

Abstract
Homologues of the amtB gene of enteric bacteria exist in all three domains of life. Although their products are required for transport of the ammonium analogue methylammonium in washed cells, only in Saccharomyces cerevisiae have they been shown to be necessary for growth at low NH4+ concentrations. We now demonstrate that an amtB strain of Escherichia coli also grows slowly at low NH4+ concentrations in batch culture, but only at pH values below 7. In addition, we find that the growth defect of an S. cerevisiae triple-mutant strain lacking the function of three homologues of the ammonium/methylammonium transport B (AmtB) protein [called methylammonium/ammonium permeases (MEP)] that was observed at pH 6.1 is relieved at pH 7.1. These results provide direct evidence that AmtB participates in acquisition of NH4+/NH3 in bacteria as well as eucarya. Because NH3 is the species limiting at low pH for a given total concentration of NH4+ + NH3, results with both organisms indicate that AmtB/MEP proteins function in acquisition of the uncharged form. We confirmed that accumulation of [14C]methylammonium depends on its conversion to γ-N-methylglutamine, an energy-requiring reaction catalyzed by glutamine synthetase, and found that at pH 7, constitutive expression of AmtB did not relieve the growth defects of a mutant strain of Salmonella typhimurium that appears to require a high internal concentration of NH4+/NH3. Hence, contrary to previous views, we propose that AmtB/MEP proteins increase the rate of equilibration of the uncharged species, NH3, across the cytoplasmic membrane rather than actively transporting—that is, concentrating—the charged species, NH4+.
The AmtB (ammonium/methylammonium transport B) protein of enteric bacteria and its homologues [e.g., the MEP (methylammonium/ammonium permease) proteins of Saccharomyces cerevisiae] are predicted to be cytoplasmic membrane proteins with a number of transmembrane-spanning segments (1–8). Many reports claim that these proteins function in active transport—that is, concentration—of the charged species, NH4+ (2, 4, 6, 8). Contrary to previous conclusions, we present evidence here that the AmtB/MEP proteins function in acquisition of the uncharged species, NH3, and that the AmtB protein of Salmonella enterica serovar Typhimurium (S. typhimurium) does not concentrate this species. Furthermore, we find no evidence that S. typhimurium has a mechanism for active transport of NH4+ or NH3.
MATERIALS AND METHODS
Construction of a Single Copy amtB-lacZ Fusion.
Escherichia coli strain NCM1888, which carries an amtB-lacZ fusion in single copy at the trp locus, was constructed according to Elliot (9). A 1.3-kilobase (kb) NdeI–EcoNI fragment from plasmid p149B6 (10), which carries the amtB region of E. coli, was made blunt-ended with the Klenow fragment of DNA polymerase I, and ligated to the 12.5-kb BamHI fragment of pRS551 (11), also blunt-ended, to yield the plasmid pJES1070. After linearization, pJES1070 was transformed into strain TE2680 (recD) to yield strain NCM1888 (kanamycin-resistant). The amtB-lacZ fusion was transferred by phage P1vir-mediated transduction to other E. coli strains and to S. typhimurium strain SL4213 (galE) to yield SK3312. It was transferred to other S. typhimurium strains by P22-mediated transduction.
Construction of an amtB∷Spcr Gene Disruption.
Strain NCM1776 was obtained by insertion of a spectinomycin-resistance cassette (Ω Spc) into the amtB gene of strain NCM1469 (11). A 2-kb SmaI fragment from plasmid pUT-Sm/Spc (12), which carries the spectinomycin-resistance cassette, was ligated into p149B6 (10), which had been cleaved with NdeI and made blunt-ended, to yield the plasmid pJES1008. After linearization with SacI, pJES1008 was transformed into NCM1469 to yield NCM1776 (spectinomycin-resistant). To obtain NCM2019, the lesion was transferred to the wild-type strain used in this study, NCM1458, by P1vir-mediated transduction with selection for resistance to spectinomycin (50 μg/ml). Correct insertion of the cassette was confirmed by Southern blot analysis.
Construction of Ptac-amtB Plasmid.
Plasmid pJES1130 carries the amtB gene under control of the tac promotor of pJES851 (11). A 1.9-kb SphI–BamHI fragment from p149B6 was partially cleaved with HaeII to give a 1.7-kb fragment, made blunt-ended, and ligated into pBluescript KS that had been cleaved with EcoRV, to yield plasmid pJES1126. The 1.7-kb EcoRI–HindIII fragment from pJES1126 then was ligated into a 5.1-kb EcoRI–HindIII fragment of the pJES851 vector to yield the plasmid pJES1130, in which the translational start codon for amtB is positioned 67 bp downstream of the tac promotor.
Media and Growth Conditions.
Growth experiments were performed in appropriately supplemented N−C− minimal medium (13) at 37°C. Cultures used for inoculation usually were grown in N−C− at pH 7 with 0.4% glucose and 5 mM NH4Cl. The minimal medium for S. cerevisiae strains was medium 164 (6) supplemented with vitamins and trace elements and with 100 μg/ml of uracil. The pH was adjusted to 6.1 with KOH. Cells were grown aerobically at 28°C with 3% glucose and NH4Cl at the concentrations indicated.
Methylammonium Uptake Assays.
Assays for uptake of [14C]methylammonium were performed as described (14) at pH 7.0. Cells were grown in N−C− medium with 0.4% glucose and 3 mM glutamine (nitrogen-limiting conditions) or 3 mM glutamine plus 10 mM NH4Cl (nitrogen-excess conditions). Cells were harvested at OD600 of 0.5 and washed twice with the assay buffer (50 mM Hepes, pH 7/72 mM NaCl/0.2% glucose) and were concentrated 2-fold in this buffer. To perform an assay, cells were preincubated for 15 min at 37°C before addition of 6 μM [14C]methylammonium (specific activity 6 Ci/mol). Samples (0.5 ml) were removed at different times (1–20 min), rapidly filtered (Millipore filters, type HA, pore size 0.45 μm), and washed twice with cold assay buffer. Methylammonium uptake activity (in pmol/ml per OD600) was calculated as (cpm)/(specific activity × v × OD600), where v is the sample volume and specific activity was 15 cpm/pmol. The rates reported are slopes of plots of methylammonium uptake versus time (minutes). Analysis of 14C-labeled product in cells was performed as described (15). Synthesis of γ-N-methylglutamine was carried out in vitro as follows: E. coli glutamine synthetase (Sigma G-1270) was incubated for 2 hr at 37°C with 1 mM [14C]methylammonium (6 Ci/mol), 20 mM l-glutamate, 4 mM ATP, 40 mM imidazole⋅HCl (pH 7.2), and 3 mM MnCl2.
β-Galactosidase Assays.
Expression of the amtB-lacZ and glnA-lacZ fusions was assessed as described (11). The differential rates of β-galactosidase synthesis reported (in units/ml per OD600) were obtained by determining the slopes of plots of β-galactosidase activity versus the optical density of the culture.
RESULTS
Growth Defects of an amtB Strain of E. coli.
Disruption of amtB and its homologues in bacteria apparently caused no growth defects (1, 7, 8), although disruptions of MEP genes in S. cerevisiae have been shown to impair growth at low concentrations of ammonium (6, 16) (we use “ammonium” to designate both the charged and uncharged species). Given that many fungi grow best at low pH (17) and that studies with the S. cerevisiae strains were performed at pH 6.1, we sought—and found—growth defects of an E. coli strain carrying an amtB disruption at pH values below 7. We disrupted the amtB gene of E. coli by insertion of a spectinomycin-resistance cassette (see Materials and Methods). A search of the database for the E. coli genome and Southern blotting experiments performed at low stringency indicated that there is only a single copy of an amtB gene (data not shown). At pH 7 the amtB mutant strain, NCM2019, had the same doubling time (Table 1) and cell yield (not shown) in batch culture as its congenic parent strain, NCM1458, at initial concentrations of ammonium between 5 and 0.2 mM. However, at pH 5 the mutant strain had a longer doubling time than its parent at concentrations of ammonium ≤ 1 mM. This growth defect was alleviated at 5 mM ammonium, indicating that it was caused specifically by ammonium limitation and not by other effects of low pH. When the amtB disruption was complemented by a plasmid-borne copy of amtB transcribed from the tac promoter, growth defects at low ammonium also were alleviated (Table 1).
Table 1.
Effect of pH on the growth of an amtB strain of E. coli at different ammonium concentrations
| Strain | [NH4+],* mM | pH 7
|
pH
5
|
|||||
|---|---|---|---|---|---|---|---|---|
| 5 | 1 | 0.5 | 0.2 | 5 | 1 | 0.5 | ||
| NCM1458 (11) | 59 | 61 | 65 | 69 | 67 | 78 | 79† | |
| (wild type) | ||||||||
| NCM2019 | 59 | 63 | 64 | 65 | 64 | 173 | 374† | |
| (amtB) | ||||||||
Data are doubling time (min). Cells were grown in N−C− medium, with glucose as carbon source and ammonium as sole nitrogen source. The glucose concentration was 0.4% at 5 mM ammonium and 0.2% at lower ammonium concentrations.
[NH4+ + NH3].
Doubling times of the corresponding strains carrying the Ptac-amtB plasmid were 85 min (NCM3014) and 83 min (NCM3015).
Alleviation of Growth Defects of a mep1 mep2 mep3 Triple-Mutant Strain of S. cerevisiae at pH 7.1.
To date, three homologues of amtB, called MEP1, MEP2, and MEP3, have been identified in the yeast S. cerevisiae (6). As reported, a triple-mutant strain of S. cerevisiae with deletions in all three MEP genes has growth defects at pH 6.1 at concentrations of ammonium ≤ 5 mM (Table 2); these growth defects are largely alleviated by increasing the concentration of ammonium to 20 mM. By analogy to the case for the amtB mutant strain of E. coli, growth defects of the triple-mutant strain at low concentrations of ammonium were relieved noticeably by increasing the pH to 7.1. As was the case for the parental strain of E. coli, neither the ammonium concentration nor the pH had a large effect on the doubling time of the parental strain of S. cerevisiae.
Table 2.
Effect of pH on the growth of an S. cerevisiae mep1mep2mep3 triple-mutant strain at different ammonium concentrations
| Strain | [NH4+],* mM | pH 6.1
|
pH
7.1
|
||||
|---|---|---|---|---|---|---|---|
| 20 | 5 | 1 | 20 | 5 | 1 | ||
| 23344c (6) | 144 | 144 | 146 | 140 | 142 | 140 | |
| (wild type) | |||||||
| 31019b (6) | 166 | 283 | NG† | 158 | 167 | 288 | |
| (mep1Δmep2Δmep3Δ) | |||||||
Data are doubling time (min). Cells were grown in medium 164 with glucose (3%) as carbon source and ammonium as sole nitrogen source.
[NH4+ + NH3].
No growth.
Effect of Constitutively Expressed amtB on Growth of a gltB Strain of S. typhimurium.
To determine whether S. typhimurium is capable of concentrative uptake of ammonium we made use of a mutant strain, gltB, that lacks the activity of the enzyme glutamate synthase (GOGAT) and hence of the high-affinity pathway for assimilation of ammonium into glutamate (the GS-GOGAT cycle). This strain depends on the biosynthetic glutamate dehydrogenase (GDH) to synthesize glutamate, and because GDH has a relatively high Km for ammonium the strain grows slowly at low external ammonium concentrations ≤ 2 mM even at pH 7 (13, 18) (strain SK3062, Table 3). Slow growth appears to be caused by depletion of the internal glutamate pool and concomitant reduction of the steady-state K+ pool (19). As expected if glutamate depletion is caused by low catalytic activity of GDH, the glutamate and K+ pools are restored to normal at a high external ammonium concentration or by a suppressor mutation closely linked to the gdh gene that increases the amount of the GDH protein approximately 10-fold (D.Y. and S.K., unpublished results). Both restore growth to normal (Table 3). It is our working hypothesis that the suppressor mutation, which we call gdhp515, lies in the promoter-regulatory region for gdh and increases gdh expression. The fact that increased amounts of GDH improved growth of the gltB strain indicated that it probably was not capable of concentrating ammonium. However, as discussed below, it does not express amtB.
Table 3.
Effect of the constitutive expression of amtB on growth of an S. typhimurium gltB mutant strain at different pH values and ammonium concentrations
| Strain | [NH4+],* mM | pH 7
|
pH
6
|
||||
|---|---|---|---|---|---|---|---|
| 5 | 1 | 0.5 | 5 | 1 | 0.5 | ||
| SK2633 | 57 | 58 | 63 | 60 | 65 | 66 | |
| (wild type) | |||||||
| SK3390 | 62 | 63 | 62 | 65 | 68 | 64 | |
| (wild type/Ptac-amtB) | |||||||
| SK3062 | 65 | 105 | 160 | 111 | 700 | ≈1,130 | |
| (gltB†) | |||||||
| SK3391 | 68 | 106 | 160 | 105 | 360 | 450 | |
| (gltB/Ptac-amtB) | |||||||
| SK3109 | 60 | 63 | 68 | 66 | 115 | 142 | |
| (gltB gdhp515) | |||||||
| SK3392 | 64 | 68 | 69 | 70 | 88 | 108 | |
| (gltB gdhp515/Ptac-amtB) | |||||||
Data are doubling time (min). Cells were grown in N−C− medium with glucose as carbon source and ammonium as sole nitrogen source. The glucose concentration was 0.4% at 5 mM ammonium and 0.2% at lower ammonium concentrations. Isopropyl β-d-thiogalactoside was present at 1 mM.
[NH4+ + NH3].
The lesion is ΔgltB824 (31).
In E. coli expression of the glnK-amtB operon occurs only under nitrogen-limiting conditions and depends on the enhancer-binding protein NtrC (5). We have obtained evidence that transcription of the glnK-amtB operon is similarly controlled in S. typhimurium (Table 4). In agreement with the fact that growth in the presence of ammonium and gltB lesions both cause a decrease in transcription of NtrC-controlled genes, we found that amtB was poorly expressed in strain SK3062 (gltB) (data not shown). To test effects of AmtB function on growth of this strain, we introduced a plasmid-borne copy of amtB under control of the tac promoter [SK3391 (gltB/Ptac-amtB)]. This failed to improve growth at low concentrations of ammonium at pH 7 over a range of isopropyl β-d-thiogalactoside concentrations (Table 3 and not shown), commensurate with the view that the AmtB protein does not mediate concentration of ammonium. Control experiments indicated that the constitutively expressed amtB gene had no negative effects on growth of either the parental strain or the gltB gdhp515 double-mutant strain (compare strains SK3390 and SK3392 to strains SK2633 and SK3109, respectively).
Table 4.
Expression of the amtB gene and uptake of [14C]methylammonium (MA) by enteric bacteria
| Strain | Expression
of amtB-lacZ,* units/ml per
OD600†
|
MA uptake, pmol/(ml ×
OD600 ×
min)†
|
||
|---|---|---|---|---|
| N-limiting‡ | N-excess‡ | N-limiting‡ | N-excess‡ | |
| E. coli | ||||
| NCM1458 (wild type) | 5,600 | 31 | 52 | ≤3 |
| NCM2019 (amtB) | 4,300 | 27 | ≤3 | ≤3 |
| S. typhimurium | ||||
| SK2979 (wild type) (18) | 6,450 | 35 | 34 | ≤3 |
| SK3020 (ntrC) (18) | 63 | 37 | ≤3 | ≤3 |
| SK3004 [ntrB(Con)] (18) | 10,100 | 5,000 | 58 | 32 |
| SK3307 (glnA) (24) | 8,100 | 6,000 | ≤3 | ND§ |
| SK3290 [glnA ntrB(Con)] | 10,400 | 7,400 | ≤3 | ND§ |
β-Galactosidase assays were performed with congenic strains carrying the amtB-lacZ fusion, strains NCM2023, NCM2021, SK3362, SK3364, SK3382, SK3381, and SK3379, respectively.
See Materials and Methods.
Cells were grown in N−C− medium with glucose (0.4%) as carbon source and glutamine (3 mM) (N-limiting) or glutamine (3 mM) plus ammonium (10 mM) (N-excess).
Not determined.
To confirm that the constitutively expressed amtB gene was functional in strains carrying the gltB lesion, we looked for its effects at pH 6. At pH 6, the gltB strain (SK3062) grew poorly even at an ammonium concentration of 5 mM and had profound growth defects at lower ammonium concentrations. Moreover, even the gltB gdhp515 double-mutant strain (SK3109) grew poorly at ammonium concentrations ≤ 1 mM. Constitutive expression of amtB partially alleviated the growth defects of both strains (compare strains SK3391 and SK3392 to strains SK3062 and SK3109, respectively), confirming the importance of AmtB function at pH values below 7. Improvement in growth of the double-mutant strain at pH 6 (strain SK3392 vs. SK3109) occurred from initial doubling times comparable to those of the single-mutant strain SK3062 at pH 7 (100–150 min), indicating that positive effects of constitutively expressed amtB can be detected even at relatively rapid doubling times.
Role of AmtB in Transport of [14C]Methylammonium.
Because the radioactive isotope of nitrogen, 13N, has a half-life of ≈10 min and ammonium is rapidly metabolized, the radiolabeled ammonium analogue [14C]methylammonium (CH3NH3+ + CH3NH2) has been used to study ammonium transport (20). To study the role of the AmtB protein of enteric bacteria in transport of methylammonium, which is not used as a nitrogen source, we first wanted to determine the conditions under which the amtB gene was expressed. In accord with results of others, we found that expression of an amtB-lacZ fusion in a merodiploid strain of E. coli or S. typhimurium that also carried an intact amtB gene (NCM2023 or SK3362, respectively) was greatly elevated under N-limiting conditions (Table 4). As discussed above, we demonstrated that this elevation depended on an intact ntrC gene in S. typhimurium. As is true of other NtrC-controlled genes, amtB could be highly expressed in media containing ammonium as N source in a mutant strain of S. typhimurium carrying an NtrBconstitutive protein (SK3382); such a protein increases phosphorylation of NtrC and thereby converts it to an active form even under conditions of excess N availability.
Commensurate with the known role of AmtB and its homologues in acquisition of [14C]methylammonium, accumulation of this compound by washed cells could be demonstrated in amtB+ strains of E. coli and S. typhimurium grown under N-limiting conditions but not N-excess conditions (Table 4). Moreover, transport of [14C]methylammonium did not occur in an ntrC strain of S. typhimurium but did occur in a strain carrying an NtrBconstitutive protein even when it had been grown with ammonium as N source. Disruption of amtB in E. coli essentially prevented accumulation of [14C]methylammonium, as had been shown by others (4, 6–8). Addition of an excess of ammonium (60 μM) during the course of an assay for [14C]methylammonium transport by wild-type E. coli (see Materials and Methods) had an inhibitory effect (data not shown) (20).
Complementation of the amtB disruption in E. coli with an amtB+ gene under control of the tac promoter resulted in detectable restoration of [14C]methylammonium acquisition, but not to a level comparable to that seen in an amtB+ strain grown under N-limiting conditions (data not shown). Thus, although amtB is expressed in the complemented strain independent of N availability or NtrC, functional AmtB product does not appear to be present at high levels.
Concentration of Label from [14C]Methylammonium Requires Metabolism.
It has been reported that [14C]methylammonium is concentrated and hence actively transported in a number of microorganisms and it has been inferred that the same is true for ammonium (2, 4, 6, 8, 20). However, it also has been reported that in some organisms methylammonium is converted to methylglutamine (15, 21–23). If the latter is the case, accumulation cannot be equated with active transport because passage across the cytoplasmic membrane is accompanied by metabolism.
Although radiolabel in [14C]methylammonium was accumulated from an external concentration of 6 μM to an intracellular concentration of ≈ 500 μM by amtB+ strains of both E. coli and S. typhimurium (data not shown), two lines of evidence indicate that [14C]methylammonium itself was not concentrated. First, direct characterization of the radiolabeled product accumulated by E. coli indicated that essentially all of it had been metabolized to a compound with the same mobility as [14C]methylglutamine (Fig. 1), in agreement with results of others (15, 21, 22). Given that conversion of methylammonium to methylglutamine is known to be carried out by glutamine synthetase (23), it would be an energy-requiring process and hence would allow accumulation of radiolabel. Second, although amtB was highly expressed in a mutant strain of S. typhimurium carrying a nonpolar point mutation in glnA (24), which encodes glutamine synthetase, and in a glnA ntrB(Con) double-mutant strain, these strains failed to accumulate radiolabel (Table 4). The amount of radiolabel expected after equilibration of [14C]methylammonium would have been below our limit of detection, and hence we can infer only that transport per se was normal.
Figure 1.

Intracellular accumulation of 14C-labeled product from [14C]methylammonium in E. coli. Wild-type E. coli (strain NCM1458) was grown with glutamine (3 mM) as sole nitrogen source. Components of cell extracts were separated by thin-layer chromatography and subjected to autoradiography (see Materials and Methods). Lane 1, [14C]methylammonium; lane 2, [14C]methylglutamine; lanes 3–7, extracts prepared 1.5, 2.5, 5, 8, and 11 min after addition of 14CH3NH3+ to cells.
Given that accumulation of methylammonium depends on its metabolism to methylglutamine, we showed that failure of the E. coli strain carrying an amtB disruption to accumulate radiolabel could not be attributed to a regulatory effect of the amtB lesion on synthesis of glutamine synthetase: transcription of the glnA gene was normal in this strain (data not shown). Thus, AmtB appears to be the only protein capable of transporting methylammonium, at least at low concentrations.
DISCUSSION
Ammonium (NH4+ + NH3) is the preferred nitrogen source for many microorganisms, i.e., the nitrogen source that allows the fastest growth rate. Based on the growth behavior of both enteric bacteria and S. cerevisiae, we propose that AmtB/MEP proteins and their homologues increase the rate of diffusion of the uncharged species, NH3, across the cytoplasmic membrane. This proposal differs from previous views in two ways—first in maintaining that AmtB/MEP proteins recognize the uncharged rather than the charged species and second in maintaining that they do not concentrate this species. Evidence for the first conclusion derives from effects of pH on growth rate: even at low concentrations of ammonium (≤1 mM), the effects of amtB/mep disruptions are profound in batch culture only at pH values below 7. Because the pKa of NH4+ is 9.25 (25), our data indicate that AmtB function is required for optimal growth of E. coli when the external NH3 concentration drops to ≈50 nM or less. [At 1 mM (NH4+ + NH3), this would be the concentration of the uncharged species at pH 5.] To reach this low concentration of NH3 at pH 7 would require reducing the ammonium concentration to 10 μM, which would not give a detectable cell yield in batch culture. Alternatively, it should be possible to study AmtB function at pH 7 in ammonia-limited continuous culture.
We obtained no evidence that the AmtB product concentrates either NH3 or NH4+ across the cytoplasmic membrane: the dependence of [14C]methylammonium accumulation on its metabolism precludes use of [14C]methylammonium uptake assays to assess active transport and the failure of constitutively expressed AmtB to relieve the growth defects of a gltB strain, which appear to be caused by a requirement for a high internal ammonium concentration, indicates that AmtB does not concentrate ammonium. We note, as others have (26), that accumulation of the charged species, NH4+, could result in dissipation of the proton motive force if the uncharged species, NH3, could diffuse back across the phospholipid bilayer in an unmediated manner. This problem might become profound if metabolic conversion of ammonium to glutamate and glutamine were slowed or interrupted. The role of AmtB appears to be analogous to that of GlpF, the glycerol facilitator of E. coli, and the role of glutamine synthetase appears to be analogous to that of glycerol kinase (27). After ammonia has crossed the cytoplasmic membrane, it, like glycerol, is “trapped” in the cell interior by an energy-requiring metabolic conversion. Interestingly, disruption of the amtB gene actually results in faster growth of E. coli on the poor nitrogen source arginine (E.S. and S.K., unpublished work), leading to the hypothesis that NH3 may diffuse outward through AmtB when its internal concentration exceeds that in the medium.
Because there are no homologues of amtB in E.
coli, we propose that the rate of diffusion of NH3
across the cytoplasmic membrane in the absence of a protein mediator is
sufficient to support optimal growth [doubling time of 60
min/nitrogen content 14% of dry weight (28)] at external
concentrations greater than ≈250 nM, the concentration that
would be present at an ammonium concentration of 5 mM at pH 5 or 50
μM at pH 7. It is noteworthy that partition coefficients for
NH3 between water and organic solvents (k =
0.045 to 0.01) (29) and the permeability coefficients calculated from
them by using the Collander relationship [P =
(0.03 ×
k)/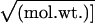 (30), i.e.,
between 3 and 0.7 × 10−4 cm/sec support this view
(see also ref. 26). Interestingly, the sharp drop in the internal
glutamine concentration and the concomitant sharp rise in
glnA-lacZ expression seen in ammonium-limited
continuous cultures of S. typhimurium at pH 7 occurred as
the external ammonium concentration dropped from 80 to <20 μM (18),
the range over which unmediated diffusion of NH3 across the
phospholipid bilayer is predicted to limit growth.
(30), i.e.,
between 3 and 0.7 × 10−4 cm/sec support this view
(see also ref. 26). Interestingly, the sharp drop in the internal
glutamine concentration and the concomitant sharp rise in
glnA-lacZ expression seen in ammonium-limited
continuous cultures of S. typhimurium at pH 7 occurred as
the external ammonium concentration dropped from 80 to <20 μM (18),
the range over which unmediated diffusion of NH3 across the
phospholipid bilayer is predicted to limit growth.
In contrast to our conclusions, other laboratories have proposed that AmtB/MEP proteins and their homologues transport the charged species NH4+ and that transport is active (2, 4, 6). Some experimental bases for the differences in view are explained above. Because the growth defects of mutant strains lacking AmtB/MEP proteins are strikingly similar for enteric bacteria and yeast, we think re-examination of the precise biochemical function of these proteins in other organisms is warranted. Similarly, because unmediated diffusion of NH3 across the cytoplasmic membrane of S. typhimurium appears to be sufficient for rapid growth at external concentrations greater than ≈250 nM, we think the importance of such unmediated diffusion to growth of all microorganisms should be reconsidered.
Acknowledgments
We thank Hiroshi Nikaido for help in obtaining partition coefficients for NH3 and calculating permeability coefficients from them. We thank M. Dean for providing plasmid p149B6, B. André for providing the S. cerevisiae strains, and J. Batut, L. Csonka, G. Fink, B. Magasanik, H.-U. Moesch, H. Nikaido, and N. Pace for valuable criticisms of the manuscript. We are grateful to Norm Pace for his continued interest in this work, which was supported by National Science Foundation Grant MCB-9405733 to S.K.
ABBREVIATIONS
- AmtB
ammonium/methylammonium transport B
- MEP
methylammonium/ammonium permease
- kb
kilobase
- GDH
glutamate dehydrogenase
References
- 1.Wray L V, Atkinson M R, Fisher S H. J Bacteriol. 1994;176:108–114. doi: 10.1128/jb.176.1.108-114.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ninnemann O, Jauniaux J C, Frommer W B. EMBO J. 1994;13:3464–3471. doi: 10.1002/j.1460-2075.1994.tb06652.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Wilson R, Ainscough R, Anderson K, Baynes C, Berks M, Bonfield J, Burton J, Connell M, Copsey T, Cooper J, et al. Nature (London) 1994;368:32–38. doi: 10.1038/368032a0. [DOI] [PubMed] [Google Scholar]
- 4.Siewe R M, Weil B, Burkovski A, Eikmanns B J, Eikmanns M, Kramer R. J Biol Chem. 1996;271:5398–5403. doi: 10.1074/jbc.271.10.5398. [DOI] [PubMed] [Google Scholar]
- 5.vanHeeswijk W C, Radenberg M, Westerhoff H V, Kahn D. Mol Microbiol. 1996;21:133–146. doi: 10.1046/j.1365-2958.1996.6281349.x. [DOI] [PubMed] [Google Scholar]
- 6.Marini A-M, Soussi-Boudekou S, Vissers S, André B. Mol Cell Biol. 1997;17:4282–4293. doi: 10.1128/mcb.17.8.4282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Rudnick P, Colnaghi R, Green A, Kennedy C. In: Biological Nitrogen Fixation for the 21st Century. Elmerich C, Kondorosi A, Newton W E, editors. Vol. 31. Paris: Kluwer; 1997. pp. 123–124. [Google Scholar]
- 8.vanDommelen A, Keijers V, Vanderleyden J, DeZamaroczy M. In: Biological Nitrogen Fixation for the 21st Century. Elmerich C, Kondorosi A, Newton W E, editors. Vol. 31. Paris: Kluwer; 1997. pp. 127–128. [Google Scholar]
- 9.Elliot T. J Bacteriol. 1992;174:245–253. doi: 10.1128/jb.174.1.245-253.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Allikmets R, Gerrard B, Court D, Dean M. Gene. 1993;136:231–236. doi: 10.1016/0378-1119(93)90470-n. [DOI] [PubMed] [Google Scholar]
- 11.He L, Soupene E, Kustu S. J Bacteriol. 1997;179:7446–7455. doi: 10.1128/jb.179.23.7446-7455.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.deLorenzo V, Herrero M, Jakubzik U, Timmis K N. J Bacteriol. 1990;172:6568–6572. doi: 10.1128/jb.172.11.6568-6572.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Csonka L N, Ikeda T P, Fletcher S A, Kustu S. J Bacteriol. 1994;176:6324–6333. doi: 10.1128/jb.176.20.6324-6333.1994. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Servin-Gonzalez L, Bastarrachea F. J Gen Microbiol. 1984;130:3071–3077. doi: 10.1099/00221287-130-12-3071. [DOI] [PubMed] [Google Scholar]
- 15.Rapp B J, Landrum D C, Wall J D. Arch Microbiol. 1986;146:134–141. [Google Scholar]
- 16.Dubois E, Grenson M. Mol Gen Genet. 1979;175:67–76. doi: 10.1007/BF00267857. [DOI] [PubMed] [Google Scholar]
- 17.Griffin D H. Fungal Physiology. New York: Wiley–Liss; 1994. [Google Scholar]
- 18.Ikeda T P, Shauger A E, Kustu S. J Mol Biol. 1996;259:589–607. doi: 10.1006/jmbi.1996.0342. [DOI] [PubMed] [Google Scholar]
- 19.Yan D, Ikeda T P, Shauger A E, Kustu S. Proc Natl Acad Sci USA. 1996;93:6527–6531. doi: 10.1073/pnas.93.13.6527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hackette S L, Skye G E, Burton C, Segel I H. J Biol Chem. 1970;245:4241–4250. [PubMed] [Google Scholar]
- 21.Barnes E M, Zimniak P, Jayakumar A. J Bacteriol. 1983;156:752–757. doi: 10.1128/jb.156.2.752-757.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Jayakumar A, Schulman I, MacNeil D, Barnes E M. J Bacteriol. 1986;166:281–284. doi: 10.1128/jb.166.1.281-284.1986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Servin-Gonzalez L, Ortiz M, Gonzalez A, Bastarrachea F. J Gen Microbiol. 1987;133:1631–1639. doi: 10.1099/00221287-133-6-1631. [DOI] [PubMed] [Google Scholar]
- 24.Krajewska-Grynkiewicz K, Kustu S. Mol Gen Genet. 1983;192:187–197. doi: 10.1007/BF00327665. [DOI] [PubMed] [Google Scholar]
- 25.Hodgman C D, Weast R C, Selby S M. Handbook of Chemistry and Physics. Cleveland, OH: Chemical Rubber Publishing; 1958. [Google Scholar]
- 26.Kleiner D. FEMS Microbiol Rev. 1985;32:87–100. [Google Scholar]
- 27.Richey D P, Lin E C C. J Bacteriol. 1972;112:784–790. doi: 10.1128/jb.112.2.784-790.1972. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Luria S E. The Bacteria. New York: Academic; 1960. [Google Scholar]
- 29.Sandell K. Naturwissenschaften. 1964;51:336. [Google Scholar]
- 30.Collander R. Physiol Plant. 1949;2:300–311. [Google Scholar]
- 31.Madonna M J, Fuchs R L, Brenchley J E. J Bacteriol. 1985;161:353–360. doi: 10.1128/jb.161.1.353-360.1985. [DOI] [PMC free article] [PubMed] [Google Scholar]