

Abstract
Vesicular stomatitis New Jersey virus (VSV-NJ) is a rhabdovirus that causes economically important disease in cattle and other domestic animals in endemic areas from southeastern United States to northern South America. Its negatively stranded RNA genome is capable of undergoing rapid evolution, which allows phylogenetic analysis and molecular epidemiology studies to be performed. Previous epidemiological studies in Costa Rica showed the existence of at least two distinct ecological zones of high VSV-NJ activity, one located in the highlands (premontane tropical moist forest) and the other in the lowlands (tropical dry forest). We wanted to test the hypothesis that the viruses circulating in these ecological zones were genetically distinct. For this purpose, we sequenced the hypervariable region of the phosphoprotein gene for 50 VSV-NJ isolates from these areas. Phylogenetic analysis showed that viruses from each ecological zone had distinct genotypes. These genotypes were maintained in each area for periods of up to 8 years. This evolutionary pattern of VSV-NJ suggests an adaptation to ecological factors that could exert selective pressure on the virus. As previous data indicated an absence of virus adaptation to factors related to the bovine host (including immunological pressure), it appears that VSV genetic divergence represents positive selection to adapt to specific vectors and/or reservoirs at each ecological zone.
Keywords: RNA virus, positive non-immune selection, ecological distribution, genetic variability
Vesicular stomatitis virus (VSV) is an insect-transmitted member of the vesiculovirus genus in the Rhabdoviridae family. The virus RNA genome is negative sense, single-stranded, and approximately 11 kb in length. VSV is capable of undergoing rapid evolution and therefore has served as model for numerous viral RNA evolution studies (1–5). In addition, VSV is an important cause of clinical disease in farm animals throughout the Americas. According to the interval between clinical disease occurrence, there are areas of epidemic (5- to 10-year intervals) and endemic (<1 year) VSV activity (6). Endemic activity is most common from northern South America to northern Mexico, whereas epidemic activity occurs mostly in southern South America, the United States, and Canada (6, 7). Extensive epidemiological studies in Costa Rica have identified two VSV New Jersey (VSV-NJ) endemic foci in that country: one located in the dry lowlands (tropical dry forest) and the other in the highlands (premontane/montane moist forest) (Table 1) (11). Typically, over 80–90% of the cattle living in these areas have antibodies to VSV-NJ, and ≈9% suffer clinical vesicular disease in a given year (11, 13, 14). Antibodies to VSV-NJ are also found in wildlife (rodents and primates) captured in these areas (ref. 12; L.L.R., unpublished data). In contrast, in wet lowlands (tropical moist/wet forest) and wet highlands at altitudes over 2500 m (lower montane and montane rain forests) VSV-NJ either does not occur or occurs at longer (4–5 years) intervals (Table 1).
Table 1.
Summary of epidemiological data supporting the existence of two distinct VSV-NJ endemic zones (one located in the highlands and the other in the lowlands)
| Area* | Seroprevalence in cattle† | Interepidemic interval‡ | Seroprevalence in wildlife† |
|---|---|---|---|
| Highlands moist | High | <12 months | High |
| Lowlands dry | High | <12 months | High |
| Lowlands wet | Low | >12 months | No data |
| available | |||
| Highlands wet | Low | >12 months | No data |
| available |
Ecological areas are classified according to average annual rainfall as dry, moist, wet, and rain forest (8, 9). Areas with intermediate rainfall values are classified as transition areas [e.g., moist (dry) = dry forest, moist transition]. Highlands moist includes three ecological zones: premontane wet forest, premontane moist forest, and lower montane wet forest. Lowlands dry includes two ecological zones: tropical moist forest (dry transition) and Tropical dry forest. Lowlands wet includes two ecological zones: tropical moist forest (wet transition) and tropical wet forest. Highlands wet includes three ecological zones: premontane rain forest, lower montane rain forest, and montane rain forest. There are no highland dry areas.
Indicates the prevalence of neutralizing antibodies to VSV-NJ (refs. 11 and 12; L.L.R., unpublished data).
Time lapse between outbreaks. Data obtained from the archives of the Ministry of Agriculture of Costa Rica and United States Department of Agriculture Foot and Mouth Prevention Program (1986–1993). Also see Pérez and Cornelissen (10).
Previous genetic analysis of several complete genes of numerous VSV natural isolates had identified a hypervariable region in the phosphoprotein gene that appeared to represent a spacer domain similar to that seen in other viral transcriptional activators (15). Comparison of evolutionary trees generated using complete gene sequences or a 450-nt region encompassing the phosphoprotein hypervariable region indicated that this fragment was highly informative and could be reliably used to predict the phylogenetic relationship of a large number of VSV isolates (2, 15–17). Using this approach, previous phylogenetic analysis of VSV-NJ isolates from throughout North and Central America had suggested an evolutionary pattern inconsistent with neutral evolution (17). Analysis of resulting evolutionary trees indicated that from the ancestral node to the branch tips, there was a geographic rather than temporal relationship among the virus samples, and no molecular clock was discernible.
In the study presented here, we analyzed the phylogenetic relationship of 50 VSV strains originating from different ecological zones of Costa Rica. The phylogenetic relationship among these viruses and those from the surrounding Central American countries showed that two distinct lineages of VSV-NJ existed and correlated with different ecological zones, suggesting that ecological factors (i.e., selective forces) play an important role in molding the natural evolution of VSV.
MATERIALS AND METHODS
Virus Isolates.
Samples from throughout North and Central America have been described elsewhere (2, 17). Costa Rican samples collected between 1982 to 1990 originated from all seven provinces of Costa Rica and represented all ecological zones where VSV is known to occur (Table 2). All viruses were identified by virus neutralization assay with VSV-NJ and VSV Indiana (VSV-IN) specific reference antisera (kindly supplied by J. Pearson, National Veterinary Services Laboratory, U.S. Department of Agriculture, Ames, IA). Ribonucleic acid was extracted either directly from epithelium and vesicular fluid or from tissue culture cells infected with low passage virus stocks, using the guanidine isothiocyanate-acid phenol procedure as previously described (18).
Table 2.
Distribution of VSV-NJ specimens by geographic and ecological zones
| Specimen* | Location† | Altitude, m | Ecological zone‡ |
|---|---|---|---|
| 7/87NCB2 | Managua, Nicaragua | 0–100 | ND |
| 10/82CRB | San Rafael, Guatuso | 0–100 | Tropical wet forest |
| 10/85HDB | Comayagua, Honduras | 500–600 | ND |
| 10/85HDB2 | Comayagua, Honduras | 500–600 | ND |
| ../87HDB3 | Comayagua, Honduras | 500–600 | ND |
| 12/82HDB | Fco Morazan, Honduras | 800–1000 | ND |
| ../76ECM | Guayas, Ecuador | 0–100 | ND |
| 11/83NCB | Matagalpa, Nicaragua | 600–800 | ND |
| 1/85NCB | Managua, Nicaragua | 500–600 | ND |
| 2/85NCB | Rivas, Nicaragua | 0–100 | ND |
| 7/83NCB | Managua, Nicaragua | 0–100 | ND |
| 7/83NCP | Managua, Nicaragua | 0–100 | ND |
| 8/86NCB | Managua, Nicaragua | 0–100 | ND |
| 7/85NCB2 | Managua, Nicaragua | 0–100 | ND |
| 7/85NCB | Managua, Nicaragua | 0–100 | ND |
| 7/86NCB | Managua, Nicaragua | 0–100 | ND |
| 10/82NCB | Managua, Nicaragua | 0–100 | ND |
| 10/89CRB2 | Zarcero, Alfaro Ruiz | 1500–2000 | Montane wet forest |
| 6/89CRB | Fortuna, San Carlos | 100–500 | Premontane wet forest |
| ../60PNB | Aguadulce, Panama | 100–500 | Tropical moist (dry) forest |
| 10/88CRB | Sandoval, Limon | 0–100 | Tropical wet forest |
| 6/87CRB | Carmona, Nandayure | 0–100 | Tropical moist (dry) forest |
| 11/87CRB3 | Upala, Upala | 0–100 | Tropical moist (wet) forest |
| 8/86CRB | Zapotal, Nandayure | 100–500 | Tropical dry forest |
| 7/86CRB | Tronadora, Tilaran | 500–600 | Tropical moist (wet) forest |
| 10/90CRB | Quebrada Honda, Nicoya | 100–500 | Tropical dry forest |
| 9/88CRB2 | Cutris, San Carlos | 0–100 | Tropical moist (wet) forest |
| 9/88CRB3 | Cutris, San Carlos | 0–100 | Tropical moist (wet) forest |
| 9/88CRB4 | Cutris, San Carlos | 0–100 | Tropical moist (wet) forest |
| 9/88CRB9 | Mayorga, Liberia | 0–100 | Tropical dry forest |
| 11/88CRB | Cutris, San Carlos | 0–100 | Tropical moist (wet) forest |
| 11/88CRB2 | Cutris, San Carlos | 0–100 | Tropical moist (wet) forest |
| 9/88CRB | San Rafael, Guatuso | 0–100 | Tropical wet forest |
| 9/88CRB5 | La Tigra, San Carlos | 500 | Tropical wet forest |
| 9/88CRB6 | Amparo, Los Chiles | 0–100 | Tropical moist (wet) forest |
| 9/88CRB7 | Amparo, Los Chiles | 0–100 | Tropical moist (wet) forest |
| 11/87CRB | Zarcero, Alfaro Ruiz | 1500–2000 | Montane wet forest |
| 9/88CRB8 | Terranova, Los Chiles | 0–100 | Tropical moist (wet) forest |
| 12/83PNB | Cocle, Panama | 0–100 | Tropical moist (dry) forest |
| 9/86PNB | Chiriqui, Panama | 1500–2000 | Premontane moist forest |
| 1/85PNB | Chiriqui, Panama | 1500–2000 | Premontane moist forest |
| 1/85PNB2 | Chiriqui, Panama | 1500–2000 | Premontane moist forest |
| 7/85PNB | Chiriqui, Panama | 1500–2000 | Premontane moist forest |
| 10/83PNB | Chiriqui, Panama | 1500–2000 | Premontane moist forest |
| 7/89CRB | Canoas, Corredores | 0–100 | Tropical wet forest |
| 10/89CRB | Atenas, Atenas | 600–1000 | Premontane moist forest |
| 11/87CRB4 | Naranjo, Naranjo | 600–1000 | Premontane wet forest |
| 11/87CRB2 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 11/88CRB3 | San Rafael, Coronado | 2000–2500 | Lower montane rain forest |
| 11/87CRB5 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 1/88CRB2 | San Isidro, Alajuela | 1000–1500 | Premontane moist forest |
| 2/88CRB4 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 2/88CRB2 | Cachi, Paraiso | 1000–1500 | Premontane moist forest |
| 1/88CRB | San Isidro, Alajuela | 1000–1500 | Premontane moist forest |
| 12/82CRB2 | Sta. Cruz, Turrialba | 1500–2000 | Lower montane wet forest |
| 9/86CRB3 | Zarcero, Alfaro Ruiz | 2000–2500 | Montane wet forest |
| 1/85CRB | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 1/90CRB2 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 12/90CRB | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 9/90CRB | San Juan, Poas | 1000–1500 | Premontane moist forest |
| 7/85CRB | Naranjito, Aguirre | 1000–1500 | Premontane moist forest |
| 12/86CRB | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 12/89CRB | Tapezco, Alfaro | 2000–2500 | Montane wet forest |
| 1/90CRB | Santo Domingo, Sta. Barbara | 1500–2000 | Lower montane wet forest |
| 2/88CRB | Quesada, San Carlos | 600–1000 | Tropical/premontane wet forest |
| 6/87CRB2 | Quesada, San Carlos | 600–1000 | Tropical/premontane wet forest |
| 12/86CRB2 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 12/86CRB4 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 9/86CRB2 | Zarcero, Alfaro Ruiz | 2000–2500 | Montane wet forest |
| 8/88CRB | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 12/86CRB3 | San Roque, Grecia | 1000–1500 | Premontane moist forest |
| 1/85PNB3 | Chiriqui, Panama | 1500–2000 | Premontane moist forest |
| 12/84CRB | Sta. Lucia, Barva | 1000–1500 | Premontane moist forest |
Samples are shown on the same order as they appear in Fig. 2. Numbers indicate month, followed by year of clinical case. CR, Costa Rica; HD, Honduras; NC, Nicaragua; PN, Panama; B, bovine. Last number distinguishes different specimens from the same month and place. .., Unknown month.
District and county are indicated for Costa Rican samples; province and country are indicated for other samples.
Reverse Transcription, DNA Amplification, and Sequence Analysis.
Viral specific sequences were amplified using the above RNA preparations as templates in a single tube reaction using reverse transcriptase and Taq polymerase as described previously (18). The single tube reaction simplified the procedure for routine purposes and minimized the opportunity for cross-contamination between samples. Primers used for both reverse transcriptase and polymerase chain reaction (PCR) reactions were specific for conserved sites in the phosphoprotein gene of VSV-NJ, resulting in amplification of a 642-bp fragment containing the hypervariable region. The nucleotide sequence of a 450-bp region of the resulting PCR products was determined by the primer extension dideoxy chain termination method as described previously (17, 19). Sequences of the VSV-NJ phosphoprotein gene fragment were aligned with those of VSV-NJ, VSV-IN, and Chandipura virus obtained previously (17, 20). Phylogenetic analysis by maximum parsimony was done using the ancstr program by Fitch (21).
Ecological Zones.
Costa Rica is a relatively small country (50,000 km2) bordered by Nicaragua (to the north) and Panama (to the south). Despite its small size, a wide diversity of ecological zones and microclimates are found among the major mountain ranges (9). Abrupt changes in altitude over short distances account for the close proximity of dramatically different ecological zones. Based on altitude, rainfall, temperature, and evapotranspiration potential, the country has been classified into 19 ecological zones (8, 9).
RESULTS AND DISCUSSION
A maximum parsimony analysis of over 180 VSV-NJ isolates based on nucleotide sequence differences in the phosphoprotein gene hypervariable region was carried out. The analysis was rooted using the nucleotide sequences of the equivalent genome region of 10 diverse VSV-IN viruses (15) and the more distantly related vesiculovirus, Chandipura (20). The overall topology of this tree was essentially identical to that generated previously using a smaller virus sample set (17), with the exception of the ancestral node of the VSV-NJ samples now being placed between North and Central American samples (Fig. 1). The root was shown in the wrong place in the earlier presentation.
Figure 1.

Summary of the evolutionary tree topology for VSV-NJ from North and Central America. Horizontal distances are proportional to the number of nucleotide steps between viruses or branch points. Vertical distances are for graphic display only. Individual branches represents multiple virus sequences. A, B, and asterisk refer to details shown in Fig. 2.
As seen previously, the phylogenetic relationship of these viruses correlated more clearly with geographic location rather than year of isolation. No molecular clock was evident, and discontinuities (i.e., long branch lengths) were seen suggestive of punctuated equilibrium. A particularly long branch is seen separating Costa Rican and Panamanian samples from Costa Rican and Nicaraguan samples (Fig. 1). A detailed picture of this section of the tree in shown in Fig. 2. The short branch lengths (i.e., genetic stasis) observed within these two lineages contrasts the long branch separating them. Analysis of the virus samples relative to the ecological zone from which they originated revealed a rather striking correlation with these two observed lineages (Fig. 2). Almost all the isolates originating in the lowlands were located in lineage A, whereas almost all the samples from the highlands were located in lineage B. Only seven exceptions to this distribution were seen among 75 viruses in this region of the tree. Inadvertent transportation of infected animals among different regions seems a likely explanation for these few exceptions. That this relationship is ecological as opposed to purely geographic can be seen from a more detailed examination of the samples. For instance, isolates such as 1/85PNB3 (from Renacimiento, Chiriqui, Panama) and 12/84CRB (from Barva, Heredia, Costa Rica), having identical nucleotide sequence, are actually over 800 km apart, but originate from the same contiguous highland ecological zone in the Talamanca mountain range (Fig. 2, indicated by ∗). In contrast, samples such as 2/88CRB and 9/88CRB5, which originated only 25 km apart but are from different ecological zones, can differ by as much as 44 nucleotides (Fig. 2, indicated by ♠). These data are consistent with VSV natural evolution predominantly resulting from virus selection by ecological factors, rather than simply neutral drift in geographically separated areas.
Figure 2.
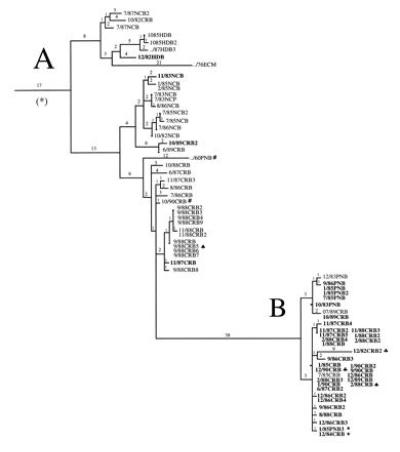
Detail of the tree branches corresponding to VSV-NJ sequences from Honduras, Nicaragua, Costa Rica, and Panama that are marked with an asterisk in Fig. 1. Number of nucleotide substitutions between viruses or branch points are indicated above branch lines. Virus nomenclature indicates month, year of clinical case, location, and host from which sample was obtained; the last number distinguishes samples from the same time and place. HD, Honduras; NC, Nicaragua; CR, Costa Rica; PN, Panama; B, bovine; M, mosquito; P, porcine. Branch A contains mostly viruses (plain font) originating from lowland (<600 m) areas. Branch B contains mostly viruses (boldface type) from highland areas (>600 m). See Table 2 for detailed geographical origin of each viral sequence.
The lack of a molecular clock or the genetic stability at each ecological zone is illustrated by two examples. In lineage A, only 16 nucleotide differences accumulated between samples obtained 30 years apart (../60PNB and 10/90CRB; Fig. 2, indicated by #) and in lineage B only 10 nucleotide differences are observed between samples obtained 8 years apart (12/82CRB2 and 12/90CRB; Fig. 2, indicated by ♣). This genetic stability is observed despite the presence of high titers of neutralizing antibodies in the cattle population from which they originated (13). Previous data showed that immunological pressure did not play a role in the evolution of VSV in a highland endemic zone of Costa Rica (22). In that prospective study, serum samples obtained before and after clinical vesicular stomatitis in cattle were equally capable of neutralizing the disease-causing VSV strain. Furthermore, sequence analysis of the region in the glycoprotein gene that encodes the major antibody neutralizing epitopes of these viruses showed no meaningful changes over a period of 4 years under the immunological pressure of a cattle population with a prevalence of neutralizing antibodies of over 90% (22). This evolutionary pattern contrasts with that of some other negative-stranded RNA viruses, e.g., human influenza A virus, in which a reliable molecular clock and an immune selection driven evolutionary pattern are observed (23–25).
It could be argued that the genetic divergence observed in VSV-NJ is an adaptation to the cattle host, since most cattle in the Costa Rican highlands are European breeds (Bos taurus, e.g., Holstein, Jersey), whereas most cattle residing in the lowlands are Indian breeds (Bos indicus, e.g., Brahman, Cebu). However, an epidemiological study determined that cattle residing in the defined highland and lowland endemic foci were at up to 10 times higher risk of being seropositive to VSV-NJ than cattle residing elsewhere, regardless of breed, sex, and other host-associated factors (11).
What ecological factors might then influence the natural evolution of VSV-NJ? Vesicular stomatitis is an insect-transmitted virus that has been isolated from several families of mosquitoes and flies (26). Sand flies (e.g., Lutzomyia shannoni) and black flies (Simuliidae spp.) have been shown to transmit VSV both transovarially (i.e., vertically) and to susceptible hosts (27–31). It is not known which are the natural insect vectors of VSV in endemic areas of Costa Rica. However, preliminary studies have shown differences in the species distribution of sand flies in highland and lowland endemic areas (32). While sand flies, including L. shannoni, are abundant in the lowland endemic areas, they are less common in the highlands. However, black flies (Simulium spp.) are found in greater numbers in the highlands than they are in the lowland endemic zones (33). It is possible that the genetic divergence observed in VSV-NJ represents adaptation to a different vector at each ecological zone. Recent analysis of VSV fitness in insect and mammalian cells has shown that selection for fitness in different host cell environments can represent a potent force molding VSV evolution (34).
Antibodies to VSV have been detected in monkeys, marsupials, bats, carnivores, deer, and rodents throughout the Americas (35–38). The absence of sustained viremias in domestic hosts, such as cattle, pigs, and horses, has led to the suggestion of a wildlife reservoir capable of sustained viremias to feed the natural cycle of VSV in insects (35, 39). However, to this day, the putative reservoir remains elusive. Preliminary wildlife serological surveys have shown that of eight small mammal species tested in the highland endemic zone, only Sigmodon hispidus (cotton rat) had VSV specific antibodies (12). In contrast, in the lowland endemic zone, four species of rodents (Liomys salviini, Ototylomus phylotis, Oryzomys spp., and S. hispidus) and one species of howler monkey (Alloata palliata) have VSV antibodies (L.L.R., unpublished data). It is then possible that the divergent VSV-NJ genetic lineages, represent adaptations to different mammalian reservoirs at each ecological zone.
In conclusion, these data are consistent with the view that the extent and frequency of alteration of selective environments and ensuing distortions of virus fitness landscapes, profoundly influence the resulting virus phylogenetic structure (40). The practical importance of understanding these processes is highlighted by the recent emergence of several viral diseases (41), including the 1995 reemergence of epidemic VSV-NJ in the United States after a 10-year hiatus.
Acknowledgments
We thank Joan Rowe and Rocío Cortés for their valuable technical help, Christina Spiropoulou for her help in RNA extractions in Costa Rica, Helene Van for her help with the phylogenetic tree analysis, the personnel of the Ministry of Agriculture of Costa Rica, and farmers who provided clinical samples and epidemiological information. This work was made possible by grants from International Foundation for Science Project B/951-3 (L.L.R.), Swedish Agency of Research Cooperation (L.L.R.), The Rockefeller Foundation Biotechnology Career Fellowship Program (L.L.R.), U.S. Department of Agriculture Animal Molecular Biology Competitive Research Grant 90–37266-5473 (S.T.N.), and National Science Foundation Grant BRR 8796183 (W.M.F.).
Footnotes
Abbreviations: VSV, vesicular stomatitis virus; VSV-NJ, VSV New Jersey; VSV-IN, VSV Indiana.
Data deposition: The sequences reported in this paper have been deposited in the GenBank data base (accession nos. U39202–U39225U39202U39203U39204U39205U39206U39207U39208U39209U39210U39211U39212U39213U39214U39215U39216U39217U39218U39219U39220U39221U39222U39223U39224U39225).
References
- 1.Holland J J, De la Torre J C, Clarke D K, Duarte E. J Virol. 1994;65:2960–2967. doi: 10.1128/jvi.65.6.2960-2967.1991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bilsel P A, Nichol S T. J Virol. 1990;64:4873–4883. doi: 10.1128/jvi.64.10.4873-4883.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Clarke D K, Duarte E, Moya A, Elena S, Domingo E, Holland J. J Virol. 1993;67:222–228. doi: 10.1128/jvi.67.1.222-228.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Clarke D K, Duarte E, Elena S, Moya A, Domingo E, Holland J. Proc Natl Acad Sci USA. 1994;91:4821–4824. doi: 10.1073/pnas.91.11.4821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Duarte E, Clarke D K, Moya A, Elena S, Domingo E, Holland J. J Virol. 1993;67:3620–3623. doi: 10.1128/jvi.67.6.3620-3623.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hanson R. In: Virus Diseases of Food Animals. Gibbs E P, editor. Vol. 2. New York: Academic; 1981. pp. 517–539. [Google Scholar]
- 7.Nichol S T. In: Encyclopedia of Virology. Webster R G, Granoff A, editors. London: Saunders; 1994. pp. 1547–1555. [Google Scholar]
- 8.Holdridge L R. Ecological Map of Costa Rica. Costa Rica: National Geographical Institute and Tropical Science Center; 1969. [Google Scholar]
- 9.Holdridge L R. Ecology Based on Life Zones. Costa Rica: Inter-American Institute for Cooperation in Agriculture Press; 1978. [Google Scholar]
- 10.Pérez E, Cornelissen B. Prev Vet Med. 1988;6:1–8. [Google Scholar]
- 11.Atwill E R, Rodríguez L L, Hird D, Rojas O. Prev Vet Med. 1993;15:303–314. [Google Scholar]
- 12.Jiménez A E, Jiménez C, Castro L, Rodríguez L. J Wild Dis. 1996;32:274–279. doi: 10.7589/0090-3558-32.2.274. [DOI] [PubMed] [Google Scholar]
- 13.Rodríguez L L, Vernon S, Morales A I, Letchworth G J. Am J Trop Med Hyg. 1990;42:272–281. doi: 10.4269/ajtmh.1990.42.272. [DOI] [PubMed] [Google Scholar]
- 14.Vanleeuwen J A, Rodríguez L L, Waltner-Toews D. Am J Trop Med Hyg. 1995;53:342–350. doi: 10.4269/ajtmh.1995.53.342. [DOI] [PubMed] [Google Scholar]
- 15.Bilsel P A, Rowe J E, Fitch W M, Nichol S T. J Virol. 1990;64:2498–2504. doi: 10.1128/jvi.64.6.2498-2504.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Nichol S T, Rowe J E, Fitch W M. Virology. 1989;168:281–291. doi: 10.1016/0042-6822(89)90268-7. [DOI] [PubMed] [Google Scholar]
- 17.Nichol S T, Rowe J E, Fitch W M. Proc Natl Acad Sci USA. 1993;90:10424–10428. doi: 10.1073/pnas.90.22.10424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rodríguez L L, Letchworth G J, Spiropoulou C F, Nichol S T. J Clin Microbiol. 1993;31:2016–2020. doi: 10.1128/jcm.31.8.2016-2020.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Winship P R. Nucleic Acids Res. 1989;17:1266–1269. doi: 10.1093/nar/17.3.1266. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Masters P S, Banerjee A K. Virology. 1987;157:298–306. doi: 10.1016/0042-6822(87)90272-8. [DOI] [PubMed] [Google Scholar]
- 21.Fitch W M. Syst Zool. 1971;20:406–416. [Google Scholar]
- 22.Vernon SD, Rodríguez L, Letchworth G J. Virology. 1990;177:209–215. doi: 10.1016/0042-6822(90)90474-6. [DOI] [PubMed] [Google Scholar]
- 23.Fitch W M, Leiter J M E, Li X, Palese P. Proc Natl Acad Sci USA. 1991;88:4270–4274. doi: 10.1073/pnas.88.10.4270. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Webster R G, Bean W J, Gorman O T, Chambers T M, Kawaoka Y. Microbiol Rev. 1992;56:152–179. doi: 10.1128/mr.56.1.152-179.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Scholtissek C, Ludwig S, Fitch W M. Arch Virol. 1993;131:237–250. doi: 10.1007/BF01378629. [DOI] [PubMed] [Google Scholar]
- 26.Francy D B, Moore C G, Smith C, Jacob W L, Taylor S A, Calisher C H. J Med Entomol. 1988;25:343–347. doi: 10.1093/jmedent/25.5.343. [DOI] [PubMed] [Google Scholar]
- 27.Tesh R B, Chaniotis B N, Johnson K M. Science. 1972;175:1477–1479. doi: 10.1126/science.175.4029.1477. [DOI] [PubMed] [Google Scholar]
- 28.Tesh R B, Chaniotis B N, Peralta P H, Johnson K M. Am J Trop Med Hyg. 1974;29:1394–1404. doi: 10.4269/ajtmh.1974.23.258. [DOI] [PubMed] [Google Scholar]
- 29.Comer J A, Tesh R B, Modi G B, Corn J L, Nettles V F. Am J Trop Med Hyg. 1990;42:483–490. doi: 10.4269/ajtmh.1990.42.483. [DOI] [PubMed] [Google Scholar]
- 30.Corn J L, Comer J A, Erickson G A, Nettles V E. Am J Trop Med Hyg. 1990;42:476–482. doi: 10.4269/ajtmh.1990.42.476. [DOI] [PubMed] [Google Scholar]
- 31.Cupp E W, Mare C J, Cupp M S, Ramberg F B. J Med Entomol. 1992;29:137–140. doi: 10.1093/jmedent/29.2.137. [DOI] [PubMed] [Google Scholar]
- 32.Herrero M V, Jiménez A E, Rodríguez L L, Pereira R. J Med Entomol. 1994;31:912–914. doi: 10.1093/jmedent/31.6.912. [DOI] [PubMed] [Google Scholar]
- 33.Vargas M V, Travis B. Rev Biol Trop (Costa Rica) 1973;21:143–175. [PubMed] [Google Scholar]
- 34.Novella I S, Clarke D E, Quer J, Duarte E A, Lee C H, Weaver S, Elena S F, Moya A, Domingo E, Holland J J. J Virol. 1995;69:6805–6809. doi: 10.1128/jvi.69.11.6805-6809.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Tesh R B, Peralta P H, Johnson K M. Am J Epidemiol. 1969;90:255–261. doi: 10.1093/oxfordjournals.aje.a121068. [DOI] [PubMed] [Google Scholar]
- 36.Trainer D, Hanson R. Am J Epidemiol. 1969;90:192–198. doi: 10.1093/oxfordjournals.aje.a121080. [DOI] [PubMed] [Google Scholar]
- 37.Zuluaga F M, Yuill T M. Bull Pan Am Health Org. 1979;87:389–404. [Google Scholar]
- 38.Stallknecht D E, Fletcher W O, Erickson G A, Nettles V F. Am J Epidemiol. 1987;125:1058–1065. doi: 10.1093/oxfordjournals.aje.a114621. [DOI] [PubMed] [Google Scholar]
- 39.Webb P, Mclean R, Smith G, Ellenberger J, Francy B, Walton T, Monath T. J Wild Dis. 1987;23:192–198. doi: 10.7589/0090-3558-23.2.192. [DOI] [PubMed] [Google Scholar]
- 40.Kauffman S A. The Origins of Order: Self-Organization and Selection in Evolution. New York: Oxford Univ. Press; 1993. pp. 29–281. [Google Scholar]
- 41.Lederberg J, Shope R E, Oaks S C J. Emerging Infections: Microbial Threats to Health in the United States. National Academy Press, Washington, DC: Institute of Medicine; 1992. [PubMed] [Google Scholar]