

Abstract
Alcaligenes eutrophus genes encoding the enzymes, β-ketothiolase (phaA), acetoacetyl-CoA reductase (phaB), and polyhydroxyalkanoate synthase (phaC) catalyze the production of aliphatic polyester poly-d-(−)-3-hydroxybutyrate (PHB) from acetyl-CoA. PHB is a thermoplastic polymer that may modify fiber properties when synthesized in cotton. Endogenous β-ketothiolase activity is present in cotton fibers. Hence cotton was transformed with engineered phaB and phaC genes by particle bombardment, and transgenic plants were selected based on marker gene, β-glucuronidase (GUS), expression. Fibers of 10 transgenic plants expressed phaB gene, while eight plants expressed both phaB and phaC genes. Electron microscopy examination of fibers expressing both genes indicated the presence of electron-lucent granules in the cytoplasm. High pressure liquid chromatography, gas chromatography, and mass spectrometry evidence suggested that the new polymer produced in transgenic fibers is PHB. Sixty-six percent of the PHB in fibers is in the molecular mass range of 0.6 × 106 to 1.8 × 106 Da. The presence of PHB granules in transgenic fibers resulted in measurable changes of thermal properties. The fibers exhibited better insulating characteristics. The rate of heat uptake and cooling was slower in transgenic fibers, resulting in higher heat capacity. These data show that metabolic pathway engineering in cotton may enhance fiber properties by incorporating new traits from other genetic sources. This is an important step toward producing new generation fibers for the textile industry.
Keywords: bioplastics, promoters, transgenic cotton, thermal properties, particle bombardment
Cotton is the premier natural fiber for textile applications. It is a biological composite of cellulose, small quantities of hemicellulose, pectins, and proteins that provides excellent wearability and aesthetics. However, further improvements in strength, length, chemical reactivities for dye binding, water absorption, thermal properties, and wrinkle and shrinkage resistance are desirable for textile and other industrial applications. Over the last several decades, significant improvements have been made in the physical properties of cotton through classical plant breeding (1). Nevertheless, the potential for further fiber property improvements through breeding is limited due to requirements for species compatibility and the traits available. In this context, we are applying recombinant DNA technology to transfer genetic traits from diverse sources into cotton (2).
Cotton fiber or seed hair is a terminally differentiated single epidermal cell made up of two walls, primary and secondary. During the initial stages of fiber development (initiation and primary wall formation), the cell elongates up to 3 cm over a period of 20 days post anthesis (DPA). The primary wall is estimated to be 100 to 200 molecules in thickness (0.1 to 0.2 μm), and is made up of cellulose (30%), other neutral acid polysaccharides, waxes, pectic compounds, and proteins (3, 4). The secondary wall (8 to 10 μm) is made up of cellulose that is deposited during the third developmental stage. The secondary wall formation occurs during 16 to 45 DPA. Maturation, the final stage of fiber development (45 to 50 DPA), is associated with changes in mineral content and protein levels. At maturity, cotton fiber is 89% cellulose (for reviews, see refs. 3 and 5). The chemical composition and microstructure of primary and secondary walls influence properties, such as chemical reactivity, thermal characteristics, water absorption, and strength of fiber (3). These properties are important for the manufacturing of textile products and must be preserved in genetically engineered cotton fiber. Hence one potential approach is to synthesize a second biopolymer within the fiber lumen without affecting fiber wall integrity. The new biopolymer is sheltered inside the cellulose walls and does not come in contact with skin. Here we consider a natural thermoplastic polyester compound, poly-d-(−)-3-hydroxybutyrate (PHB) for synthesis in fiber.
PHB is an archetype that is a natural biodegradable thermoplastic with similar chemical and physical properties as polypropylene (6). They are produced by many genera of bacteria as inclusion bodies to serve as carbon sources and electron sink (7). The formation of PHB in bacteria involves three enzymes: β-ketothiolase, NADPH-dependent acetoacetyl-CoA reductase, and PHA synthase (for reviews, see refs. 7 and 8). Two molecules of acetyl-CoA are joined by β-ketothiolase to form acetoacetyl-CoA. Acetoacetyl-CoA is reduced by acetoacetyl-CoA reductase to R-(−)-3-hydroxybutyryl-CoA. This activated monomer is then polymerized by PHA synthase to form PHB. It has been shown that PHB can be synthesized in transgenic plants (9). Our objective is to produce PHB in cotton fiber lumen to modify the chemical and thermal properties of fiber.
MATERIALS AND METHODS
Materials.
Cotton plants (Gossypium hirsutum L. cv DP50) were grown in the greenhouse. The protocols for RNA, DNA, and protein isolations from fibers were as described (10, 11). Protocols for Northern and Southern blot analyses, plasmid subcloning, and other standard molecular biology techniques have been described by Ausubel et al. (12). PCR was used to amplify the coding region of the phaB gene (12). The reaction mix included two primers, ATTAAGGATCCATGACTCAGCGCATTGCG and GGATTAGGATCCGCAGGTCAGCCCATATGC, and the genomic DNA of Alcaligenes eutrophus. The primers were based on the sequence of the gene and contained convenient cloning sites (13). The amplified DNA after digestion with BamHI and XhoI was cloned into pBCSK+ (Stratagene) and was sequenced to confirm the correct primary structure (Lofstrand Laboratories, Gaithersburg, MD). The phaB gene was linked to a poly(A) addition signal (280 bp) of Agrobacterium nopaline synthase gene at the 3′ end and was ligated to a cotton promoter E6 (10). A cauliflower mosaic virus 35s promoter-linked β-glucuronidase (GUS) gene was added to the above plasmid to generate cotton expression vector, pE6-B. A similar construct with a second cotton promoter, FbL2A was generated (pFbL2A-B). The E6 and FbL2A promoter fragments contained 33 and 44 bp of untranslated 5′ leader sequences, respectively, and translation initiation is expected to be at the first ATG codon of phaB gene. The phaC gene coding region (1770 bp; ref. 14) was amplified from A. eutrophus DNA as two fragments and ligated together to form the complete gene. Primers AACATGAATTCATGGCGACCGGCAAAGG and AATTAGGATCCGCGAGATCTTGCCGCGTG were used to amplify a 580-bp 5′ fragment of the gene containing a unique BglII site. The 3′ end of the gene (1200 bp) was then amplified using primers CACGCGGCAAGATCTCGC and TGTAAGGATCCTCATGCCTTGGCTTTGACG. The 580-bp fragment was digested with EcoRI and BamHI and cloned into pBCSK+ vector. The pBCSK+ plasmid containing the 580-bp 5′ fragment and the 1200-bp PCR product were digested with BglII and BamHI and ligated together. The sequence of the insert was determined. The phaC insert was also cloned into the expression vector containing 35s promoter and Nos poly(A) addition signal to generate p35-C plasmid. Enzymatic assays for thiolase were done according to Nishimura et al. (15). Acetoacetyl-CoA reductase activity was measured by a NADPH-dependent spectrophotometric assay as described by Saito et al. (16), and that of PHA synthase by a radiometric assay with [3H]-dl-(−)-3-hydroxybutyryl-CoA (17).
Transformation of Cotton.
Chimeric genes, pE6-B (or pFbL2A-B), and p35-C were introduced into cotton by particle bombardment as described in detail by McCabe and Martinell (18). In brief, surface-sterilized DP50 seeds were germinated and embryonic axes were removed. The meristem of each seed axis was exposed by dissection under a microscope and incubated in Murashige and Skoog salt mixture medium overnight at 15°C in the dark. The meristems were subjected to particle bombardment using an Agracetus Accell electrical discharge device described by Christou et al. (19). Plasmids were precipitated onto 1.0 μm gold beads (Degussa, South Plainfield, NJ) loaded onto 18 × 18 mm squares of aluminized Mylar (DuPont 50MMC) carrier sheets at a rate of 0.05 mg gold + DNA complex per cm2. The carrier sheets were then accelerated toward the cotton seed axes by discharge of an 18 kV arc (18). The seed axes were then allowed to germinate and grow for 4 to 6 weeks before testing leaves for expression of GUS (20). The resulting plants are chimeric for the input genes. Each leaf is tested for GUS activity and selectively pruned to isolate nodes or axillary buds subtending the transformed leaves. The process is continued until a transformed plant is obtained. The transformation process resulted in either epidermal or germ-line transformants. In the former, only the epidermis was transformed and the transgene was not passed on into its seeds. The germ-line transformants, on the other hand, passed the transgene into their progeny in a Mendelian fashion (18). Cotton fibers are epidermal cells and, therefore, both epidermal or germ-line transformants are useful in evaluating fiber modifications.
Detection of PHB and Measurement of Thermal Properties.
Transgenic fibers were treated with Nile blue A fluorescent stain and examined at excitation wavelengths of 546 nm (21). PHB granules were also detected by transmission electron microscopy analysis (University of Wisconsin, Madison) (22). PHB was extracted and converted to crotonic acid and detected by HPLC (23). Gas chromatography (GC)–mass spectrometry (MS) analysis of the ethyl ester derivative of PHB was done as described by Findlay and White (24) at the University of Wisconsin (Madison) Chemistry Department.
Thermal conductivity (TC), thermogravimetric analysis, specific heat, and differential scanning calorimetry (DSC) measurements of cotton fibers were undertaken by MATECH Associates (Scranton, PA) using TA instruments (New Castle, DE). Thermal analysis protocols conformed with those recommended by International Confederation for Thermal Analysis (25).
RESULTS
Expression of PHB Biosynthetic Pathway in Transgenic Fibers.
Plasmids containing chimeric genes, acetoacetyl-CoA reductase (pE6-B or pFbL2A-B), and PHA synthase (p35-C), were introduced into 14,000 cotton seed axes by particle bombardment. A total of 30 transgenic plants (21 epidermals and 9 germ lines) were selected based on GUS expression. The transformation frequency (0.21%) was similar to previous experiments (26). Several of the transformants were tested for pE6-B or p35-C by Southern blot analysis and examples are shown in Fig. 1 A and B. Both epidermal and germ-line transformants showed the presence of phaB and phaC genes, whereas DP50 genomic DNA showed no hybridization. The expression of phaB and phaC genes in one of the transformants, no. 7148, was tested by Northern blot analysis (Fig. 1 C and D). The fibers showed 1.0-kb phaB and 1.9-kb phaC transcripts, respectively (Fig. 1 C and D). The above results indicate that both epidermal and germ-line transformants contain transcriptionally active phaB and phaC genes.
Figure 1.
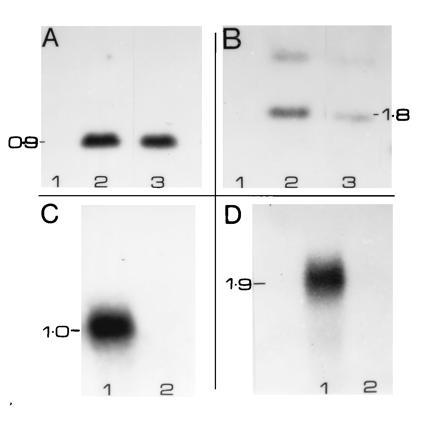
(A) Southern blot analysis of transgenic cotton. Genomic DNA (20 μg each) was isolated from the leaves of DP50 and no. 6888–7 or fibers of no. 7148, and was blotted to nitrocellulose after digestion with XbaI and HindIII restriction enzymes. These enzymes excise the coding region of the phaB gene. Blots were hybridized to 32P-labeled phaB coding region (1 × 108 cpm/μg; 5 × 105 cpm/ml) and washed under stringent conditions (0.1× SSC at 53°C). Autoradiography was done at −70°C for 72 h. Molecular sizes were determined based on standards (1 kb markers; BRL). Lanes: 1, DP50; 2, no. 6888–7; 3, no. 7148. (B) Genomic DNA of no. 6888–7, no. 7148, and DP50 were digested with XbaI and subjected to Southern blot analysis. The blot was hybridized to the coding region of phaC. XbaI digestion is expected to release a 1.8-kb coding region of phaC gene. Lanes: 1, DP50; 2, no. 7148; 3, no. 6888–7. (C) Northern blot analysis of transgenic fiber RNAs. Total RNA (20 μg each) was isolated from 15 DPA fibers of no. 7148 and DP50 control. They were size fractionated on formaldehyde/agarose gels and blotted to nitrocellulose. The blots were hybridized to 32P-labeled (1 × 108 cpm/μg; 0.5 × 105 cpm/ml) insert of phaB and washed under stringent conditions (0.1× SSC at 53°C). Molecular weights were estimated based on 1-kb marker (BRL). Lanes: 1, no. 7148; 2, DP50. (D) Fiber RNAs of no. 7148 and DP50 were hybridized to the insert of the phaC gene. All other conditions were similar to C. Lanes: 1, no. 7148; 2, DP50.
Various tissues from the transgenic plants were tested for acetoacetyl-CoA reductase activities. The fibers of 10 transgenic plants (33%), including the R1 progenies of three germ-line transformants, showed acetoacetyl-CoA reductase activities in the range of 0.07 to 0.52 μmol/min per mg protein. Leaf tissues of plants containing the E6 promoter showed 10- to 20-fold lower activities, while reductase activity was not detected in the leaf of plants containing the FbL2A promoter (not shown). The E6 and FbL2A promoters are active during different stages of fiber development. In transgenic plants containing the E6 promoter, the acetoacetyl-CoA reductase activity was detected in early fiber development (5 to 20 DPA) with maximum activity appearing during 10 to 15 DPA. In transformants containing the FbL2A promoter, the enzyme appeared in late fiber development (20 to 45 DPA) with maximal activity occurring during 35 to 40 DPA (30). Fibers of eight plants showed PHA synthase activities, as measured by radioactivity incorporation (0.3 to 2.8 × 105 cpm). All plants showed endogenous thiolase activities in the range of 0.01 to 0.03 μmol/min per mg in the fiber (not shown).
Detection and Characterization of PHB in Transgenic Fibers.
Transgenic fibers were subjected to epifluorescence microscopy after Nile blue A staining (Fig. 2). Nile blue A binding to PHB granules produces a strong orange fluorescence at the excitation wavelength of 546 nm and indicates the presence of PHB in bacteria (21). The transgenic fibers consistently showed fluorescent granules (Fig. 2 D–F). However, occasionally diffused fluorescence was observed in control DP50 fibers (Fig. 2C). Therefore, the Nile blue staining by itself did not confirm PHB in fibers.
Figure 2.
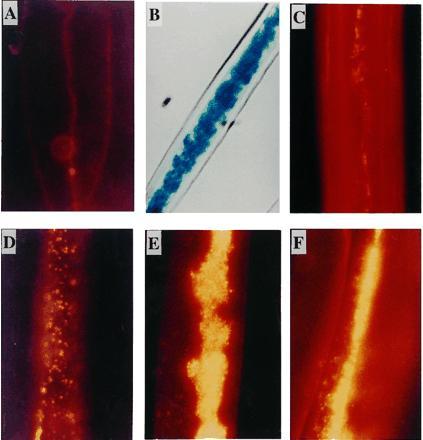
Epifluorescence microscopy of transgenic fiber. Transgenic (no. 7148) and control DP50 fibers were stained with Nile blue A and subjected to epifluorescence microscopy under excitation wavelength of 546 nm. (A) Control (DP50) 15 DPA fiber. The thin primary walls and lumen are visible. (B) Optical micrograph of transgenic no. 7148 fiber (30 DPA) that was subjected to histochemical staining for GUS to visualize the lumen. (C) Mature DP50 fiber stained with Nile blue A. (D) Transgenic 15 DPA fiber. (E and F) Mature transgenic fibers stained with Nile blue A.
Fibers from one of the transgenic plants (no. 7148) were examined by transmission electron microscopy, which has been used to visualize electron-lucent PHB granules in plants and in bacteria (9, 22). This technique was also used to measure the size and location of PHB in fibers (Fig. 3). The granules measured 0.15 to 0.3 μm in size and were present in the cytoplasm. Control fibers lacked similar clusters of granules (not shown). Comparable results were also reported in transgenic Arabidopsis containing phaB and phaC genes (9).
Figure 3.
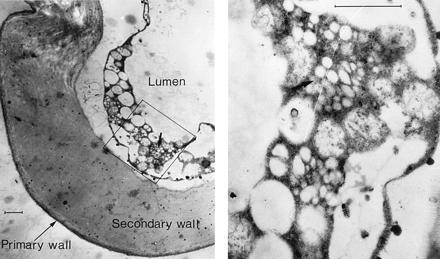
Electron-lucent PHB granules in transgenic fibers. Cotton fibers (no. 7148, 30 DPA) were fixed with 1% paraformaldehyde/2% glutaraldehyde in 0.05 M phosphate buffer (pH 7.2) for 2 h at room temperature. After washing and fixing, they were embedded in Spurrs epoxy resin. Sections (75–90 nm thick) were then placed on nickel (300 mesh) grids and stained with 2% uranyl acetate followed by Reynolds lead citrate. Transmission electron (JEOL 100C X II) micrographs of cross sections of cotton fibers are shown. Arrows indicate clusters of granules in the range of 0.15 to 0.3 μm. (Bars = 1 μm.)
Epifluorescence microscopy and transmission electron microscopy indicated the presence of granules in transgenic fibers. However, to ascertain their chemical nature, additional studies were undertaken. Quantitative methods have been developed to detect crotonic acid by HPLC after acid hydrolysis of extracted PHB (23). This method was used to detect PHB in the fiber extracts after acid hydrolysis (Fig. 4). Duplicate experiments were carried out using extracts of control DP50 fibers. The extract of control DP50 fibers did not contain crotonic acid, whereas no. 7148 extract contained a peak that corresponded to standard crotonic acid (Fig. 4). PHB content of fibers from 12 transgenic plants was measured by HPLC and estimation based on computer analysis of peak size ranged from 30 to 3440 μg/gm of dry fiber. Fibers of no. 7148 showed the highest PHB content (3440 μg/gm fiber).
Figure 4.
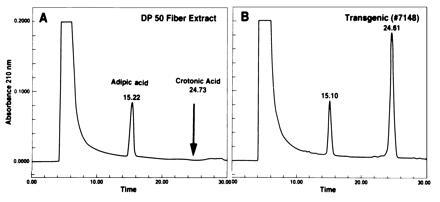
Detection of PHB by HPLC. Lyophilized immature fibers were homogenized in chloroform using a polytron. The resulting fine suspension was incubated at 65°C for 18 h, and then the chloroform was separated from debris by filtration. It was dried by N2 stream at 42°C and hydrolyzed with sulfuric acid at 90°C for 45 min before subjecting it to HPLC analysis on Aminex HPX-87H column using a Beckman System Gold Gradient HPLC system with variable wavelength detector. Quantitation was done by system gold computer software. Standard HPLC retention time for crotonic acid was obtained by subjecting crotonic acid (Sigma) to HPLC analysis (not shown). Adipic acid was used as an internal standard. (A) HPLC profile of DP50 extract. The arrow indicates the retension time for standard crotonic acid, obtained in duplicate experiment. (B) The retention time of a compound in transgenic fiber (no. 7148) extract matched with that of standard crotonic acid.
The identity of the new compound in transgenic fibers (no. 7148) was further confirmed by GC–MS analysis. Bacterial PHB can be converted to ethyl ester derivatives, and detected by GC–MS (23). Fiber extracts were treated with ethanol-chloroform-hydrochloric acid mixtures and subjected to GC analysis. Duplicate experiments were done with control DP50 extracts and bacterial PHB (Fig. 5). As seen in Fig. 5, the chloroform soluble transgenic fiber extract contained a compound with identical retention time as that of bacterial PHB derivative (Fig. 5). Transesterified DP50 fiber extract did not show any compound with a similar retention time (Fig. 5A).
Figure 5.
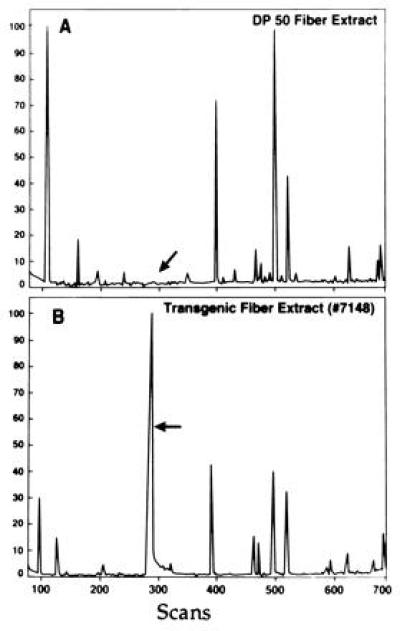
GC separation of beta-hydroxy acids from transgenic fibers. (A) PHB was extracted as described in the legend for Fig. 4 and redissolved by heating in 0.5 ml of chloroform for 10 min at 100°C. Next, 1.7 ml of ethanol and 0.2 ml of HCl were added and heated at 100°C for 4 h. The mixture was subjected to GC analysis on a Carlo-Ebra GC with an electron impact detector and a DB 5 column (50 meters; Supelco). Scans were taken at 1-sec intervals (700 total). Calibrated range was 17–600 mass units. The GC profile was then compared with that of PHB from A. eutrophus (Sigma) extracted and treated under identical conditions (not shown). The arrow marks the position of the peak present in the profile of PHB from A. eutrophus. Similar peak is not present in DP50 extract. (A) DP50 extract. (B) GC analysis of no. 7148 fiber extract. Arrow marks the peak corresponding to the ethyl ester of beta-hydroxy acid.
Analysis of ethyl ester derivatives by GC–MS confirmed that bacterial PHB and the compound from no. 7148 transgenic fibers had similar mass fragmentation patterns (not shown). They were similar to the results of previous mass spectrometry studies of bacterial and plant derived PHBs (9, 24). In addition, the mass fragmentation pattern of reference compound, ethyl ester hydroxybutyrate, was similar to that of the fiber derived material (not shown).
Thus, HPLC and GC–MS data support the conclusion that the new polymer synthesized in transgenic fibers is PHB. The molecular mass of the PHB polymer in fibers was estimated by gel permeation chromatography. PHB was fractionated, and each fraction was converted to crotonic acid and analyzed by HPLC. Based on the elution time of molecular weight standards, a major portion (68.3%) of the PHB in cotton fiber had a molecular mass of 0.6 × 106 Da or more; of this portion, 31% had a molecular mass of 1.8 × 106 Da or more (Table 1).
Table 1.
Molecular mass distribution of PHB in transgenic fiber
| Fraction no. | Molecular weight × 103 Da | Fiber
|
A. eutrophus
|
||
|---|---|---|---|---|---|
| μg | % | μg | % | ||
| 1 | >1800 | 0.65 | 2.2 | 0.0 | 0.0 |
| 2 | 1800 | 8.6 | 28.9 | 0.71 | 7.8 |
| 3 | 900 | 6.5 | 21.8 | 1.6 | 17.8 |
| 4 | 600 | 4.6 | 15.4 | 1.4 | 15.6 |
| 5 | 400 | 3.1 | 10.4 | 1.9 | 21.1 |
| 6 | 220 | 2.1 | 7.0 | 1.3 | 14.5 |
| 7 | 165 | 1.7 | 5.7 | 0.84 | 9.2 |
| 8 | 110 | 1.2 | 4.0 | 0.66 | 7.2 |
| 9 | <110 | 1.4 | 4.6 | 0.6 | 6.6 |
| Total | — | 29.85 | 100 | 9.01 | 99.8 |
| Recovery | 29.85/29.7 | 100.5% | 9.01/9.3 | 96.9% | |
PHB isolated from fibers was size-fractionated by gel permeation HPLC using ProGel TSK column (G5000-HXL; Supelco), and 0.5 ml fractions were collected. Each fraction was converted to crotonic acid by acid hydrolysis and quantitated by HPLC. Similarly a duplicate experiment was conducted using PHB granules from A. eutrophus (Sigma). Polystyrene molecular weight standards (nominal mol. wt. range of 0.11 × 106 to 1.8 × 106; Supelco) were used to calibrate the column.
Accumulation of PHB in Developing Fibers.
Quantitative estimation of PHB accumulation in developing fibers was undertaken by HPLC analysis. The amounts of PHB from the bolls of various ages from the same plant (no. 7148) were estimated by HPLC analysis of crotonic acid (Fig. 6). When the amount of PHB was displayed as a function of fiber weight, the PHB level increased up to 15 DPA and then decreased (Fig. 6A). It is likely that this decrease is due to the increase in the fiber weight occurring after 15 DPA. Deposition of large quantities of cellulose occurs during the secondary wall synthesis stage of fiber development (16–45 DPA; ref. 27). When the total weight of PHB during development was estimated per boll, as shown in Fig. 6B, no significant decrease is seen. Extraction of PHB from mature fibers is difficult and this fact may account for the small apparent decrease in PHB during fiber maturation (Fig. 6B).
Figure 6.
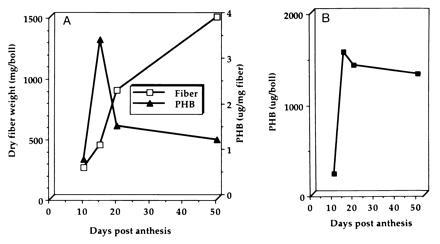
PHB accumulation during fiber development. (A) PHB accumulation in relation to fiber weight during development. PHB levels were determined from 10, 15, 20, and 50 DPA fibers of no. 7148. The increase in fiber weight per boll is also shown. The weight of PHB as a function of fiber weight decreased during fiber maturation due to increased fiber weight and not due to degradation of PHB. (B) PHB level in developing bolls. The total weight of PHB per boll during development is shown. The results show that PHB level does not decrease significantly during fiber maturation.
Thermal Properties of Polyester Containing Transgenic Fibers.
DSC measures qualitative and quantitative heat and temperature transitions by measuring the heat flow rate through the sample. The DSC measurements, when compared with DP50, indicate that the onset of decomposition of no. 7148 fiber was advanced. This result was confirmed by thermogravimetric analysis (not shown). The total heat uptake for no. 7148 (690.7 J/g) was 11.6% higher than for DP50 (619.0 J/g); as shown in Fig. 7A. The heat uptake measurements were repeated with DP50 and no. 7148 samples. The heat uptake for three independent samples of DP50 were 618.8, 620.1, and 623.5 J/g each, whereas the values of 695.3 and 692.0 J/g were obtained from repeat measurements of no. 7148 samples. Two other transgenic samples, no. 6888–7 containing 30 μg PHB per gm fiber and no. 8801 containing 423 μg PHB per gm fiber, were subjected to heat uptake measurements by DSC. Heat uptake values of 627.5 J/g and 642.3 J/g were obtained for no. 6888–7 and no. 8801, respectively. Thus, the heat uptake capacity appears to be related to the amount of PHB present in fiber. The precise relationship between thermal properties and amounts of PHB in fiber can be established as fibers with greater PHB content are developed. The transgenic fibers (no. 7148) were spun into yarn by miniature spinning (Starlab VY-5 direct sliver-to-yarn spinning frame) and knitted into cloth by a knitting machine. Unbleached and undyed fabric were then subjected to thermal property measurements by DSC along with the control fabric (DP50). The DSC measurements showed a heat uptake of 695.4 J/g for no. 7148 and 617.8 J/g for DP50 fabric, respectively (not shown).
Figure 7.
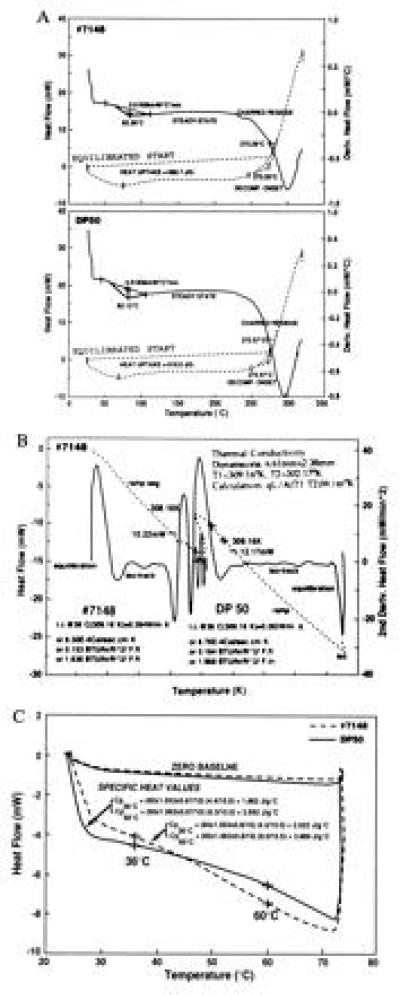
(A) Heat uptake of transgenic fibers. The heat gain/loss characteristics of fibers were measured by DSC. Sample sizes of 10 mg (no. 7148) and 10.06 mg (DP50) were used. (B) Thermal conductivity measurements of transgenic fibers. The heat flow rate through samples no. 7148 and control DP50 fibers was monitored against temperature using steady-state equilibrium heat flow method. A modified DSC (TA Instruments) was used for the thermal conductivity measurements. Experimental conditions and results are summarized in the figure. All conditions for DP50 were identical to those shown for no. 7148, except: dimension = 6.61 mm × 2.36 mm and T2 = 302.18 K. T1, temperature at the bottom of the sample; T2, temperature at the top of the sample; q, heat flow; L, heat flow path; A, total area of the sample. (C) Comparison of specific heat values of transgenic and control fibers by DSC. Specific heat values were obtained for 36°C and 60°C each. A sample size of 10 mg was used in the DP50 control, whereas 10.5 mg was used for no. 7148. Zero baseline was established by measurement of Sapphire. Cell constant values are E36°C = (0.7978 × 10 × 60.83)/(60 × 0.8 × 9.6) = 1.053. E60°C = (0.8432 × 10 × 60.83)/(60 × 0.8 × 10.05) = 1.063.
The relative heat transmission capacities of no. 7148 and DP50 fibers were determined by thermal conductivity (TC) measurements (Fig. 7B). The TC of no. 7148 fibers (0.264 W/m°K) was 6.7% lower than DP50 (0.283 W/m°K), indicating slower cooling down of the material. Thus, the DP50 fibers have faster heat dissipation properties.
The heat retention of samples was determined by specific heat measurements at two temperatures (36°C and 60°C) and are shown in Fig. 7C. Sample no. 7148 showed a 8.6% higher heat retention than the DP50 sample at 36°C, while the difference was 44.5% higher for transgenic fiber at 60°C (Fig. 7C). Thus, these results agree with the TC measurements and confirm that sample no. 7148 has higher heat capacity.
DISCUSSION
A number of different genetic strategies are obvious for the modification of physical and chemical properties of cotton fiber. Existing fiber properties, such as strength and length, may be enhanced by modifying corresponding genes. However, so far none of the genes responsible for various fiber properties have been identified. Therefore, another plausible strategy is to identify already characterized genes from animals, plants, or bacteria that may have the potential to modify fiber, such as synthesis of structural proteins, new enzyme(s) to use existing substrates, or enzymes that create new substrates and new products. Synthesis of PHB in fiber is an example of introducing new enzymes to use an existing substrate, acetyl-CoA, to produce a new polymer in fiber.
Metabolic pathway engineering for synthesis of PHB in fiber required only the integration of functional phaB and phaC genes, since β-ketothiolase, which is involved in the synthesis of mevalonate, is ubiquitous in plants. We introduced phaB and phaC along with GUS gene into cotton by particle bombardment. Eight of the transformants identified in this study expressed all three genes, phaB, phaC, and GUS. This result provides conclusive evidence that particle bombardment is suitable for introducing several genes into a single plant. This is in contrast to Agrobacterium-mediated transformation that has been routinely used to transform cotton (28). It is limited by the number of genes that can be accommodated on the Ti-based vector DNA, as well as limited to cultivars that can be regenerated through tissue culture. Thus, particle-mediated transformation is ideal to genetically engineer plants with more complex pathways involving many genes.
To avoid any detrimental effects of PHB production during plant growth, one of the genes, phaB, was expressed predominantly in fiber. Reductions in growth and seed production were reported in transgenic Arabidopsis as a result of phaB overexpression in leaves and other tissues (9). All transgenic cotton expressing phaB and phaC showed normal growth and morphology. The temporal regulation of phaB in fiber conforms to the known characteristics of corresponding E6 and FbL2A promoters. The optimal levels of reductase and PHA synthase activities, as well as their time of expression in fiber, are likely to be important for maximum synthesis of PHB. From the limited number of plants studied, it appears that high levels of reductase and PHA synthase levels during early fiber development are conducive for PHB synthesis. Transgenic plants, which contained the promoter FbL2A (active in later fiber development), showed moderately high levels of acetoacetyl-CoA reductase and PHA synthase activities, but did not result in high levels of PHB (not shown). It is possible that decreased levels of acetyl-CoA during late fiber development may be a contributing factor for reduced PHB synthesis.
As previously shown for transgenic Arabidopsis (9), the transgenic cotton that expressed phaB and phaC genes produced a high molecular weight polymer whose chemical identity matches that of PHB produced by bacteria. The PHB synthesis was detected as early as 10 DPA. No significant decrease in the total amount of PHB occurred during fiber maturation. Thus, it appeared that no depolymerase activity was present in fiber to degrade PHB as in bacteria (29). We have not detected any decrease in PHB levels in fibers stored at room temperature for several months (not shown). However, the stability of PHB in cotton fiber in finished textile products has not been explored yet.
Acetyl-CoA is a critical component of various metabolic pathways in the cell. Partial utilization of the acetyl-CoA pool for the synthesis of a new polymer seemed to have no detrimental effect on cotton fiber properties as the fiber exhibited normal strength, length, and micronaire (not shown). On the other hand, the new polymer enhanced fiber properties, as shown by the studies of thermal characteristics. The transgenic cotton fibers exhibited measurable changes in thermal properties that suggested enhanced insulation characteristics. The transgenic fibers conducted less heat, cooled down slower, and took up more heat than conventional cotton fibers. Modified fibers with superior insulating properties may have applications in winter wear or other textile uses where enhanced insulating properties are advantageous. However, the changes in thermal properties are relatively small as expected from the small amounts of PHB in fibers (0.34% fiber weight). It is likely that a severalfold increase in PHB synthesis is required for product applications. Nevertheless, the positive changes in fiber qualities demonstrated here are indication of potential of this technology. As new generations of fibers are developed through genetic engineering, they will impact the growth of the textile industries that are significant segments of economies of many countries.
Acknowledgments
We acknowledge the expert technical help of Jennifer Rinehart, Lori Spatola, and Michael Petersen. We are thankful to Dr. B. Chowdhury (MATECH Associates) for conducting the thermal studies and useful discussions. We are grateful to Cheryl Scadlock and Peggy Wagner for editorial assistance. Permission for the use of A. eutrophus genes was obtained from Metabolix (Cambridge, MA). This study was partially funded by the Department of Commerce, National Institute of Technology, Advanced Technology Program Grant 7ONANB5H1061.
Footnotes
Abbreviations: PHB, poly-d-(−)-3-hydroxybutyrate; DPA, days post anthesis; GUS, β-glucuronidase; PHA, polyhydroxyalkanoate; TC, thermal conductivity; DSC, differential scanning calorimeter; GC, gas chromatography; MS, mass spectroscopy.
References
- 1.Meredith W R. Cotton Fiber Cellulose: Structure, Function and Utilization. Memphis, TN: Natl. Cotton Coun.; 1992. pp. 289–302. [Google Scholar]
- 2.John M E. CHEMTECH. 1994;24:27–30. [Google Scholar]
- 3.Arthur J C. In: Polymers: Fibers and Textiles, A Compendium. Kroschwitz J I, editor. New York: Wiley; 1990. pp. 118–141. [Google Scholar]
- 4.Ryser U. Eur J Cell Biol. 1985;39:236–256. [Google Scholar]
- 5.Basra A S, Malik C P. Int Rev Cytol. 1984;89:65–113. [Google Scholar]
- 6.Steinbuchel A. Acta Biotechnol. 1991;5:419–427. [Google Scholar]
- 7.Steinbuchel A, Hustede E, Liebergesell M, Pieper U, Timm A, Valentin H. FEMS Microbiol Rev. 1992;9:217–230. doi: 10.1111/j.1574-6968.1992.tb05841.x. [DOI] [PubMed] [Google Scholar]
- 8.Anderson A J, Dawes E A. Microbiol Rev. 1990;54:450–472. doi: 10.1128/mr.54.4.450-472.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Poirier Y, Dennis D, Klomparens K, Somerville C. Science. 1992;256:520–523. doi: 10.1126/science.256.5056.520. [DOI] [PubMed] [Google Scholar]
- 10.John M E, Crow L J. Proc Natl Acad Sci USA. 1992;89:5769–5773. doi: 10.1073/pnas.89.13.5769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.John M E, Keller G L. Plant Physiol. 1995;108:669–676. doi: 10.1104/pp.108.2.669. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Current Protocols in Molecular Biology. New York: Wiley; 1987. [Google Scholar]
- 13.Peoples O P, Sinskey A J. J Biol Chem. 1989;264:15293–15297. [PubMed] [Google Scholar]
- 14.Peoples O P, Sinskey A J. J Biol Chem. 1989;264:15298–15303. [PubMed] [Google Scholar]
- 15.Nishimura T, Saito T, Tomita K. Arch Microbiol. 1978;116:21–27. doi: 10.1007/BF00408729. [DOI] [PubMed] [Google Scholar]
- 16.Saito T, Fukui T, Ikeda F, Tanaka Y, Tomita K. Arch Microbiol. 1977;114:211–217. doi: 10.1007/BF00446864. [DOI] [PubMed] [Google Scholar]
- 17.Schubert P, Steinbuchel A, Schlegel H G. J Bacteriol. 1988;170:5837–5847. doi: 10.1128/jb.170.12.5837-5847.1988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.McCabe D E, Martinell B J. Bio/Technology. 1993;11:596–598. [Google Scholar]
- 19.Christou P, McCabe D E, Martinell B J, Swain W F. Trends Biotechnol. 1990;80:145–151. [Google Scholar]
- 20.Jefferson R A. Plant Mol Biol Rep. 1987;5:387–405. [Google Scholar]
- 21.Ostle A G, Holt J G. Appl Environ Microbiol. 1982;44:238–241. doi: 10.1128/aem.44.1.238-241.1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hustede E, Steinbuchel A, Schlegel H G. FEMS Microbiol Lett. 1992;93:285–290. doi: 10.1016/0378-1097(92)90476-5. [DOI] [PubMed] [Google Scholar]
- 23.Karr D B, Waters J K, Emerich D W. Appl Environ Microbiol. 1983;46:1339–1344. doi: 10.1128/aem.46.6.1339-1344.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Findlay R H, White D C. Appl Environ Microbiol. 1983;45:71–78. doi: 10.1128/aem.45.1.71-78.1983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hill J O. For Better Thermal Analysis and Calorimetry. 3rd Ed. New Castle, Australia: Intl. Confed. for Thermal Analysis; 1991. pp. 6–50. [Google Scholar]
- 26.John M E. In: Industrial Biotechnological Polymers. Gebelein C G, Carraher C E, editors. Lancaster, PA: Technomic; 1995. pp. 69–79. [Google Scholar]
- 27.Meinert M C, Delmer D P. Plant Physiol. 1977;59:1088–1097. doi: 10.1104/pp.59.6.1088. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Umbeck P, Johnson G, Barton K, Swain W. Bio/Technology. 1987;5:263–266. [Google Scholar]
- 29.Senior P J, Dawes E A. Biochem J. 1973;134:225–238. doi: 10.1042/bj1340225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Rinehart, J. A., Petersen, M. W. & John, M. E. (1996) Plant Physiol., in press. [DOI] [PMC free article] [PubMed]