate excreted in urine and 5−20% of the endogenous oxalate produced in the body each day, if it is assumed that 50% of urinary oxalate, ∼15 mg/day, is derived from endogenous synthesis.
The possible fates of glyoxylate produced from the metabolism of hydroxyproline are illustrated in Figure 5. The steps shown apply to human liver tissue where AGT1 is localized in peroxisomes. In other species, including the rat and mouse, AGT1 may also be localized in mitochondria. The dual localization in these rodents and a much lower AGT1 activity compared to human tissue suggests that caution must be exercised when extrapolating any results of experiments involving glyoxylate metabolism in these species to humans. Kidney and heart tissue also possess the enzymes required for hydroxyproline breakdown.4,12,13 As only 5% of the ingested hydroxyproline was recovered in urine as glycolate and oxalate, the bulk of the glyoxylate produced from hydroxyproline metabolism was most likely converted to glycine. Although plasma glycine had doubled after ingestion of gelatin, the glycine produced from hydroxyproline metabolism could not be determined as gelatin contains high amounts of glycine. The absence of AGT1 activity in individuals with PH1 provides further insight into the magnitude of this pathway as the conversion of glyoxylate to glycine is blocked. The combined urinary excretion of glycolate and oxalate in these individuals is usually in the range of 200−500 mg/day. If daily collagen turnover is indeed 2−3 g/day and the ingestion of some hydroxyproline occurs, hydroxyproline metabolism may contribute to the bulk of their oxalate and glycolate excretion. The combined excretion of oxalate and glycolate is much less in individuals with PH2, implying that AGT1 may convert a significant amount of the glyoxylate produced from hydroxyproline metabolism to glycine.
Figure 5. Hydroxyproline transport and metabolism in hepatocytes.
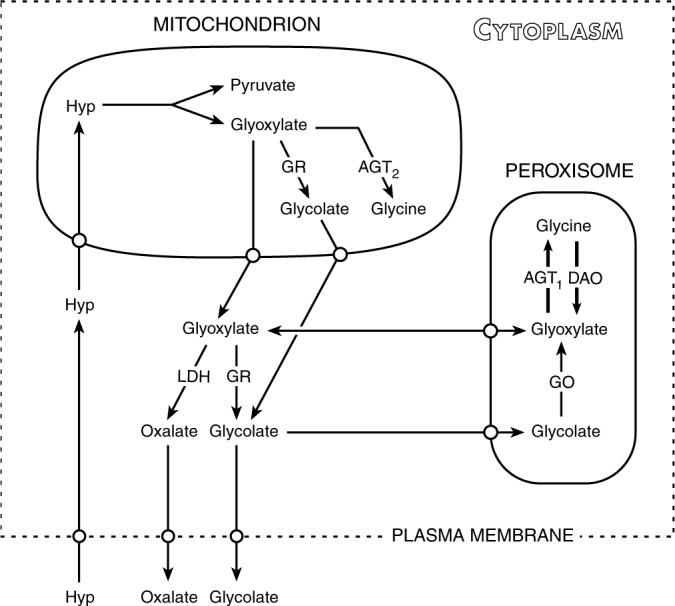
Hyp, trans-4-hydroxy-L-proline; GR, glyoxylate reductase; DAO, D-amino oxidase; AGT, alanine:glyoxylate aminotransferase; GO, glycolate oxidase; LDH, lactate dehydrogenase.
Of interest also is the potential renal metabolism of hydroxyproline. If the circulating hydroxyproline concentration is 10 μM and the glomerular filtration rate is 100 ml/min, 188 mg of hydroxyproline would be filtered each day. Following ingestion of 30 g of whey protein, mean urinary hydroxyproline was only 4 mg per day. This is in close agreement with a previous study, which reported that <5 mg hydroxyproline is normally excreted in urine per day.14 These data indicate that the bulk of filtered hydroxyproline is reabsorbed by the kidney. Furthermore, our data showed that plasma hydroxyproline following the ingestion of 30 g gelatin increased to approximately 100 μM. Thus, the filtered load on the kidney would increase to 1.88 g/day. Urine hydroxyproline measurements showed that only 14 mg hydroxyproline was excreted following ingestion of 30 g gelatin. This indicates that even at 10 times the normal filtered load of hydroxyproline per day, the kidney reabsorbs over 99%. Some hydroxyproline may be metabolized in the renal cortex as it contains enzymes in the hydroxyproline degradation pathway.12 Based on the relative weights of the tissues and their activities, it might be expected that 15% of the hydroxyproline is metabolized in the renal cortex. The kidney contains glyoxylate reductase activity and can therefore convert hydroxyproline-derived glyoxylate to glycolate.15,16 The organ lacks AGT1 activity, but does contain AGT2 activity, which has a much lower affinity for glyoxylate than AGT1.17 Thus, in comparison to the liver, the kidney may have a reduced capacity to convert glyoxylate to glycine. AGT2 has no homology with AGT1, is found in mitochondria of most tissues, and has D-3-aminobutyrate:pyruvate aminotransferase, β-alanine:pyruvate aminotransferase and dimethylarginine:pyruvate aminotransferase activities, as well as AGT activity. Lowry et al.12 reported that isolated rat proximal tubules can convert hydroxyproline-derived glyoxylate to glycine. Whether any glyoxylate is converted to oxalate and whether the kidney is a significant source of endogenous oxalate synthesis remains to be determined. Much of the hydroxyproline reabsorbed by the kidney may return to the circulation, as hydroxyproline levels were 33% higher than the original fasting levels 24 h after the ingestion of 10 g gelatin. These results also suggest that this load of gelatin exceeded the body's capacity to rapidly metabolize the free hydroxyproline it produced.
The studies we have performed also provide some insight into glycolate production, its clearance, and its potential renal metabolism. We have previously reported that an average 39 mg of glycolate is excreted by individuals on self-selected diets.18 This is in close agreement to this study, where a mean of 35 mg glycolate was excreted by study subjects per day when on self-selected diets. Harris and Richardson19 estimated that the mean daily glycolate intake is 33 mg. They further provided evidence that glycolate is efficiently absorbed and metabolized in rats, suggesting that the glycolate excreted in human urine was derived predominantly from the diet. Their method for glycolate analysis in food extracts utilized a nonspecific colorimetric procedure. Analyses in our laboratory by ion chromatography coupled with electrospray mass spectrometry detection of mushrooms, peaches, and coffee, which Harris and Richardson found to contain some of the highest glycolate levels of the foods they tested, indicated that their values were overestimated 25- to 50-fold (J Knight and RP Holmes, unpublished results). Our results indicated that in fasting subjects, the mean circulating glycolate concentration was 6.5 μM, and reached 10 μM 4 h after a meal. We speculate that the bulk of this glycolate is derived from metabolism and not from the diet. Assuming an average plasma glycolate concentration of 7.5 μM during the day and a glomerular filtration rate of 100 ml/min, 82 mg is filtered each day. It appears, therefore, that half of the filtered glycolate is reabsorbed and presumably metabolized, and the other half excreted in urine. The average clearance ratio of 0.6 in these studies supports these calculations. The higher clearance ratio reported here is likely due to the increased glycolate load on the kidney from ingestion of hydroxyproline and its metabolism to glycolate. Whether this glycolate is oxidized in part to oxalate warrants further investigation. We have observed that both rat and human renal tissue contain glycolate oxidase activity that may be due to the presence of large amounts of HAOX2 (long-chain hydroxy-fatty acid oxidase), which has some affinity for glycolate (Holmes RP, unpublished results).
Whereas these studies indicate that hydroxyproline metabolism is likely to be a significant source of glyoxylate production in humans, a more definitive picture of the contribution of hydroxyproline metabolism to daily glycolate and oxalate synthesis may only emerge by following the metabolism of isotopic hydroxyproline. Better quantitative evidence is required to determine if blocking this metabolic pathway would be useful in limiting oxalate synthesis in patients with PH or calcium oxalate stone disease.
MATERIALS AND METHODS
Subjects
Healthy subjects with no history of nephrolithiasis were recruited from Wake Forest University Medical Center. Subjects provided informed consent before participating in this study, which was approved by the Institutional Review Board. All subjects had a body mass index <27. In addition, they were not taking any medications or dietary supplements and refrained from vigorous exercise.
Gelatin/whey study
Ten subjects (five male and five female subjects; mean age, 32±4 years; age range, 28−39 years) participated in the study. Subjects were initially asked to collect three consecutive 24 h urines while consuming self-selected diets. On the last 2 days of this portion of the study, blood was drawn 4 h after breakfast. Following these collections, subjects consumed diets controlled in its content of calories, fat, protein, carbohydrate, calcium, magnesium, sodium, phosphorus, and oxalate, as described previously,1 for two 5-day periods separated by at least a 1-week washout period. During one 5-day period, the diet contained a quarter of its protein as gelatin (Knox). The second 5-day period of the diet contained a quarter of its protein as whey. The hydroxyproline content of the gelatin, measured by AAA Laboratory (Mercer Island, WA, USA), was 9.17 g per 100 g, and the whey protein was free of hydroxyproline. Ten grams of either gelatin or whey protein were provided as dry powders mixed in with cranberry juice with each meal. On the final 3 days of the gelatin and whey diet period, each subject provided a blood sample 4 h after breakfast and collected 24 h urines. Controlled diets were prepared in the metabolic kitchen of the institutional General Clinical Research Center. The calcium content was targeted at 1000 mg/2500 kcal and the oxalate content 150 mg/2500 kcal. This oxalate content is within the reported range of normal oxalate intake.20 Diets were adjusted to within 5% of these amounts, and with a ratio of calcium to oxalate in each meal >5:1. Each diet contained 15−16% protein, 29−30% fat, and 54−55% carbohydrate. To closely control the volume of urine, subjects drank only the fluids provided, which included 2 l of bottled water per day. Sodium, calcium, magnesium, phosphate, uric acid, citrate, and urea were measured in urine samples as described previously.18 These measurements provided a means of assessing subject compliance with the dietary protocol.
Gelatin-loading studies
Six subjects (three male and three female subjects; mean age, 28±7 years; age range, 23−42 years) were recruited for this study to investigate the effect of varying gelatin loads on glycolate and oxalate in plasma and urine, and the time course of metabolism of hydroxyproline over 24 h following a 10 g gelatin load. Throughout this study subjects were on self-selected diets, but were required to avoid oxalate-rich foods and eat moderate amounts of protein and calcium. The day before each load, subjects fasted overnight (14 h). A fasting 2 h urine collection was obtained with a blood sample at the midpoint of this collection. Gelatin was provided as a dry powder mixed with 6 ounces of yogurt and was consumed at breakfast along with a sausage biscuit and 250 ml water. Gelatin loads of 0, 1, 2, 5, and 10 g of gelatin were tested. Each load was separated by at least 2 days. Urine was collected for 6 h and blood was drawn 3 h after ingestion of each load. The time course of changes in plasma hydroxyproline, oxalate, and glycolate levels and the urinary excretion of oxalate and glycolate was examined on a 10 g load. Four sequential 2 h urine collections were obtained after the 10 g load together with a blood sample at the midpoint of each collection. Over the next 16 h, subjects obtained an evening collection (6 h) and another in the morning of the following day (10 h). Subjects were required to drink 250 ml bottled water per hour over the 6-h period following gelatin loads, and the initial 8-h period after the 10 g load to ensure good urine output.
Plasma and urine analyses
Blood samples were obtained by venipuncture into heparinized tubes and immediately centrifuged at 4°C for 10 min (1500 g). If not analyzed the same day, plasma samples were stored at −80°C. Plasma was filtered using acid washed Ultrafree-MC centrifugal filters (Millipore Corp., Bedford, MA, USA) with a 10000 nominal molecular weight limit. Twenty-four urine collections were collected in boric acid. For oxalate analysis, urines were diluted in 2 mM HCl before storage at −80°C. Total urine oxalate and plasma oxalate were determined by ion chromatography, as described previously.21,22
Urine and plasma glycolate were determined by ion chromatography coupled with electrospray mass spectrometry (Dionex Corp., Sunnyvale, CA, USA). The ion chromatography equipment consisted of a Dionex ED50 conductivity detector, Dionex AS11_HC 2 × 250-mm ion exchange column with guard column at a controlled temperature of 30°C, and a Dionex ASRS-ULTRA 2-mm suppressor. A sodium hydroxide gradient 0.5−1 mM over 25 min at a flow rate of 0.4 ml/min was used to separate glycolate from the other sample anions, which enabled sensitive quantitation of glycolate by mass detection. The limit of detection, defined as the mean blank signal plus 10 times the s.d. of the blank, was 1 pmole (0.2 μM). A typical calibration curve for glycolate measured by mass detection is shown in Figure 6. A linear response was obtained up to 10 μM. The intra-sample coefficient of variation (n = 6) for the measurement of glycolate in urine and plasma was 4.1 and 6.1%, respectively. The mean recovery of 10 μM glycolate added to four different plasma samples was 105±5%, range 101−110%. The mean recovery of 50 μM glycolate added to four different urine samples was 103±7.5%, range 99−112%, respectively. Amino-acid analysis was performed by the AccQ.Tag method (Waters Corp., Milford, MA, USA), as described previously.23
Figure 6.

| Calibration curve for glycolate determination by ion chromatography coupled with electrospray mass spectrometry.
Statistical analyses
Results are presented as mean values±s.d., unless otherwise indicated. The statistical significance of results was assessed by analysis of variance and considered significant if P-values were 0.05 or less. The Holm–Sidak method was used to correct for multiple comparisons and all assays were conducted using SigmaStat (Systat Software Inc., Richmond, CA, USA).
ACKNOWLEDGMENTS
This research was supported by NIH Grants RO1 DK54468, RO1 DK62284, and MO1 RR07122. The assistance of Linda Easter in the design and preparation of diets was greatly appreciated. We gratefully acknowledge the invaluable contribution of Martha Kennedy and Persida Tahiri for assisting with these experiments.
REFERENCES
- 1.Holmes RP, Goodman HO, Assimos DG. Contribution of dietary oxalate to urinary oxalate excretion. Kidney Int. 2001;59:270–276. doi: 10.1046/j.1523-1755.2001.00488.x. [DOI] [PubMed] [Google Scholar]
- 2.Danpure CJ. Primary hyperoxaluria. In: Scriver CR, Beaudet AL, Sly WS, Vallee D, Childs B, Kinzler KW, Vogelstein B, editors. The Metabolic and Molecular Bases of Inherited Disease. 8th edn McGraw-Hill; New York: 2001. pp. 3323–3367. [Google Scholar]
- 3.Knight J, Holmes RP. Mitochondrial hydroxyproline metabolism: implications for primary hyperoxaluria. Am J Nephrol. 2005;25:171–175. doi: 10.1159/000085409. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Phang JM, Hu CA, Valle D. Disorders of proline and hydroxyproline metabolism. In: Scriver CR, Beaudet AL, Sly WS, Vallee D, Childs B, Kinzler KW, Vogelstein B, editors. The Metabolic and Molecular Bases of Inherited Disease. McGraw-Hill; New York: 2001. pp. 1821–1838. [Google Scholar]
- 5.Bushinsky DA, Asplin JR, Grynpas MD, et al. Calcium oxalate stone formation in genetic hypercalciuric stone-forming rats. Kidney Int. 2002;61:975–987. doi: 10.1046/j.1523-1755.2002.00190.x. [DOI] [PubMed] [Google Scholar]
- 6.Takayama T, Fujita K, Suzuki K, et al. Control of oxalate formation from l-hydroxyproline in liver mitochondria. J Am Soc Nephrol. 2003;14:939–946. doi: 10.1097/01.asn.0000059310.67812.4f. [DOI] [PubMed] [Google Scholar]
- 7.Khan SR, Glenton PA, Byer KJ. Modeling of hyperoxaluric calcium oxalate nephrolithiasis: experimental induction of hyperoxaluria by hydroxy-l-proline. Kidney Int. 2006;70:914–923. doi: 10.1038/sj.ki.5001699. [DOI] [PubMed] [Google Scholar]
- 8.Mandel NS, Henderson JD, Hung LY, et al. A porcine model of calcium oxalate kidney stone disease. J Urol. 2004;171:1301–1303. doi: 10.1097/01.ju.0000110101.41653.bb. [DOI] [PubMed] [Google Scholar]
- 9.Adams E, Frank L. Metabolism of proline and the hydroxyprolines. Ann Rev Biochem. 1980;9:1005–1061. doi: 10.1146/annurev.bi.49.070180.005041. [DOI] [PubMed] [Google Scholar]
- 10.Inoue H, Iguch H, Kouno A, et al. Fluorometric determination of N-terminal prolyl dipeptides, proline and hydroxyproline in human serum by pre-column high performance liquid chromatography using 4-(5,6-dimethoxy-2-phthalimidinyl)-2-methoxyphenylsulfonyl chloride. J Chromatogr. 2001;57:369–373. doi: 10.1016/s0378-4347(01)00162-1. [DOI] [PubMed] [Google Scholar]
- 11.Wold JP, Lundby F, Egelandsdel B. Quantification of connective tissue (hydroxyproline) in ground beef by autofluorescence spectroscopy. J Food Sc. 1999;4:377–383. [Google Scholar]
- 12.Lowry M, Hall DE, Brosnan JT. Hydroxyproline metabolism by the rat kidney: distribution of renal enzymes of hydroxyproline catabolism and renal conversion of hydroxyproline to glycine and serine. Metabolism. 1985;4:955–961. doi: 10.1016/0026-0495(85)90145-3. [DOI] [PubMed] [Google Scholar]
- 13.Hu C, Lin WW, Steel G, et al. Identification of the gene encoding hydroxyproline oxidase and delineation of mutations responsible for hyperhydroxyprolinemia. Am J Hum Gen. 2001;69(Suppl 1):1708. [Google Scholar]
- 14.Kivirikko KI. Urinary excretion of hydroxyproline in health and disease. Int Rev Connect Tiss Res. 1970;5:93–163. doi: 10.1016/b978-0-12-363705-5.50008-7. [DOI] [PubMed] [Google Scholar]
- 15.Cramer SD, Ferree PM, Lin K, et al. The gene encoding hydroxypyruvate reductase (GRHPR) is mutated in patients with primary hyperoxaluria Type II. Hum Mol Genet. 1999;11:2063–2069. doi: 10.1093/hmg/8.11.2063. [DOI] [PubMed] [Google Scholar]
- 16.Giafi CF, Rumsby G. Kinetic analysis and tissue distribution of human d-glycerate dehydrogenase/glyoxylate reductase and its relevance to the diagnosis of primary hyperoxaluria type 2. Ann Clin Biochem. 1998;35:104–109. doi: 10.1177/000456329803500114. [DOI] [PubMed] [Google Scholar]
- 17.Lee ISM, Muragaki Y, Ideguchi T, et al. Molecular-cloning and sequencing of a cDNA-encoding alanine-glyoxylate aminotransferase 2 from rat-kidney. J Biochem. 1995;117:856–862. doi: 10.1093/oxfordjournals.jbchem.a124787. [DOI] [PubMed] [Google Scholar]
- 18.Holmes RP, Goodman HO, Hart LJ, et al. Relationship of protein intake to urinary oxalate and glycolate excretion. Kidney Int. 1993;44:366–372. doi: 10.1038/ki.1993.253. [DOI] [PubMed] [Google Scholar]
- 19.Harris KS, Richardson KE. Glycolate in the diet and its conversion to urinary oxalate in the rat. Invest Urol. 1980;18:106–109. [PubMed] [Google Scholar]
- 20.Holmes RP, Kennedy M. Estimation of the oxalate content of foods and daily oxalate intake. Kidney Int. 2000;57:1662–1667. doi: 10.1046/j.1523-1755.2000.00010.x. [DOI] [PubMed] [Google Scholar]
- 21.Hagen L, Walker VR, Sutton RA. Plasma and urinary oxalate and glycolate in healthy subjects. Clin Chem. 1993;39:134–138. [PubMed] [Google Scholar]
- 22.Holmes RP, Ambrosius WT, Assimos DG. Dietary oxalate loads and renal oxalate handling. J Urol. 2005;174:943–947. doi: 10.1097/01.ju.0000169476.85935.e2. [DOI] [PubMed] [Google Scholar]
- 23.Reverter M, Lundh T, Lindberg JE. Determination of free amino acids in pig plasma by precolumn derivatization with 6-Naminoquindyl-N-hydroxysuccinimidyl carbamate and high performance liquid chromatography. J Chromatogr B. 1997;696:1–8. doi: 10.1016/s0378-4347(97)00217-x. [DOI] [PubMed] [Google Scholar]