

Abstract
G proteins regulate intracellular signaling by coupling a cycle of guanine nucleotide binding and hydrolysis to transient changes of cellular functions. The mechanisms that control the recycling of transducin, the “pace-setting” G protein that regulates mammalian phototransduction, are unclear. We show that a novel retinal specific RGS-motif protein specifically binds to an intermediate conformation involved in GTP hydrolysis by transducin and accelerates phosphate release and the recycling of transducin. This specific interaction further rationalizes the kinetics of the phototransduction cascade and provides a general hypothesis to explain the mechanism of interaction of RGS proteins with other G proteins.
Keywords: phototransduction, G protein, retina, recovery
The heterotrimeric G protein transducin (TD) mediates signal transduction in vertebrate photoreceptor cells. Photon absorption activates rhodopsin and it catalyzes the binding of GTP to the α subunit of transducin (TDα) and the dissociation of the βγ subunits (TDβγ). GTP-bound transducin (TDαGTP) activates a cyclic GMP phosphodiesterase (cG-PDE) by sequestering its γ subunit. The activated phosphodiesterase lowers cyclic GMP levels and thus modifies ion channel function and initiates an electrical response in the photoreceptor (1, 2). A variety of mechanisms are involved in terminating this signal. One process involves the hydrolysis of GTP bound to TDα. This is a relatively slow step in vitro, and the process is accelerated in vivo. However, the precise mechanism of acceleration is not clear. The γ subunit of cG-PDE that interacts with TDα may be involved (3), and there may be additional protein components that control the rate of GTP hydrolysis and thus the conformational states of TDα and its recycling (4). A candidate protein that could be involved in accelerating the GTP hydrolysis and the recycling of TD has emerged from recent studies of proteins containing the RGS (regulator of G protein signaling) domain (5). The initial member of this gene family, SST2, was discovered in yeast (6). Mutations in this gene increased the sensitivity of yeast cells to pheromone. Genetic experiments suggested that the SST2 protein interacted with the yeast G protein α subunit (GPA-1) and negatively regulated its activity (7). An homologous family of genes was found in Caenorhabditis elegans and in mammalian systems. All members of this family share amino acid sequence homology with SST2, and genetic evidence suggests that they interact with the G protein α subunit (Gα; refs. 8 and 9). One of these RGS-proteins, GAIP, was shown to bind specifically to the Gαi3 protein (10). More recently, evidence has been presented that both GAIP and another member of the family, RGS-4, can catalyze GTP hydrolysis by some Gα proteins, namely the Gαi family but not by Gαs (11).
We report here a specific interaction of one intermediate conformation of transducin with a novel retinal specific RGS protein, RGS-r. Our data show that RGS-r facilitates TD recycling by enhancing GTP hydrolysis through binding a conformation of the protein that stabilizes the transition state. These findings are significant for understanding the mechanisms of regulation of G protein-coupled signal transduction by RGS proteins.
MATERIALS AND METHODS
Reverse Transcription PCR.
Total mouse retinal RNA was prepared by RNeasy kit (Qiagen, Chatsworth, CA), according to the manufacturer’s instructions. One microgram of the RNA was randomly primed and reverse-transcribed using a cDNA cycling kit (Stratagene). A volume 1/20 of that of the reverse transcription reaction mixture was used as template for the subsequent PCR. The sequence of the 5′ primer (rgs-f1) is: AAAGAATTCGA(G/A)GA(G/A)AA(T/C)ATIGA(T/C)TT(T/C)TGG. The sequence of the 3′ primer (rgs-b1) is: AAAGGATCCIC(G/T)(A/G)TAI(C/G)(A/T)(A/G)TC(T/C)TT(T/C)TCCAT. The PCR was performed in 50 μl of reaction mixture containing template, 10 mM Tris·HCl (pH 8.3), 50 mM KCl, 1.5 mM MgCl2, 200 μM each of dATP, dCTP, dGTP, and dTTP, 5 units of Taq polymerase, and 1 μM primers. The PCR was performed using the following conditions: 94°C for 3 min, followed by 35 cycles of 94°C for 1 min, 48°C for 1 min, and 72°C for 1 min, and a final incubation of 72°C for 10 min. The 250-bp products were subcloned into pGEM-T (Promega). Clones that generated a 250-bp insert after EcoRI and BamHI restriction digestion were further sequenced to determine their identities.
Northern Blot Analysis.
One microgram each of poly(A)+ RNA from various rat tissues (CLONTECH) was blotted onto nylon membranes (Amersham) and hybridized to probes labeled by random priming the coding region of the rat RGS-r cDNA fragment. The specific activity of the 32P-labeled probe is >109 cpm/μg. ExpressHyb solution (CLONTECH) was used under high-stringency conditions for hybridization (65°C), and high-stringency washes were performed in 0.1× SSC (15 mM NaCl/15 mM sodium citrate, pH 7.0) plus 0.3% SDS at 65°C.
Protein Purification.
Rod outer segment (ROS) membranes were prepared from bleached bovine retina as described (12). Transducin was separated from the membranes by hypotonic elution in the presence of 100 μM GTP or GTP[γS], and the subunits were resolved by affinity chromatography on Blue Sepharose (Bio-Rad; ref. 13). Urea-treated ROS membranes were prepared from the eluted membranes as described (14).
For expression and purification of RGS-r protein, a 5′-flanking NdeI site and a 3′-flanking BamHI site were introduced into the mRGS-r coding region by standard PCR techniques. The fragment was subcloned into the NdeI and BamHI sites of pET15B (Novagen). Escherichia coli strain BL21(DE3) was used as an expression host. Cells were grown on Luria broth medium at 30°C to OD600 = 0.4, then the expression of mRGS-r was induced by adding isopropyl β-d-thiogalactoside (IPTG) to a final concentration of 1 mM. Cells were harvested 3 hr after induction and were sonicated in buffer A (20 mM Tris·HCl, pH 8.0/100 mM NaCl/2 mM MgCl2/6 mM 2-mercaptoethanol/5% glycerol). The soluble extract was applied to a 2-ml Ni2+-NTA Superflow column (Qiagen). The column was washed extensively with buffer A and buffer A containing 25 mM imidazole, and the mRGS-r protein was eluted with a linear gradient of 25–400 mM imidazole in buffer A. To obtain a fair comparison for the activity of mRGS-r with RGS-4, the mRGS-r used in Fig. 6 was denatured by buffer A containing 5 M urea, renatured by removing urea while bound to the Ni2+-NTA column, washed, and eluted as described above.
Figure 6.
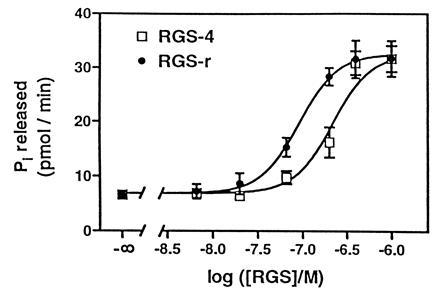
Comparison of the effect of mRGS-r protein and RGS-4 protein on GTP hydrolysis by TD. The hydrolysis of GTP (10 μM) by 200 nM TD plus 10 μg of urea-treated ROS membranes was determined at the indicated concentrations of mRGS-r protein (•) or RGS-4 protein (□) after 10 min of incubation at 30°C. The data shown are mean ± SD of triplicate determinations.
Rat RGS-4 cDNA was cloned by reverse transcription-PCR from rat brain poly(A)+ RNA (CLONTECH). The coding region was subcloned into pET15B using PCR-introduced NdeI and BamHI sites. We solubilized the protein in 5 M urea, loaded the denatured RGS-4 onto a Ni2+-NTA column, and then washed the urea away to allow the renaturation of RGS-4. The renatured RGS-4 was then washed with buffer A containing 25 mM imidazole and eluted as described above for RGS-r. The purity of recombinant RGS-4 and mRGS-r was >90%, as judged by SDS/PAGE.
Transducin–RGS-r Interaction Assayed by Gel Filtration.
Purified TDαGDP (6 μM) was incubated in buffer A (total volume of 200 μl) with 18 μM mRGS-r for 10 min at 30°C in the absence and presence of aluminum magnesium fluoride (AMF; 100 μM AlCl3/6 mM MgCl2/10 mM NaF). Thereafter, the mixture was applied to a Superose 12 column (Pharmacia) equilibrated with buffer A with or without AMF. Proteins were eluted at a flow rate of 0.5 ml/min and 0.25-ml fractions were collected. One hundred and fifty microliters of each protein-containing fraction was concentrated to 10 μl using a Minicon-10 device (Amicon) and applied to SDS/PAGE (10% polyacrylamide in resolving gel). Proteins were visualized by staining with Coomassie brilliant blue. To calculate the relative molecular sizes of the proteins and the complex, the column was calibrated using a mixture of immunoglobulin G (160 kDa), human serum albumin (67 kDa), β-lactoglobulin (35 kDa), and cytochrome c (12 kDa).
Transducin–RGS-r Interaction Assayed by Column Trapping.
TDαGTPγS or heterotrimeric TDαβγ (3.5 μM each) were incubated in buffer A with or without 10 μM mRGS-r in a total volume of 100 μl for 10 min at 30°C in the absence and presence of AMF. Thereafter, the mixtures were applied to Ni2+-NTA Superflow (Qiagen) columns (bed volume, 100 μl) equilibrated with buffer A. The columns were washed with 1 ml of buffer A containing 25 mM imidazole. For samples containing AMF during incubation, AMF was also added to the washing buffer. Bound proteins were eluted with 400 μl of buffer A containing 400 mM imidazole, and the eluate was concentrated to 10 μl. Proteins were visualized by staining with Coomassie brilliant blue after SDS/PAGE (10% polyacrylamide in resolving gel).
Guanine Nucleotide Binding Assay and GTPase Assay.
Reconstitution of rhodopsin–TD interaction was performed in a reaction mixture (100 μl) containing 50 mM triethanolamine·HCl (pH 7.4), 100 mM NaCl, 5 mM MgCl2, 0.1 mg of bovine serum albumin per ml, 0.5 μM TDαGDP, 0.5 μM TDβγ, and 10 μg of urea-treated ROS membranes. For determination of GTP hydrolysis, the reaction mixture was equilibrated for 5 min at 30°C. Reaction was initiated by addition of 10 μM [γ-32P]GTP (0.2 μCi; 1 Ci = 37 GBq) and conducted for the indicated periods of time. Termination of reaction and determination of Pi released were performed as described (15). Under each condition used, Pi release was found to be linear with time (data not shown). For determination of GTP analog binding, reaction was initiated by addition of either 100 nM GTP[γ35S] (0.5 μCi) or 1 μM [3H]Gpp[NH]p (0.5 μCi) and conducted for 20 min at 30°C. The amount of binding was determined as described (16).
RESULTS
Identification of RGS-r.
To search for possible involvement of an RGS-like protein in the phototransduction cascade, we looked for a retinal specific member of the family, since many of the proteins involved in this signal transduction process are preferentially expressed in the retina. We sequenced 40 reverse transcription-PCR products and found three distinct sequences that contained the RGS domain: one identical to RGS-4 (8, 9), another identical to RGS-8, and another one (mRGS-r) was not identical to any known RGS sequence. To obtain the full-length cDNA for mRGS-r, we screened a mouse retinal cDNA library that was generously provided by Wolfgang Baehr (University of Utah). We obtained four positive clones, and one of them, mRGS-r3, corresponding to 2.2 kb, was found to have an open reading frame of 201 aa. The sequence flanking the initiation codon matched the Kozak sequence (17), and a mRNA corresponding to 2.2 kb was detected in the retina (Fig. 1), indicating that mRGS-r3 is probably a full-length cDNA clone. The rat RGS-r (rRGS-r) cDNA was obtained by RT-PCR from rat poly(A)+ RNA (CLONTECH) using primers specific for mRGS-r. Northern blot analysis shows that rRGS-r is expressed predominantly in the retina, where it is found in two forms with mobilities corresponding to 2.2 kb (major form) and 3.2 kb (minor form). However, the 2.2-kb form is also found at relatively low levels in the liver and is barely detectable in the other tissues that were tested. The derived amino acid sequences of rRGS-r and mRGS-r are shown compared with the sequences of the GAIP and RGS-4 proteins (Fig. 2). rRGS-r and mRGS-r are highly homologous, showing <6% difference in amino acid sequence. On the other hand, the RGS-4 clone shows much more divergence (45% overall identity), and the GAIP clone diverges even further from the RGS-r clones.
Figure 1.
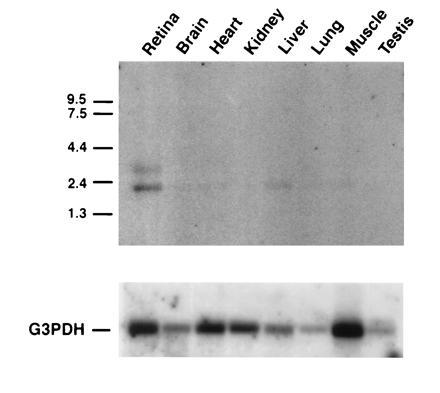
Northern blot analysis of 1 μg of poly(A)+ RNA from various rat tissues (CLONTECH) hybridized with rRGS-r probe (Upper). Rat glyceraldehyde-3-phosphate dehydrogenase (G3PDH) probe (CLONTECH) was used as a control (Lower).
Figure 2.

Sequence alignment of mouse RGS-r (mRGS-r), rat RGS-r (rRGS-r), rat RGS-4 (rRGS-4), and GAIP.
RGS-r Protein Interacts with an Intermediate Conformation of TD.
We next looked for the interaction of RGS-r with TD. We prepared the (His)6-tagged mRGS-r protein by overexpression in E. coli and purified it by affinity chromatography. The ability of the recombinant mRGS-r protein to interact with purified TD was then examined. Fig. 3 shows the results of gel filtration experiments designed to measure the mobilities of mixtures of GDP-bound α transducin (TDαGDP) and mRGS-r protein in the presence and absence of AMF. We observed a distinct mobility shift in the presence but not in the absence of AMF, which indicates that only the GDP·AMF form of transducin (TDαGDP·AMF) is capable of binding to mRGS-r. The stoichiometry of the complex between mRGS-r and TDαGDP·AMF appears to be one to one as judged by the intensities of both species on SDS/PAGE and by the apparent molecular weight of the complex as determined by its mobility on gel filtration. In other experiments shown in Fig. 4, a Ni2+-NTA column was used to trap mRGS-r protein to examine its binding to TDα under various conditions. We found that mRGS-r protein bound TDαGDP only in the presence of AMF. In the absence of AMF, both TDαGDP and TDαGTPγS were not bound to mRGS-r protein and thus did not get trapped on the column.
Figure 3.
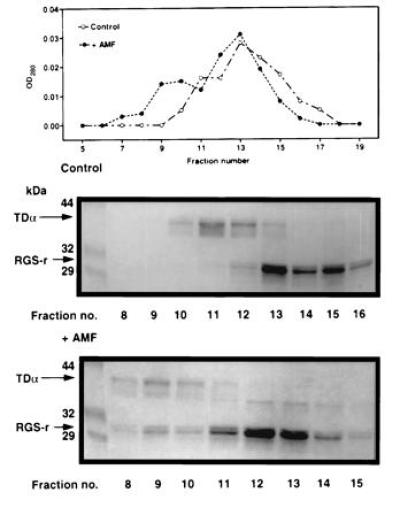
Complex formation of mRGS-r protein with TDαGDP·AMF analyzed by gel filtration. Purified TDαGDP (6 μM) was incubated with 18 μM mRGS-r for 10 min at 30°C in the absence (○ and Middle) and presence of AMF (• and Bottom). Fractions in the protein profile (Top) were analyzed by SDS/PAGE, followed by Coomassie brilliant blue staining. Complex formation of mRGS-r protein with TDα was observed in the presence of AMF. The calculated molecular sizes were 40 kDa, 35 kDa, and 72.5 kDa for TD, mRGS-r protein, and the complex, respectively.
Figure 4.
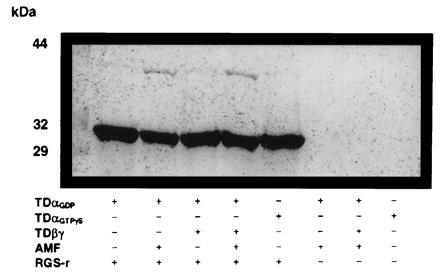
Complex formation of mRGS-r protein with TDαGDP·AMF analyzed by binding to Ni2+-NTA. Experiments were performed as described. Only the TDαGDP·AMF was found to bind to the matrix by its interaction with mRGS-r protein.
RGS-r Protein Enhances GTP Hydrolysis by TDα but Does Not Affect GTP Analog Binding.
Fig. 5A shows that mRGS-r protein can increase the rate of Pi release >6-fold in the presence of bleached rhodopsin, GTP, and heterotrimeric TD. Using 50 pmol of TD, we observed Pi release of ≈75 pmol/min, indicating that each TD had turned over more than once. Moreover, the maximal effect of mRGS-r protein is reached at 20 pmol, indicating that it acts substoichiometrically (Fig. 5B). Furthermore, the mRGS-r protein does not change GTP analog binding to TDα, indicating that receptor·G protein coupling is not affected (Fig. 5 C and D), and the mRGS-r protein does not bind tightly to TD loaded with nonhydrolyzable GTP analogs (Fig. 5E).
Figure 5.
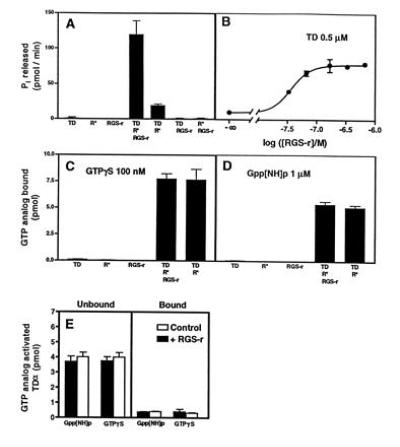
Enhanced GTP hydrolysis of TDα by mRGS-r protein without affecting the loading of GTP. (A) mRGS-r protein increases the Pi release from TDα. The hydrolysis of GTP (10 μM) by 0.5 μM TD, 10 μg of urea-treated ROS membranes (R*, bleached rhodopsin in rod outer segment membranes), 0.7 μM mRGS-r, or combinations of these components was measured for 3 min at 30°C. Hydrolysis of GTP was observed in the presence of R* and TD. mRGS-r protein increased the Pi release from TD ≈7-fold. (B) Dose–response curve of mRGS-r protein. Half-maximal and maximal stimulation by mRGS-r protein was observed at 50 and 200 nM, respectively. (C and D) The binding of nonhydrolyzable GTP analogs to TD was not affected by mRGS-r protein. GTP[γS] (100 nM) or Gpp[NH]p (1 μM) was used as described. (E) TD activated by nonhydrolyzable GTP analogs does not bind to mRGS-r protein. TD loaded with labeled nonhydrolyzable GTP analogs was prepared by centrifugation of the reaction mixtures from C and D. The supernatants, containing the labeled TD and mRGS-r protein, were applied to Ni2+-NTA columns. The amount of protein-bound radioactivity in the flow-through and the 400 mM imidazole eluate was determined. In all parts of this figure, means ± SD of triplicate determinations are shown.
RGS-r Is More Effective than RGS-4 in Activating GTP Hydrolysis by TD.
To address the specificity of mRGS-r protein for TD, we reconstituted the system with another RGS protein, RGS-4, which is similar to RGS-r but is expressed primarily in brain and has been shown to interact with Gαo and to Gαi. Because >95% of (His)6-tagged RGS-4 expressed in BL21 (DE3) is insoluble, we solubilize it in 5 M urea and renature it by washing away the urea. mRGS-r protein was treated the same way to get a fair comparison. Fig. 6 shows that both mRGS-r and RGS-4 proteins facilitate GTP hydrolysis. However, mRGS-r protein is at least 3-fold more effective than RGS-4 protein. Thus, we suggest that RGS-r plays an important role in the phototransduction cascade by regulating the lifetime and effective concentration of activated TDα.
DISCUSSION
We have presented data that point to a specific interaction between RGS-r protein and an intermediate conformation of TDα, the TDαGDP·AMF (Figs. 3 and 4). AMF has been shown to bind to TDαGDP and to mimic the presence of the phosphate group that is released as a result of hydrolysis of GTP by TDα. The crystal structures of TDαGDP·AMF and Gαi1GDP·AMF suggest that these complexes reflect a conformation of the protein involved in stabilizing the transition state that is formed during the hydrolysis of GTP (18, 19). By binding and stabilizing this conformational state of TDα, mRGS-r protein does accelerate the rate of GTP hydrolysis (Figs. 5 and 6). One plausible interpretation of the data that would be consistent with previous thinking about the mechanisms of TD function is that different proteins bind to and stabilize distinct conformations of the TDα protein at different stages in its interaction with guanine nucleotide. Transducin in the TDαGDP conformation is stabilized by binding to the TDβγ complex forming the heterotrimer. The activated receptor, photolyzed rhodopsin, has been shown to bind to the nucleotide-free form of TD (20). The activated TDαGTP conformation binds the γ subunit of cG-PDE. Our work indicates that the RGS-r protein specifically interacts with another conformation of TD, one that has been implicated in stabilizing the transition state and enhancing GTP hydrolysis, thus terminating the activation of TDα. Evidence has been presented to suggest that the rate-limiting step in cycling TDα from the activated form is the release of Pi after hydrolysis of GTP (21, 22). By interacting with a transition state-stabilizing conformation of TDα, we postulate that the RGS-r protein may accelerate both the GTPase activity and Pi release.
Recently, data have been presented showing that different RGS proteins bind to the transition state conformation of members of the Gαi family and accelerate their ability to hydrolyze GTP (23). Furthermore, a specific Gαz-interacting RGS protein has been found (24), suggesting that individual RGS family members regulate certain G-protein circuits. Our results further demonstrate this principle. In the context of the phototransduction cascade, RGS-r may play a very special role in regulating the kinetics of signaling. In vitro experiments indicate that GTP hydrolysis by TDα is too slow to account for the rapid shut-off of the photoresponse observed in vivo. The finding that RGS-r enhances GTP-hydrolysis by TD may help to resolve the difference. However, we need to study the relationship between RGS-r and the γ subunit of cG-PDE. They may act sequentially on TD and there could be other factors or modulators of RGS-r that further regulate its activity. The precise role of RGS-r in vivo will ultimately be determined by extensive reconstitution experiments and genetic analysis.
Acknowledgments
We thank Dr. Wolfgang Baehr for mouse retinal cDNA library and members of the Simon lab and, in particular, Bo Yu for stimulating discussion. This work was supported from the National Institute on Aging (AG 12288). T.W. is the recipient of a fellowship from the Deutsche Forschungsgemeinschaft.
Footnotes
References
- 1.Yarfitz S, Hurley J B. J Biol Chem. 1994;269:14329–14332. [PubMed] [Google Scholar]
- 2.Lagnado L, Baylor D. Neuron. 1992;8:995–1002. doi: 10.1016/0896-6273(92)90122-t. [DOI] [PubMed] [Google Scholar]
- 3.Arshavsky V Y, Bounds M D. Nature (London) 1992;357:416–417. doi: 10.1038/357416a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Angleson K A, Wensel T G. Neuron. 1993;11:939–949. doi: 10.1016/0896-6273(93)90123-9. [DOI] [PubMed] [Google Scholar]
- 5.Roush W. Science. 1996;271:1056–1058. doi: 10.1126/science.271.5252.1056. [DOI] [PubMed] [Google Scholar]
- 6.Dietzel C, Kurjan J. Mol Cell Biol. 1987;7:4169–4177. doi: 10.1128/mcb.7.12.4169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Dohlman H G, Apaniesk D, Chen Y, Song J, Nusskern D. Mol Cell Biol. 1995;15:3635–3643. doi: 10.1128/mcb.15.7.3635. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Koelle M R, Horvitz H R. Cell. 1995;84:115–125. doi: 10.1016/s0092-8674(00)80998-8. [DOI] [PubMed] [Google Scholar]
- 9.Druey K M, Blumer K J, Kang V H, Kerl J H. Nature (London) 1996;379:742–745. doi: 10.1038/379742a0. [DOI] [PubMed] [Google Scholar]
- 10.De Vries L C, Mousli H, Wurmser A, Farquhar M G. Proc Natl Acad Sci USA. 1995;92:11916–11920. doi: 10.1073/pnas.92.25.11916. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Berman D M, Wilkie T M, Gilman A G. Cell. 1996;86:445–452. doi: 10.1016/s0092-8674(00)80117-8. [DOI] [PubMed] [Google Scholar]
- 12.Papermaster D S, Dreyer W J. Biochemistry. 1974;13:2438–2444. doi: 10.1021/bi00708a031. [DOI] [PubMed] [Google Scholar]
- 13.Wieland T, Ulibarri I, Gierschik P, Jakobs K H. Eur J Biochem. 1991;196:707–716. doi: 10.1111/j.1432-1033.1991.tb15869.x. [DOI] [PubMed] [Google Scholar]
- 14.Wieland T, Ulibarri I, Gierschik P, Hall A, Aktories K, Jakobs K H. FEBS Lett. 1990;274:111–114. doi: 10.1016/0014-5793(90)81342-l. [DOI] [PubMed] [Google Scholar]
- 15.Gierschik P, Steisslinger M, Sidiropoulos D, Herrmann E, Jakobs K H. Eur J Biochem. 1989;183:97–105. doi: 10.1111/j.1432-1033.1989.tb14901.x. [DOI] [PubMed] [Google Scholar]
- 16.Wieland C, Jakobs K H, Wieland T. Cell Signalling. 1994;6:487–492. doi: 10.1016/0898-6568(94)90002-7. [DOI] [PubMed] [Google Scholar]
- 17.Kozak M. Nucleic Acids Res. 1987;15:8125–8148. doi: 10.1093/nar/15.20.8125. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sondek J, Lambright D G, Noel J P, Hamm H E, Sigler P B. Nature (London) 1994;372:276–279. doi: 10.1038/372276a0. [DOI] [PubMed] [Google Scholar]
- 19.Coleman D E, Berghuis A M, Lee E, Linder M E, Gilman A G, Sprang S R. Science. 1994;265:1405–1412. doi: 10.1126/science.8073283. [DOI] [PubMed] [Google Scholar]
- 20.Kahlert M, Koenig B, Hofmann K P. J Biol Chem. 1995;265:18928–18932. [PubMed] [Google Scholar]
- 21.Ting T D, Ho Y K. Biochemistry. 1991;30:8996–9007. doi: 10.1021/bi00101a013. [DOI] [PubMed] [Google Scholar]
- 22.Vuong T M, Chabre M. Nature (London) 1990;346:71–74. doi: 10.1038/346071a0. [DOI] [PubMed] [Google Scholar]
- 23.Watson N, Linder M E, Druey K M, Kehrl J H, Blumer K J. Nature (London) 1996;383:172–175. doi: 10.1038/383172a0. [DOI] [PubMed] [Google Scholar]
- 24.Hunt T W, Fields T A, Casey P J, Peralta E G. Nature (London) 1996;383:175–177. doi: 10.1038/383175a0. [DOI] [PubMed] [Google Scholar]