

Abstract
Mouse clones were produced by serial nuclear transfer commencing with the transfer of four-cell nuclei at metaphase into unfertilized ooplasts. The donor four-cell-stage nuclei were synchronized in metaphase with nocodazole. The oocytes receiving a four-cell nucleus at metaphase formed two nuclei after artificial activation and inhibition of cytokinesis with cytochalasin B. To obtain embryos with diploid sets of chromosomes, nuclei from each reconstructed embryo were transferred individually into separate enucleated fertilized one-cell embryos, thus doubling the number of identical embryos. This procedure produced a high frequency of development of reconstructed embryos to the blastocyst stage. Of 11 sets of identical embryos produced by serial nuclear transplantation, 83% developed into blastocysts, including three sets of identical septuplet blastocysts. After transfer to recipient mice, a total of 25 (57%) live young were obtained, which included one set of identical sextuplet and two sets of identical quadruplet mice.
Unfertilized metaphase II oocytes have the potential to induce reprogramming of nuclei from cleavage-stage embryos, making them fully totipotent. To date, full-term development of embryos derived from the transplantation of nuclei from preimplantation-stage embryos has been reported in several mammalian species, including mice (1, 2), rabbits (3), cattle (4, 5), pigs (6), and sheep (7). However, in mice, reprogramming of embryonic nuclei appears restricted after embryonic genome activation (1, 2, 8), which occurs at the two-cell stage (9), because no live young have been produced from the transfer of embryonic nuclei beyond the two-cell stage (1, 8). Thus, to date, identical twin mice have not been produced by nuclear transfer to enucleated oocytes (ooplasts). Whether this failure is due to a lack of cytoplasmic factors required for reprogramming cleavage-stage nuclei in ooplasts or simply to technical difficulties is unknown.
Previously, we examined whether G2-phase nuclei undergo premature chromosome condensation after transfer to enucleated metaphase II oocytes (1, 2). This process is regulated by the cytoplasmic maturation promoting factor, a complex of cyclin and p34cdc2 (10). After artificial activation, normal diploid nuclei were formed following karyokinesis. Oocytes that received nuclei from late two-cell-stage embryos developed to term, but the rate of success was low because of the high frequency of chromosomal aberrations seen in the reconstituted eggs (11). This finding indicates that oocytes receiving a G2-stage nucleus do not develop a normal metaphase spindle. Recently, we improved the development of such reconstituted eggs to the blastocysts stage by transferring the nuclei from the donor embryos at the metaphase stage (O.Y.K. and T.K., unpublished data). Here, we report on the production of genetically identical mice by the transfer of metaphase-stage nuclei from four-cell mouse embryos. Nuclei from a single four-cell embryo produced six identical offspring.
MATERIALS AND METHODS
Oocytes and Embryos.
The oocyte and embryo donors were female B6CBF1 mice (C57BL/6j × CBA). They were superovulated with injections of 5 units of equine chorionic gonadotrophin (Peamex, Sankyo Ltd., Tokyo) and 5 iu of human chorionic gonadotrophin (Puberogen, Sankyo Ltd., Tokyo) given 48 h apart. Oocytes at metaphase II were released from the oviducts 14 h after injection of human chorionic gonadotrophin, and the cumulus cells were removed by digestion with 300 units/ml hyaluronidase in M2 medium (12). Late four-cell-stage embryos were flushed from the oviducts of superovulated females that had been mated with males of the same strain. The donor embryos were cultured in M16 medium (13) containing 1 μg/ml nocodazole, a microtubule polymerization inhibitor, for 4–6 h to induce metaphase arrest.
Nuclear Transplantation.
Identical embryos were reconstructed by serial nuclear transfer, as shown in Fig. 1. Nuclear transfer procedures are described elsewhere (1). All micromanipulations were performed in M2 medium containing 5 μg/ml cytochalasin B and 1 μg/ml nocodazole in a micromanipulation chamber. After enucleation of metaphase II chromosomes (14), a karyoplast containing metaphase chromosomes from the four-cell donor embryos was introduced with inactivated Sendai virus (hemagglutinating virus of Japan) 2700 hemagglutinating activity units per milliliter into the perivitelline space of the enucleated oocytes. After 4 h of culture in CZB medium (15, 16), the manipulated oocytes were artificially activated by electric stimulation (1.5 kV/cm for 100 μs × 6) (17, 18), followed by two further DC pulses of 1.5 kV/cm for 50 μs at 20 and 40 min after the initial stimulation. These multiple stimulations improve both the rate of activation and development to the blastocyst stage (19). The reconstituted eggs were cultured in CZB medium containing 5 μg/ml cytochalasin B to inhibit an extrapolar body extrusion and induce the formation of two “pronuclei-like nuclei.” Each nucleus was transferred into previously enucleated fertilized one-cell embryos obtained from mated superovulated females. This procedure allowed the reconstituted embryos to develop into blastocysts at high frequency (O.Y.K. and T.K., unpublished data).
Figure 1.
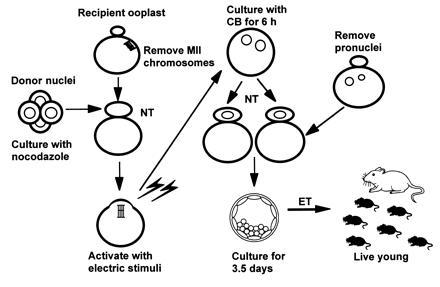
Construction of identical mouse embryos by nuclear transfer.
Culture and Embryo Transfer.
Each set of identical nuclear transplant embryos was cultured in CZB medium (15) to the late four-cell stage, then in modified CZB medium containing 27 mM glucose (16) in an atmosphere of 5% CO2, 5% O2, and 90% N2 at 37°C. Identical blastocysts derived from single donor embryos were transferred to the uterine horns of females on day 3 of pseudopregnancy (2.5 days post coitum).
RESULTS
Four-cell-stage donor embryos were synchronized at metaphase with nocodazole, fused individually with ooplasts, and artificially activated by electrical stimulation. The rates of fusion and activation were relatively constant (97%, 43/44 and 88%, 38/43, respectively). Both diploid nuclei were retained in the embryo after the completion of mitosis by incubation in cytochalasin B to prevent cytokinesis. To produce embryos with a diploid chromosome constitution, each nucleus was transferred into an enucleated fertilized one-cell embryo, thus doubling the number of identical embryos. This procedure produced a high frequency of development of reconstructed embryos to the blastocyst stage in vitro compared with that of embryos reconstituted in an artificially activated and enucleated one-cell parthenogenetic embryo (83%, 58/70 versus 16%, 8/49). The majority of embryos reconstituted with parthenogenetic cytoplasts arrested at the two-cell stage.
Eleven sets of identical embryos were produced by serial nuclear transplantation (Fig. 1), and four to eight identical one-cell embryos were obtained from a single four-cell embryo (Table 1). After 4 days of culture, 83% of 70 reconstructed embryos developed to blastocysts, which included three sets of seven identical blastocysts (Table 1 and Fig. 2a). To examine totipotency, 11 sets of identical blastocysts were transferred to recipient females on day 3 of pseudopregnancy, and the eight recipients became pregnant (Table 2). A total of 25 (57%) live young were obtained, which included one set of sextuplet (Fig. 2b) and two sets of quadruplet mice. The sex ratio of clones was biased toward males; only one set of triplets was female, whereas the other seven sets were males. To examine fertility of the identical sextuplet males, they were mated with CD-1 females. All females became pregnant and produced 8–14 live young, showing that the identical sextuplet mice possess normal reproductive ability.
Table 1.
In vitro development of identical mouse eggs reconstructed with four-cell nuclei synchronized at metaphase
| Identical sets | No. of eggs
examined
|
No. of reconstructed eggs developed to
|
|||||
|---|---|---|---|---|---|---|---|
| 1st NT* | 2nd NT† | Two-cell | Four-cell | Eight-cell | Morula | Blastocyst | |
| i | 4 | 8 | 8 | 7 | 7 | 7 | 7 |
| ii | 4 | 8 | 8 | 7 | 7 | 7 | 7 |
| iii | 4 | 8 | 8 | 6 | 5 | 5 | 5 |
| iv | 4 | 7 | 7 | 7 | 6 | 6 | 3 |
| v | 4 | 7 | 7 | 7 | 7 | 7 | 7 |
| vi | 4 | 7 | 7 | 6 | 6 | 6 | 6 |
| vii | 4 | 6 | 6 | 6 | 5 | 5 | 5 |
| viii | 4 | 6 | 6 | 6 | 6 | 6 | 6 |
| ix | 4 | 5 | 5 | 5 | 5 | 5 | 5 |
| x | 4 | 4 | 4 | 3 | 3 | 3 | 3 |
| xi | 3 | 4 | 4 | 4 | 4 | 4 | 4 |
| Total | 43 | 70 | 70 | 64 | 61 | 61 | 58 |
| (%) | (100) | (91) | (87) | (87) | (83) | ||
NT, Nuclear transfer.
Nuclear transferred eggs formed two pronuclei.
After the first NT, each nuclei was transferred into enucleated fertilized zygotes obtained from superovulated mated females.
Figure 2.
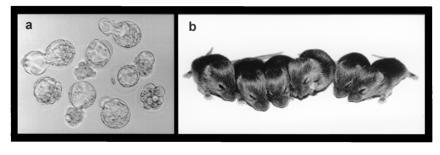
Sets of identical septuplet blastocysts (a) and identical sextuplet mice (b) obtained from a four-cell embryo.
Table 2.
In vivo development of identical embryos after transfer to recipients
| Identical sets | No. of blastocysts transferred | No. pregnants/no. of recipients | No. of live young |
|---|---|---|---|
| i | 7 | + | 6 ♂ |
| ii | 7 | + | 4 ♂ |
| iii | 5 | − | — |
| iv | 3 | + | 1 ♂ |
| v | 7 | + | 3 ♀ |
| vi | 6 | − | — |
| vii | 5 | + | 1 ♂ |
| viii | 6 | + | 3 ♂ |
| ix | 5 | + | 3 ♂ |
| x | 3 | − | — |
| xi | 4 | + | 4 ♂ |
| Total (%) | 58 | 8 (73) | 25 (57) |
Three to seven control blastocyts obtained from an albino mouse were transferred with identical embryos.
DISCUSSION
We have shown that live young can be produced from metaphase nuclei derived from four-cell-stage mouse embryos. Moreover, when the nuclei obtained after completion of karyokinesis in the activated ooplast are transferred individually to enucleated fertilized one-cell embryos, eight identical embryos were reconstituted from a single four-cell-stage embryo (see Fig. 1). By means of this serial nuclear transfer technique, we obtained the first identical sextuplet mice, which were fertile. The results demonstrate that all the nuclei derived from the original four-cell embryo are totipotent when serially passaged through cytoplasts of unfertilized oocytes and fertilized one-cell embryos. Most of the live young obtained were males; only one set of identical female triplet pups were female. This may be due to the selection of the faster developing four-cell embryos for nuclear transfer because it has been shown previously in mice that the embryos developing faster during preimplantation tend to have an XY sex chromosome constitution (20).
Many attempts have been made to clarify cell cycle effects of donor nuclei on the developmental ability of reconstructed embryos (2, 3, 7, 21–24). It is thought that donor nuclei at G1 or G1/S phase are at a suitable stage for embryo cloning; therefore, attempts to control the cell cycle of embryonic nuclei have used chemicals with low toxicity such as aphidicolin, as well as DNA polymerase I inhibitor. In rabbits, experiments on nuclear transfer of G1-phase and late S-phase nuclei synchronized with colcemid and aphidicolin showed that G1-phase nuclei have a greater potential to contribute development of the reconstituted embryos (25). Recently, Cheong et al. (26) have reported that transferring nuclei at G1 phase from two-, four-, and eight-cell mouse embryos into enucleated oocytes resulted in the birth of live offspring. However, we failed to confirm their results, even though we repeated the experiments precisely. The reason for this is not clear, but it is extremely difficult to obtain embryonic nuclei at G1-phase because it is so transitory (unpublished data). The early cell cycles of mouse embryos are easily synchronized at metaphase by culturing the embryos with nocodazole, which inhibits tubulin polymerization. Exposure of embryos to the concentration of nocodazole necessary to cause cell cycle arrest does not affect the ability of the manipulated embryos to complete development to term (ref. 27 and unpublished data). Thus, large numbers of identical embryos can be produced from the donor nuclei.
Development to the blastocyst stage was improved significantly (16% to 83%) when pronuclei-like nuclei from the reconstructed one-cell embryos were transferred into enucleated fertilized rather than parthenogenetic one-cell embryos. This shows that the ability of cytoplasm to support development differs markedly between fertilized and parthenogenetic one-cell embryos. The reason is unknown. One possibility is that new transcripts from the male genome may support further development of the reconstituted embryos, because it has been reported that zygotic genome activation (28) and exogenous gene expression (29) occur in the late one-cell mouse embryos. Therefore, when fertilized early one-cell-stage embryos that were expected to be beginning transcription from the male genome were used in the second nuclear transfer, development of the reconstituted embryos was not improved (data not shown). Alternatively, nucleocytoplasmic interaction (30) in hybrid embryos may be responsible for this phenomenon. The reconstitution of embryos with fertilized nuclei and parthenogenetic cytoplasm may have a detrimental effect on development.
In domestic animals, live young have been produced from ICM cells and a cultured embryonic cell line in cattle (31) and sheep (32). Identical clones have also been produced in both species, the maximum number reported being 10 in cattle (4). A significant proportion of cattle and sheep embryos produced by nuclear transfer give rise to larger than normal offspring at birth (33, 34). So far, it has not been observed in offspring of mice and pigs produced by nuclear transfer. How this phenomenon arises is not known, but one possibility is that epigenetic modifications occur in imprinted or unimprinted genes that lead to changes in gene expression with subsequent effects on fetal growth.
Our results show clearly that metaphase-stage nuclei from four-cell mouse embryos can be reprogrammed by the present serial nuclear transfer system, and the reconstructed embryos have the capacity to complete embryonic development to term. Nevertheless, how and when donor metaphase nuclei are reprogrammed and acquire totipotency is not known. Perhaps reprogramming occurs mainly during decondensation of nuclei in reconstructed oocytes after parthenogenetic activation. Also, it is possible that additional reprogramming occurs in the first mitotic cycle after the second nuclear transfer, because developmental ability was significantly improved by the transfer of reconstituted pronuclei-like nuclei to enucleated zygotes. The present manipulative techniques suggest that larger scale cloning by nuclear transfer is possible in mice and other species.
Acknowledgments
We thank Prof. D. G. Whittingham for critically reading the manuscript, and Dr. T. Nakahara and Prof. Y. Tsunoda for helpful discussions.
References
- 1.Kono T, Kwon O Y, Nakahara T. J Reprod Fertil. 1991;93:165–172. doi: 10.1530/jrf.0.0930165. [DOI] [PubMed] [Google Scholar]
- 2.Kono T, Kwon O Y, Watanabe T, Nakahara T. J Reprod Fertil. 1992;94:481–487. doi: 10.1530/jrf.0.0940481. [DOI] [PubMed] [Google Scholar]
- 3.Collas P, Robl J M. Biol Reprod. 1990;43:877–884. doi: 10.1095/biolreprod43.5.877. [DOI] [PubMed] [Google Scholar]
- 4.Stice S L, Keefer C L. Biol Reprod. 1993;48:715–719. doi: 10.1095/biolreprod48.4.715. [DOI] [PubMed] [Google Scholar]
- 5.Kono T, Sotomaru Y, Aono F, Takahasi T, Ogiwara I, Sekizawa F, Arai T, Nakahara T. Theriogenology. 1994;41:1463–1471. doi: 10.1016/0093-691x(94)90197-q. [DOI] [PubMed] [Google Scholar]
- 6.Prather R S, Sims M M, First N L. Biol Reprod. 1989;41:414–418. doi: 10.1095/biolreprod41.3.414. [DOI] [PubMed] [Google Scholar]
- 7.Campbell K H S, Loi P, Cappai P, Wilmut I. Biol Reprod. 1994;50:1385–1393. doi: 10.1095/biolreprod50.6.1385. [DOI] [PubMed] [Google Scholar]
- 8.Tsunoda Y, Tokunaga T, Imai H, Uchida T. Development (Cambridge, UK) 1989;107:407–412. doi: 10.1242/dev.107.2.407. [DOI] [PubMed] [Google Scholar]
- 9.Flach G, Johnson M H, Braude P R, Taylor R A S, Bolton V N. EMBO J. 1982;1:681–686. doi: 10.1002/j.1460-2075.1982.tb01230.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kubiak J Z, Weber M, de Pennart H, Winston N J, Maro B. EMBO J. 1993;12:3773–3778. doi: 10.1002/j.1460-2075.1993.tb06055.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Kono T, Ogawa M, Nakahara T. Jpn J Reprod Dev. 1993;39:301–307. [Google Scholar]
- 12.Quinn P, Barros C, Whittingham D G. J Reprod Fertil. 1982;66:161–168. doi: 10.1530/jrf.0.0660161. [DOI] [PubMed] [Google Scholar]
- 13.Whittingham D G. J Reprod Fertil. 1971;14:7–12. [PubMed] [Google Scholar]
- 14.Kono T, Sotomaru Y, Sato Y, Nakahara T. Mol Reprod Dev. 1993;34:43–46. doi: 10.1002/mrd.1080340107. [DOI] [PubMed] [Google Scholar]
- 15.Chatot C, Lewis J, Torres I, Ziomek C. Biol Reprod. 1990;42:432–440. doi: 10.1095/biolreprod42.3.432. [DOI] [PubMed] [Google Scholar]
- 16.Chatot C, Williams J, Torres I, Ziomek C. Mol Reprod Dev. 1994;37:407–412. doi: 10.1002/mrd.1080370407. [DOI] [PubMed] [Google Scholar]
- 17.Stice S, Robl J M. Biol Reprod. 1988;39:657–664. doi: 10.1095/biolreprod39.3.657. [DOI] [PubMed] [Google Scholar]
- 18.Onodera M, Tsunoda Y. Gamete Res. 1989;22:277–283. doi: 10.1002/mrd.1120220305. [DOI] [PubMed] [Google Scholar]
- 19.Collas P, Fissore R, Robl J M, Sullivan E J, Barnes F L. Mol Reprod Dev. 1993;34:212–223. doi: 10.1002/mrd.1080340214. [DOI] [PubMed] [Google Scholar]
- 20.Tsunoda Y, Tokunaga T, Sugie T. Gamete Res. 1985;12:301–304. [Google Scholar]
- 21.Campbell K H S, Ritchie W A, Wilmut I. Biol Reprod. 1993;49:933–942. doi: 10.1095/biolreprod49.5.933. [DOI] [PubMed] [Google Scholar]
- 22.Pinto-Correia C, Collas P, Leon F A P D, Robl J M, De-Leon F A P. Mol Reprod Dev. 1993;34:33–42. doi: 10.1002/mrd.1080340106. [DOI] [PubMed] [Google Scholar]
- 23.Barnes F L, Collas P, Powell R, King W A, Westhusin M, Shepherd D. Mol Reprod Dev. 1993;36:33–41. doi: 10.1002/mrd.1080360106. [DOI] [PubMed] [Google Scholar]
- 24.Collas P, Balise J, Robl J M. Biol Reprod. 1992;46:492–500. doi: 10.1095/biolreprod46.3.492. [DOI] [PubMed] [Google Scholar]
- 25.Collas P, Robl J M. Biol Reprod. 1991;45:455–465. doi: 10.1095/biolreprod45.3.455. [DOI] [PubMed] [Google Scholar]
- 26.Cheong H T, Takahashi Y, Kanakawa H. Biol Reprod. 1993;48:958–963. doi: 10.1095/biolreprod48.5.958. [DOI] [PubMed] [Google Scholar]
- 27.Kato Y, Tsunoda Y. J Reprod Fertil. 1992;95:39–43. doi: 10.1530/jrf.0.0950039. [DOI] [PubMed] [Google Scholar]
- 28.Latham K, Solter D, Schultz R M. Dev Biol. 1992;149:457–462. doi: 10.1016/0012-1606(92)90300-6. [DOI] [PubMed] [Google Scholar]
- 29.Ram P, Schultz R M. Dev Biol. 1993;156:552–556. doi: 10.1006/dbio.1993.1101. [DOI] [PubMed] [Google Scholar]
- 30.Reik W, Romer I, Barton S C, Surani M A, Howlett S K, Klose J. Development (Cambridge, UK) 1993;119:933–942. doi: 10.1242/dev.119.3.933. [DOI] [PubMed] [Google Scholar]
- 31.Sims M, First L. Proc Natl Acad Sci USA. 1994;91:6143–6147. doi: 10.1073/pnas.91.13.6143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Campbell K H S, McWhir J, Ritchie W A, Wilmut I. Nature (London) 1996;380:64–66. doi: 10.1038/380064a0. [DOI] [PubMed] [Google Scholar]
- 33.Wilson J M, Williams J D, Bondioli K R, Looney C R, Westhusin M E, McCalla D F. Anim Reprod Sci. 1995;38:73–83. [Google Scholar]
- 34.Walker S K, Hartwich K M, Seamark R F. Theriogenology. 1996;45:111–120. [Google Scholar]