
FIGURE 8. Mitochondrial redox status and ROS production in response to inhibition of enzymes responsible for mitochondrial GSH regeneration and GSH transport.
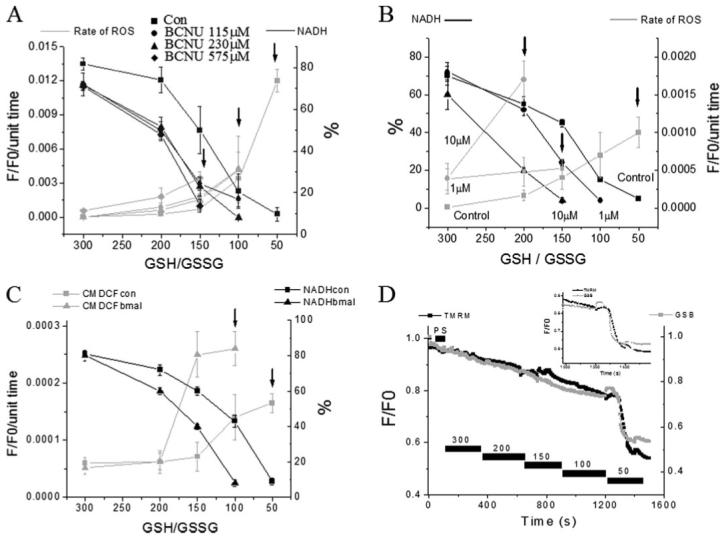
NADH (as % of initial level) and ROS production in saponin-permeabilized cardiomyocytes were exposed to different GSH/GSSG ratios. A, concentration-dependent effects of the glutathione reductase inhibitor BCNU. B, concentration-dependent effects of the NADH/NADPH transhydrogenase inhibitor NBD chloride. C, rate of oxidation of the ROS probe (F/F0/unit time) and NADH (in % of initial fluorescence before permeabilization) in the absence or the presence of 200 μm butyl malonate obtained from four cells (mean ± S.E.; two experiments). The rates of ROS production and NADH oxidation were similar to the control in the presence of 100 μm butyl malonate (not shown). The arrows denote the GSH/GSSG ratio at which ΔΨm collapsed irreversibly. ΔΨm collapses at increasingly higher (more reduced redox potentials) GSH/GSSG ratios in the presence of increasing concentrations of the inhibitors. D, timing of the depletion of reduced glutathione levels and ΔΨm induced by decreasing the GSH/GSSG ratio in saponin-permeabilized cardiomyocytes. Cells were loaded with 100 nm TMRM and 50 μm MCB and then permeabilized to remove the cytoplasmic component of GSB. Inset, note that the rapid oxidation of the mitochondrial GSB signal occurs ∼30 s before the irreversible collapse of ΔΨm. Key to symbols: con, control; bmal, butyl malonate.