
Figure 6. The inhibition of NMDAR current by AMPARs is independent of voltage, the direction of monovalent cation flow and the availability of extracellular monovalent cations.
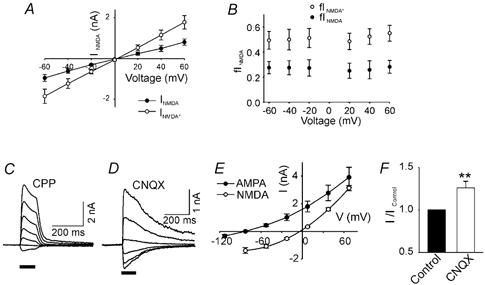
A, NMDAR current in the absence (INMDA, •) and presence of CNQX (10 μM, INMDA*, ○) plotted against voltage clamp holding potential. B, fINMDA* is larger than fINMDA at all holding potentials (-60≈+60 mV, P < 0.01, two-way ANOVA, n = 4). D-AP5 (20 μM) was used to isolate NMDAR current. C and D, current responses to glutamate pulses at different holding potentials (70, 40, 10, −20, −50, −80, −110 mV) in Na+- and K+-free, 10 mm Ca2+ extracellular medium. C, the AMPAR component of the glutamate-induced current was measured after blocking the NMDAR component with 25 μM CPP. D, the NMDAR component of the glutamate-induced response was measured after blocking the AMPAR component with 10 μM CNQX. E, I-V curves for the AMPAR and NMDAR current components under conditions described for C, and D. Note the very different reversal potentials for the two channel subtypes. F, after voltage clamping at the AMPAR channel reversal potential (VH −77 mV), when the average AMPAR channel current is zero, the NMDAR current increased significantly in the presence of 10 μM CNQX (n = 7). A later application of CPP abolished the glutamate-induced current in the presence of CNQX (data not shown).