
Figure 11. Comparison of human rod responses obtained from paired-flash ERGs and from suction pipette recordings, under dark-adapted and light-adapted conditions.
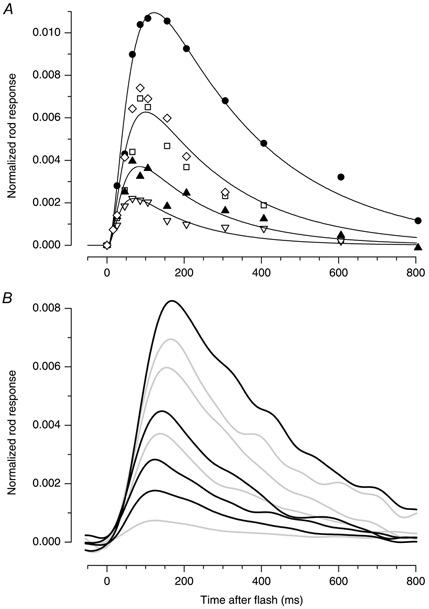
A, rod responses derived with the paired-flash technique for subject TDL. Results from Fig. 10B have been replotted on a slower time base, and for comparison with the suction pipette recordings have been normalized with respect to the dark current, i.e. the ordinate plots the fraction of dark current suppressed per photoisomerization (compare Fig. 9A). On the basis of a troland conversion factor of K= 8.6 photoisomerizations rod−1 Td−1 s−1, the steady backgrounds delivered < 0.1, 46, 180 and 370 R* s−1 (symbols as in Fig. 10B) B, data are from Kraft et al. (1993, Fig. 9), kindly supplied by Dr J. L. Schnapf, and are expressed in the same units as in A. For an assumed effective collecting area of 1.7 μm2, the backgrounds were calculated to deliver 0, 14, 27, 51, 96, 230, 430 and 820 R* s−1. Black traces indicate responses obtained on backgrounds that most closely matched those used in A. Curves in A show a model for rod responses suggested by Nikonov et al. (1998, eqn (19)), comprising the algebraic sum of three exponential decay terms. For the dark-adapted response, the three rate constants were set to 3.5, 25 and 50 s−1. For responses on the three backgrounds, we scaled up each of these rate constants by a fixed factor, of 1.28 (at 46 R* s−1), 1.42 (at 180 R* s−1) and 1.7 (at 370 R* s−1), and we scaled the amplitude of the responses in proportion to the fraction of steady current remaining on the background, i.e. we multiplied by 0.82, 0.68 and 0.56, respectively. In all cases the total delay term was td= 3 ms. The curves should be interpreted with caution, both because we needed to use an unusually high value for the amplification constant, of A= 19 s−2 (cf. ≈4 s−2 for the rising phase of the response, see Fig. 10), and because the formulation ignores calcium feedback mechanisms.