

Abstract
Vertebrate limb tendons are derived from connective cells of the lateral plate mesoderm. Some of the developmental steps leading to the formation of vertebrate limb tendons have been previously identified; however, the molecular mechanisms responsible for tendinous patterning and maintenance during embryogenesis are largely unknown. The eyes absent (eya) gene of Drosophila encodes a novel nuclear protein of unknown molecular function. Here we show that Eya1 and Eya2, two mouse homologues of Drosophila eya, are expressed initially during limb development in connective tissue precursor cells. Later in limb development, Eya1 and Eya2 expression is associated with cell condensations that form different sets of limb tendons. Eya1 expression is largely restricted to flexor tendons, while Eya2 is expressed in the extensor tendons and ligaments of the phalangeal elements of the limb. These data suggest that Eya genes participate in the patterning of the distal tendons of the limb. To investigate the molecular functions of the Eya gene products, we have analyzed whether the highly divergent PST (proline-serine-threonine)-rich N-terminal regions of Eya1–3 function as transactivation domains. Our results demonstrate that Eya gene products can act as transcriptional activators, and they support a role for this molecular function in connective tissue patterning.
Keywords: Eyes absent, Splotch, tendon patterning, transcription factor
A major question in development is how precursor cells of various tissue types become organized and arranged appropriately. Limb development has been extensively studied as a model for analyzing mechanisms involved in patterning and morphogenesis. Most studies have focused on the early stages of limb morphogenesis, such as the formation of limb skeletal elements and, more recently, formation of muscle (1–3). Mechanisms responsible for the formation of the limb tendons have received less attention.
The morphological events of tendon formation in the avian limb have been described previously (4, 5). Limb tendons are derived from connective cells of the lateral plate mesoderm. The distal tendons of the foot initially consist of single dorsal and ventral blastemas. As development proceeds, the dorsal blastema differentiates into the extensor tendons, which insert into the base of each phalange, while the ventral blastema undergoes cleavage into the distinct flexor tendons of the digits. As with other components of the limb, both growth and differentiation of the tendons progress in a proximodistal sequence. Previous studies have shown that the distal tendons of the limb can be formed in the absence of muscles (2, 6, 7); however, in the chick, muscles are required for tendon survival—otherwise they undergo degeneration (6, 8).
The growth and patterning of the tendons have been suggested to be independent of skeletal elements (9). For example, although removal of the apical ectodermal ridge (AER) produced truncated digits missing one or two phalanges, the extensor and flexor tendons of the missing phalanges extended distally, joining each other over the regions that corresponded to the normal digits (9). Previous studies have also suggested that the pattern of the musculature appears to be controlled by the connective tissue (10). For example, when pieces of unsegmented thoracic mesoderm are transplanted in place of branchial somites, the transplanted mesoderm can form an appropriate branchial musculature (11), suggesting that the connective tissue in some way governs both tissue and cellular patterning. These facts point to the existence of precise embryonic mechanisms controlling the patterning and maintenance of the limb long tendons.
Hox genes are differentially expressed in mesenchymal cells that give rise to limb skeletal structures, and probably also to connective tissue (12). However, although they are important for specification of skeletal elements (12, 13), Hox genes have not been reported to be expressed during limb tendon formation. Recently, two members of a highly divergent family of homeobox genes, Six1 and Six2 (homologues of the sine oculis gene of Drosophila), have been shown to be expressed in different sets of developing tendons (14), raising the prospect that patterning of connective tissue may be controlled by specific transcription factors. Several other molecules have also been found to be expressed in developing limb tendons, such as the Eph-related receptor tyrosine kinase gene Cek-8 and bone morphogenic protein-7 (Bmp-7) (15, 16). However, their roles in tendon morphogenesis remain unclear.
Three mouse Eya genes, homologues of the eyes absent (eya) gene of Drosophila, have been recently been isolated, and their expression during embryogenesis has been studied (17). Eya1 and Eya2 are widely expressed in cranial sensory placodes and at the sites of inductive tissue interactions during organogenesis, often in complementary or overlapping patterns (17). These features suggest major roles for Eya genes in the development of vertebrate organs and sensory systems. Furthermore, it has been shown that the expression of Eya1 and Eya2 in prospective lens and nasal ectoderm is undetectable in Pax6-deficient Small eye (Sey) mutant embryos, indicating that Eya1 and Eya2 require Pax6 function for their expression in the lens and nasal ectoderm (17). Besides the cranial placodes and developing eye, the Eya genes are widely coexpressed with Pax and Six genes in many tissues during organogenesis, suggesting possible interactions between their gene products and the existence of a conserved Pax–Six–Eya regulatory hierarchy.
In this report, we show that Eya1 and Eya2 transcripts are initially found in connective tissue precursor cells in the limb buds at embryonic day 10.5 (E10.5). Later, Eya1 and Eya2 expression patterns become differentially localized to specific subsets of limb tendons, with Eya1 largely restricted to the flexor tendons (similar to Six2 expression) and Eya2 largely restricted to the extensor tendons (similar to Six1 expression). Our results suggest that the Eya genes function along with the Six genes to establish dorsal/ventral patterning in limb tendon morphogenesis. In addition, we provide insight into the previously unknown molecular function of the Eya gene products by showing that their N-terminal PST domains can activate transcription. We propose that the Eya gene products function as transcription factors involved in specifying connective tissue identity.
MATERIALS AND METHODS
In Situ Hybridization.
Whole-mount and tissue section in situ hybridizations, washing, and RNase treatment were performed as described (17).
Genotypes.
Genotype analysis of Splotch (Sp) embryos at E11.5 was performed as described previously (18).
GAL4/Eya Expression Plasmid.
The GAL4/Eya1–3 expression plasmids were constructed in pBXG1 (19), which expresses the DNA-binding domain of GAL4 (amino acids 1–147) under the control of simian virus 40 enhancer/promoter (ori). Fragments encoding the N-terminal regions of Eya1–3 (amino acids 2–320 for Eya1, amino acids 27–261 for Eya2, and amino acids 2–145 for Eya3) were amplified from mouse Eya1–3 cDNA clones (17). These were cloned as EcoRI fragments into a polylinker immediately downstream of and in frame with the GAL4 DNA-binding domain. All constructs were confirmed by DNA sequencing.
Chloramphenicol Acetyltransferase (CAT) Assay.
P19 cells were plated in 60-mm Petri dishes and transfected with plasmid DNA coated with Transfectam (Promega) according to the manufacturer’s instructions. The pG5ECAT reporter plasmid contains five GAL4 binding sites inserted upstream of the adenovirus E1b minimal promoter driving the CAT gene (20). Each dish was transfected with 2 μg of pG5ECAT DNA and 0.01–1 μg of a GAL4 expression plasmid. The plasmid expressing the GAL4 DNA-binding domain only was used as a negative control. A plasmid expressing the GAL4 DNA-binding domain and activation region II (amino acids 768–881) (21) and a human GAL4-PAX6 PST expression plasmid (19) were used as positive controls. The amount of plasmid transfected into each dish was equalized by the addition of pGEM3Z DNA. Cell extracts were prepared after 48 h and assayed for CAT activity as described (22). CAT assays were performed a minimum of three times, and extracts were diluted as necessary to measure activity in the linear range. The percent acetylation was quantified directly from thin-layer chromatography plates by using a PhosphorImager (Molecular Dynamics) and multiplied by a dilution factor to give relative transcriptional activity.
RESULTS
Eya1 and Eya2 Are Initially Expressed in Myoblast and Connective Tissue Precursors in the Limb Buds.
Both Eya1 and Eya2 are expressed in somites at stage E9.5 (17), and their expression was detected in the dermomyotome at stages E10.5–11.5 (Fig. 1 and data not shown). Myoblast precursor cells for the limb musculature are derived from the lateral portion of the dermomyotome adjacent to the limb bud. These cells migrate into the limb-forming region of the embryo (23–25). Expression of Eya1 in the developing limb was first observed at E10.0 (data not shown), several hours earlier than Eya2 expression. At E10.5, the expression of Eya1 and Eya2 appears to correlate with the two major ventral and dorsal collections of myoblast cells (Fig. 1 A and B), and it progresses distally by E11.5 (Fig. 1 C and D). The ventral expression domain of Eya1 is broader than that of Eya2 at E10.5, and its expression level is higher than that of Eya2 at E11.5 (Fig. 1 A and C). To determine whether the Eya1 and Eya2 limb expression specifically resided in myoblast precursor cells, Eya1 and Eya2 expression was analyzed in Pax3-deficient Splotch mutant embryos (26). In Sp/Sp mutant embryos, myoblast precursor cells do not migrate into the limb and myogenic genes are not expressed there (27, 28). The dorsal and ventral expression domains of Eya1 and Eya2 are not observed in the limbs of Sp/Sp mutant embryos (Fig. 1 E and F). However, in Sp/Sp mutant embryos, some expression of Eya1 is detected in both proximal and distal regions (Fig. 1E, indicated by arrows). In addition, Eya2 expression was observed in discontinuous patches in the limbs of Sp/Sp mutant embryos (Fig. 1F, indicated by arrows). Although our data do not prove that Eya1 and Eya2 are expressed in myogenic precusors, the findings that Eya1 and Eya2 are expressed in somites, lateral dermomyotome, and dorsal and ventral limb mesenchyme and that the latter expression domains are lost in Sp/Sp mutant embryos all support this interpretation. In addition, Eya1 and Eya2 are coexpressed with myogenin later in limb muscle development (data not shown). We thus conclude that a component of Eya1 and Eya2 expression, likely corresponding to myogenic precursor cells, is absent from the limbs of Sp/Sp mutant embryos and that the residual expression of Eya1 and Eya2 corresponds to connective tissue precursors. This view implies that, in the absence of myogenic precursors, connective tissue precursors exhibit anomalous patterns within limb mesenchyme, thus accounting for the dispersed patches of Eya expression.
Figure 1.

Eya1 and Eya2 are expressed in myoblast and connective tissue precursors. (A–F) Transverse sections of 35S-labeled in situ hybridization in wild-type and Sp/Sp mutant limbs. (A–D) The dorsal (d) and ventral (v) Eya1- and Eya2-expressing cell populations. At E11.5, Eya1 expression levels in the ventral domain are higher than that of Eya2 and the expression extends more distally (C). In contrast, Eya2 expression in the distal region is broader than that of Eya2 at E11.5 (D). (E and F) Eya1 and Eya2 expression in the limbs of Sp/Sp mutant embryos. The dorsal and ventral expression domains are lost in the mutant limbs. The residual expression of Eya1 and Eya2 (as indicated by arrows) corresponds to connective tissue precursors. dm, Dermomyotome; so, somites.
Eya1 and Eya2 Are Expressed in Distinct Sets of Tendons During Limb Development.
To study the role of Eya genes in connective tissue patterning during limb development, we further analyzed their expression during tendon morphogenesis. At E12.5, high levels of Eya1 expression are observed ventrally in the foot plate (Fig. 2A). At E13.5, individual digits are visible and cartilage begins to condense. Eya1 expression is found in the developing tendons (Fig. 2F). By E14.5, different digits are well formed and separated. Eya1 expression remains strong in the developing tendons (Fig. 2 B and G). Transverse sections reveal that the expression of Eya1 in condensing mesenchymal cells is associated with the development of flexor tendons (Fig. 2 C–E). (In Fig. 2 C–E, the ectoderm staining is artifactual and arises because the whole mount was intentionally overdeveloped to show mesenchymal Eya1 expression.) Eya1 expression is initially observed in a broad domain (Fig. 2C). As cartilage condensation proceeds, the broad domain of Eya1 expression becomes concentrated and predominantly mesenchymal (Fig. 2 D and E and data not shown). Eventually, Eya1-expressing cells are seen below the cartilage elements, and these tendon-like structures become localized toward the center of the limb (Fig. 2G). Eya1 expression remains strong in the flexor tendons at E17.5, the latest stage examined (Fig. 1H). Besides the developing tendons, Eya1 expression is also observed in the zones of joint formation (Fig. 2I), and in the interdigital regions at E13.5 as well as in the perichondrium at E15.5 (data not shown).
Figure 2.

Eya1 is expressed in tendons of the limb. Whole mount (A–E) and radioactive (F–I) in situ hybridization in the developing limb. (A and B) Ventral views of whole-mount limbs, showing the expression in the foot plates at E12.0–14.0. (C–E) Transverse sections of whole-mount limbs show ventral (v) expression initially in a broad domain and in condensing mesenchyme flanking the cartilage condensations, as indicated by arrows. Arrows in D point to two domains of expression. The ectoderm staining is artifactual and results because the whole-mount in situ hybridization was intentionally overdeveloped to show the mesenchymal Eya1 expression. (F–H) Longitudinal sections show the expression in the developing tendons (t) at E13.5–17.5. (I) Longitudinal section shows the expression of Eya1 in the zone of joint formation (arrows) at E14.5. (F–I) Anterior is to the left and distal is up. Other abbreviations: a, anterior; p, posterior; d, dorsal.
Eya2 expression is strongly detected in the condensing mesenchyme flanking the cartilage of individual digits at E12.0–13.5 (Fig. 3 A, C, and F), and in regions flanking the phalangeal components of the digits at E14.5 (Fig. 3 B, D, E, G, and H). (As in Fig. 2 C–E, the ectoderm staining shown in Fig. 3 C–E is artifactual.) At E14.5, Eya2 expression is also observed dorsally in the condensing mesenchymal cells that will form extensor tendons (Fig. 3 E, arrows, and H). Although, the condensed dorsal extensor blastemas are smaller than ventral flexor blastemas, Eya2 expression remains in these structures at E17.5, the latest stage examined (data not shown). Eya2 transcripts were also found in the perichondrium at E14.5–15.5, but not in the interdigital regions (data not shown). Our data show that the Eya2-expressing cells in the limbs correspond to tendons and probably also to ligaments of the phalangeal elements of the developing digits. Interestingly, Eya2 is expressed in the extensor tendons, whereas Eya1 is expressed in the flexors.
Figure 3.

Eya2 is expressed in tendons and ligaments of the limb. Whole-mount (A–E) and radioactive (F–H) in situ hybridization in the developing limb. (A and B) Dorsal views of whole-mount limbs, showing the expression anteroposteriorly in the mesenchymal cells flanking the cartilage (A, arrows) at E12.0, and flanking the phalangeal components of the digits (B, the regions indicated by the arrow) at E14.0 in the foot plates. (C–E) Transverse sections of whole-mount limbs show the expression of Eya2 anteroposteriorly in the condensing mesenchyme flanking the cartilage condensations at E13.0 (C, arrow), flanking the phalangeal components of the digits at E14.0 (D, arrow, and E) and in the dorsal condensed mesenchyme that will form extensor tendons (E, arrows). The ectoderm staining is artifactual and results because the whole-mount in situ hybridization was intentionally overdeveloped to show the mesenchymal Eya2 expression. (F–H) Longitudinal sections show the expression anteroposteriorly in the condensing mesenchyme flanking the cartilage at E13.5 (F), in the mesenchymal cells flanking the phalangeal components of the digits, and in the developing tendons (t) at E14.5 (G and H). In F–H anterior is to the left and distal is up. Other abbreviations: a, anterior; d, dorsal; p, posterior; v, ventral.
Transcriptional Activation Function of the Eya N-Terminal PST Domain.
The molecular functions of the Eya proteins are currently unknown. The highly divergent Eya N termini consist of 35–40% proline, serine, and threonine residues and resemble the proline–serine–threonine (PST) transactivation domains found in other transcription factors (29, 30). To determine whether the PST domains of Eya1–3 can function as transactivation domains, we constructed a series of expression plasmids in which the N-terminal region of Eya1, -2, or -3 was fused in frame to the 147-amino acid DNA-binding domain of yeast GAL4 (Fig. 4A) and cotransfected them into P19 embryonic carcinoma cells with a reporter plasmid containing the CAT gene (see Materials and Methods). The GAL4-Eya1 and -Eya2 fusion proteins strongly stimulated CAT expression over a wide range of plasmid concentrations (Fig. 4B), at a level approximately one-fifth that of the GAL4(II) positive control and similar to the level of the GAL4-PAX6 PST positive control (Fig. 4C). In contrast, the Eya3 PST domain expression construct stimulated CAT expression relatively weakly, with 8-fold less activity compared with Eya1 and Eya2 (Fig. 4 B and C). Similar data were obtained from three independent experiments. We conclude that the Eya proteins are transcriptional activators and their activation function resides within the N-terminal PST domains.
Figure 4.

Transcriptional activation by the N-terminal PST domains of Eya1–3 proteins. (A) Schematic diagram of GAL4 expression constructs. The GAL4 DNA-binding domain (BD; amino acids 1–147) was used as a negative control. The GAL4 activation domain II (AD; amino acids 768–881) and PAX6 PST domains were used as positive controls. N-terminal regions of Eya1 (amino acids 2–320), Eya2 (amino acids 27–261), and Eya3 (amino acids 2–145) are fused to the GAL4 DNA-binding domain. (B) CAT assay. P19 cells were transfected with the pG5ECAT reporter and 0.01–1 μg of a plasmid expressing the GAL4 DNA-binding domain only or with the GAL4 activation domain, PAX6 PST domain, or the Eya1 PST domain fused to the GAL4 DNA-binding domain. (C) Relative activity of GAL4 expression constructs (symbols are defined in A). The PST domain of Eya1 stimulates transcription approximately  as well as the GAL4 activation domain and approximately
as well as the GAL4 activation domain and approximately  as well as the PAX6 PST domain. The PST domain of Eya2 stimulates transcription approximately
as well as the PAX6 PST domain. The PST domain of Eya2 stimulates transcription approximately 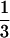 as well as the GAL4 activation domain, while Eya3 only weakly stimulates transcription and shows 8-fold less activity compared with either Eya1 or Eya2, although 6-fold higher than the negative control (see Inset).
as well as the GAL4 activation domain, while Eya3 only weakly stimulates transcription and shows 8-fold less activity compared with either Eya1 or Eya2, although 6-fold higher than the negative control (see Inset).
DISCUSSION
Eya Genes and Connective Tissue Patterning.
In Splotch mutant embryos, the Eya1 and Eya2 expression patterns in the limb buds appear to include both myoblast precursors and mesenchymal cells. Eya1 expression in the developing limb was detected several hours earlier than Eya2 expression (data not shown). At E10.5–11.5, both genes are expressed in the dorsal and ventral limb, where the expression level of Eya1 is higher than that of Eya2 (Fig. 1). In the developing tendons, Eya1 is expressed ventrally in mesenchymal cells (the flexor blastemas) flanking the cartilage condensations (Fig. 2). In contrast, the cells expressing Eya2 are found anteroposteriorly in the regions flanking the cartilage condensations, and later on in the regions flanking the phalangeal components of the digits and in the developing extensor tendons (Fig. 3). The cells expressing Eya2 probably also correspond to the ligaments of the phalangeal elements of the developing digits. The Eya1 and Eya2 expression patterns in the dorsal and ventral limb, and their proximodistal extension, coincide with tendon blastema formation (9). The distal limb tendons initially consist of a single dorsal and ventral blastema, which upon further subdivision will give rise to the different tendons that insert in the digit phalanges. As with other limb components, the growth and differentiation of tendons occurs in a proximodistal direction. The expression of Eya1 specifically in the flexor blastemas and Eya2 in the extensor blastemas indicates that these genes may pattern the flexors and extensors of the phalangeal elements. In addition, Eya1 and Eya2 may specify cell type precursors involved in connective tissue development. Interestingly, haploinsufficiency for human EYA1 has recently been found responsible for the branchio-oto-renal (BOR) syndrome (31), a dysmorphosis not known to involve connective tissue. However, this result does not discount a role for Eya1 in connective tissue patterning, because such function might be observed only in the context of complete loss of gene function.
Recently, expression of several other genes has been identified in the developing long tendons of the autopod (14–16). The expression of Eya1 in the developing limb tendons shows a pattern of expression similar to that of Six2, with expression initially confined to a broad sheet under the ectoderm that rapidly condenses into a mesenchymal layer. However, one difference is that Eya1 is expressed only ventrally in the condensing mesenchymal cells, whereas Six2 is expressed both dorsally and ventrally in the condensing mesenchymal cells, although stronger ventrally (14). Eya1 is expressed in limb buds one day earlier than Six2 (14). Eya2 shows an expression pattern similar to that of Six1 in the developed phalanges at E14.5 (14). It will be interesting to assess the relation between Eya gene expression and Six gene expression, because the same pair of genes are also widely coexpressed in many other tissues during organogenesis, with Eya1 and Six2 in the craniofacial tissues, nasal placode, gut mesenchyme, nephrogenic cord, genital tubercle, and kidney, Eya2 and Six1 in the craniofacial tissues, nasal placode, branchial arch, tongue, kidney, somite, dorsal root ganglion, and future intercostal muscles. These data suggest the possibility of direct molecular interactions between the Eya and Six gene products during organogenesis. The Eya genes together with Six genes provide a key opportunity to explore the link between patterning mechanisms and tissue formation in developing limbs.
Both Eya1 and Eya2 are expressed not only in developing connective tissue but also in somites. Thus, Eya1 and Eya2 may be expressed at the same time both in myoblast precursors migrating from the somite into the limb and in a subpopulation of lateral plate-derived mesenchymal cells that will give rise to the connective tissue. This may ensure that the tendon and muscle precursor cells expressing the same set of genes migrate to appropriate places. Besides in the developing limb, Eya1 and Eya2 are also expressed in other body skeletal muscles and connective tissues (17). In conclusion, we propose that Eya1 and Eya2 are involved in patterning limb connective tissue (tendon and ligaments), and these genes may also subserve this function in other body connective tissues and in skeletal muscles in which they are expressed.
The Molecular Functions of Eya Gene Products.
On the basis of their amino acid content, the N-terminal regions of Eya1–3 were tested for transcriptional activity. Consistent with the observation that Drosophila Eya is localized in the cell nucleus (32), our data indicate that the N-terminal PST domains of Eya1–3 are transcriptional activators in cultured cells (Fig. 4). Interestingly, the PST domains of Eya1 and Eya2 show strong transcriptional activity, while that of Eya3 is comparatively weak (Fig. 4). Eya3 is the most divergent of the three mouse Eya family members, and its expression is different from that of Eya1 and Eya2 during mouse embryonic development (17). Eya1 and Eya2 are expressed in all cranial placodes and their derivatives at E9.5, whereas Eya3 is expressed in the underlying craniofacial and branchial arch mesenchyme at the same stage (17). Taken together, these data suggest that Eya3 may have distinct developmental roles during mouse embryonic development.
The Drosophila Eya N-terminal region has not been tested for transcriptional activation function, but it is also PST-rich and contains a large number of alanine, glycine, and glutamine residues, suggesting that despite their sequence divergence, the Drosophila Eya and murine Eya gene products could have a conserved molecular function during evolution. Although the Eya proteins do not possess a known DNA-binding motif, analysis of the Eya protein sequences suggests that the highly conserved Eya domain could interact either with DNA or a DNA-binding protein to activate transcription, since the Eya domain is predicted to be hydrophobic and to contain two conserved α-helical regions. In conclusion, the identification of transcriptional activation functions for the Eya proteins provides insight into the functional role of these gene products during mouse embryonic development.
Acknowledgments
This work was supported by National Eye Institute Grant 1RO1 EY10123 and by the Howard Hughes Medical Institute.
ABBREVIATIONS
- En
embryonic day n
- CAT
chloramphenicol acetyltransferase
References
- 1.Saunders J W J. In: Vertebrate Limb and Somite Morphogenesis. Ede D A, Hinchliffe J R, Balls M, editors. Cambridge, U.K.: Cambridge Univ. Press; 1977. pp. 1–24. [Google Scholar]
- 2.Shellswell G B, Wolpert L. In: Vertebrate Limb and Somite Morphogenesis. Ede D A, Hinchliffe J R, Balls M, editors. Cambridge, U.K.: Cambridge Univ. Press; 1977. pp. 71–86. [Google Scholar]
- 3.Robson L G, Kara T, Crawley A, Tickle C. Development. 1994;120:1265–1276. doi: 10.1242/dev.120.5.1265. [DOI] [PubMed] [Google Scholar]
- 4.Wortham R A. J Morphol. 1948;83:105–148. doi: 10.1002/jmor.1050830106. [DOI] [PubMed] [Google Scholar]
- 5.Sullivan G E. Aust J Zool. 1962;10:458–516. [Google Scholar]
- 6.Kieny M, Chevallier A. J Embryol Exp Morphol. 1979;49:153–165. [PubMed] [Google Scholar]
- 7.Holder N. Development. 1989;105:585–593. doi: 10.1242/dev.105.3.585. [DOI] [PubMed] [Google Scholar]
- 8.Brand B, Christ B, Jacob H J. Am J Anat. 1985;173:321–340. doi: 10.1002/aja.1001730408. [DOI] [PubMed] [Google Scholar]
- 9.Hurle J M, Ros M A, Ganan Y, Macias D, Critchlow M, Hinchliffe J R. Cell Differ Dev. 1990;30:97–108. doi: 10.1016/0922-3371(90)90078-b. [DOI] [PubMed] [Google Scholar]
- 10.Chevallier A, Kieny M. Roux’s Arch Dev Biol. 1982;191:277–280. doi: 10.1007/BF00848416. [DOI] [PubMed] [Google Scholar]
- 11.Butler J, Cosmos E, Cauwenbergs P C. Development. 1988;102:763–772. [Google Scholar]
- 12.Oliver G, Sidell N, Fiske W, Heinzmann C, Mohandas T, Sparkes R, De Robertis E M. Genes Dev. 1989;3:641–650. doi: 10.1101/gad.3.5.641. [DOI] [PubMed] [Google Scholar]
- 13.Morgan B A, Tabin C J. Curr Opin Genet Dev. 1993;3:668–674. doi: 10.1016/0959-437x(93)90105-x. [DOI] [PubMed] [Google Scholar]
- 14.Oliver G, Wehr R, Jenkins N, Copeland N G, Cheyette B N R, Hartenstein V, Zipursky L, Gruss P. Development. 1995;121:693–705. doi: 10.1242/dev.121.3.693. [DOI] [PubMed] [Google Scholar]
- 15.Macias D, Ganan Y, Sampath T K, Piedra M E, Ros M A, Hurle J M. Development. 1997;124:1109–1117. doi: 10.1242/dev.124.6.1109. [DOI] [PubMed] [Google Scholar]
- 16.Patel K, Nittenberg R, D’Souza D, Irving C, Burt D, Wilknson D G, Tickle C. Development. 1996;122:1147–1155. doi: 10.1242/dev.122.4.1147. [DOI] [PubMed] [Google Scholar]
- 17.Xu P-X, Woo I, Her H, Beier D R, Maas R L. Development. 1997;124:219–231. doi: 10.1242/dev.124.1.219. [DOI] [PubMed] [Google Scholar]
- 18.Epstein J A, Shapiro D N, Cheng J, Lam P Y P, Maas R L. Proc Natl Acad Sci USA. 1996;93:4213–4218. doi: 10.1073/pnas.93.9.4213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lillie J W, Green M R. Nature (London) 1989;338:39–44. doi: 10.1038/338039a0. [DOI] [PubMed] [Google Scholar]
- 20.Ma J, Ptashne M. Cell. 1987;48:847–853. doi: 10.1016/0092-8674(87)90081-x. [DOI] [PubMed] [Google Scholar]
- 21.Glaser T, Jepeal L, Edwards J G, Young S R, Favor J, Maas R L. Nat Genet. 1994;7:463–470. doi: 10.1038/ng0894-463. [DOI] [PubMed] [Google Scholar]
- 22.Gorman C M, Moffat L F, Howard B H. Mol Cell Biol. 1982;2:1044–1051. doi: 10.1128/mcb.2.9.1044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Chevallier A, Kieny M, Mauger A. J Embryol Exp Morphol. 1977;41:245–258. [PubMed] [Google Scholar]
- 24.Jacob M, Christ B, Jacob H J. Anat Embryol. 1978;153:179–193. doi: 10.1007/BF00343373. [DOI] [PubMed] [Google Scholar]
- 25.Soulursh M, Drake C, Meier S. Dev Biol. 1987;121:389–396. doi: 10.1016/0012-1606(87)90175-8. [DOI] [PubMed] [Google Scholar]
- 26.Epstein D J, Vekemans M, Gros P. Cell. 1991;67:767–774. doi: 10.1016/0092-8674(91)90071-6. [DOI] [PubMed] [Google Scholar]
- 27.Bober E, Franz T, Arnold H H, Gruss P, Tremblay P. Development. 1994;120:603–612. doi: 10.1242/dev.120.3.603. [DOI] [PubMed] [Google Scholar]
- 28.Goulding M D, Lumsden A, Paquette A J. Development. 1994;120:957–971. doi: 10.1242/dev.120.4.957. [DOI] [PubMed] [Google Scholar]
- 29.Mermod N, O’Neill E A, Kelly T J, Tjian R. Cell. 1989;58:741–753. doi: 10.1016/0092-8674(89)90108-6. [DOI] [PubMed] [Google Scholar]
- 30.Theill L E, Castrillo J-L, Wu D, Karin M. Nature (London) 1989;342:945–948. doi: 10.1038/342945a0. [DOI] [PubMed] [Google Scholar]
- 31.Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, Well D, Cruaud C, Sahly I, Leibovici M, Bitner-Glindzicz M, Francis M, Lacombe D, Vigneron J, Charachon R, Boven K, Bedbeder P, van Regemorter N, Weissenbach J, Petit C. Nat Genet. 1997;15:157–164. doi: 10.1038/ng0297-157. [DOI] [PubMed] [Google Scholar]
- 32.Bonini N M, Leiserson W M, Benzer S. Cell. 1993;72:379–395. doi: 10.1016/0092-8674(93)90115-7. [DOI] [PubMed] [Google Scholar]