

Abstract
Application of exogenous retinoic acid (RA) to zebrafish during the initial stages of photoreceptor differentiation results in a precocious development of rod photoreceptors and an inhibition of cone photoreceptor maturation. The acceleration of rod differentiation is observed initially within the ventral retina 3 days after fertilization, following 24 hr of RA application, and within the dorsal retina 4 days after fertilization, following 48 hr of RA application. The differentiation of rods was impeded significantly when the synthesis of endogenous retinoic acid was inhibited by citral prior to the initial stage of rod differentiation. RA-treated embryos labeled for bromodeoxyuridine (BrdU) uptake revealed that RA exerts its effect on a postmitotic cell population within the developing retina. During normal development in zebrafish, rod differentiation is most robust within the ventral retina, a region previously shown to be rich in RA. Our data suggest that the RA signaling pathway is involved in the differentiation and maturation of both the rod and cone photoreceptors within the developing zebrafish retina.
Keywords: retina, rod photoreceptor, cone photoreceptor, zebrafish, teleost
Recent experiments suggest that retinoic (vitamin A) acid (RA) plays an important role in early eye development. Application of an excess of RA to zebrafish embryos during the formation of the optic primordia—9.5–11.5 hr after fertilization (pf)—induces a proliferation of cells in the ventral part of the eye and the formation of a second retina (1). Inhibition of RA synthesis in the zebrafish eye at the same stage results in the formation of half-eyes that lack a ventral retina (2). A similar ocular phenotype is seen in embryonic mice in which RA receptors have been genetically eliminated via molecular manipulation (3). A study of the effects of RA on molecular markers expressed in early stages of zebrafish eye development indicates that RA exerts a ventralizing effect on the retina; i.e., it enhances ventral retinal characteristics while diminishing dorsal retinal features (4).
In postnatal rat and embryonic chicken cell cultures, RA promotes the differentiation and survival of retinal photoreceptors, suggesting that RA may also play a role later in development, in photoreceptor differentiation (5, 6). The adult zebrafish retina contains rods and four classes of cones that can be distinguished morphologically (7). Short single cones are ultraviolet-sensitive, whereas long single cones are blue-sensitive. The principal member of the double cone is red-sensitive, whereas the accessory member is green-sensitive (8).
Photoreceptors first develop at about 2 days pf in zebrafish in a patch of retina located ventrally (9). Subsequent photoreceptor differentiation in zebrafish is complex and does not occur uniformly across the retina (10, 11). Thus, distinct differences in the distribution and extent of maturation of rods and cones are observed in various retinal regions in zebrafish 3 days or older. In this study, we report the effects of RA on the early stages of photoreceptor cell differentiation in vivo. RA, or an RA synthesis inhibitor, were given exogenously to developing zebrafish embryos between 2 and 5 days pf.
MATERIALS AND METHODS
Retinoid and Citral Treatments.
Breeding fish were maintained and embryos were reared at 28°C as described (1). Frozen stocks of all-trans-RA, retinal, or retinol in dimethyl sulfoxide were thawed immediately before use and diluted in tank water. Retinoid stocks were prepared and administered in the dark. Embryos were immersed in tank water containing 0.3 μM RA, retinal, or retinol starting at 48 hr pf (hpf). Fresh RA solution was added every 24 hr. Treated embryos were raised to 3, 4, or 5 days pf and prepared for analysis. Citral was prepared as described (2). Fresh solutions of citral (ranging from 3 to 18 μM) were added every 1–1.5 hr for 18 consecutive hours starting at 48 hpf. After the final citral exposure, embryos were rinsed in fish tank water and allowed to develop for an additional 6 hr to 3 days pf before they were fixed for analysis.
Morphological Analysis.
Fixation, embedding, and sectioning procedures were similar to those previously described (12). Embryos prepared for light microscopic analysis were embedded in Epon-Araldite (EM Science), sectioned, and stained with 1% methylene blue/azure II in 1% borax buffer.
In Situ Hybridization.
The whole mount in situ hybridization protocol used was similar to that as described (13). Embryos were prehybridized for 3–5 hr at 65°C. Labeled probe at 0.5 ng/μl, final concentration, was added to prehybridized embryos, and the hybridization reaction was allowed to proceed at 65°C for 20 hr. RNA probes were generated as described (13) from plasmids containing cDNA of zebrafish red opsin, blue opsin, or rhodopsin, and goldfish ultraviolet opsin. After in situ whole mount staining, embryos were cleared in a glycerol series (25, 50, 70, 95, 100%) and mounted for photography in glycerol within deep-well slides without coverslips. Stained embryos to be sectioned were dehydrated and embedded in an Epon-Araldite mixture. Six-micrometer sections were cut on an LKB microtome and counter-stained with a 0.5% eosin solution.
Immunocytochemistry.
Monoclonal antibodies 5E11 and 1D1 were generated by standard methodologies from splenocytes isolated from a BALB/c mouse immunized with whole zebrafish retina in Ribi adjuvant (Ribi). Hybridoma supernatants were screened on both frozen sections of adult retina and Western blots of total retinal protein. Immunostaining was visualized using peroxidase-conjugated goat anti-mouse secondary antibodies (Sigma) and diaminobenzidine as substrate. Whole mount zebrafish embryos were prepared for antibody detection by the protocol as described (14) using horseradish peroxidase as a marker. Embryos were cleared in glycerol and photographed as described above or embedded in Epon-Araldite for histological analysis.
BrdU Labeling.
The yolk sacs of embryos at 48 hpf were microinjected with 10–20 nl of 10 mM BrdU (Sigma) dissolved in distilled water. The embryos were exposed to RA for 3 days and then were fixed for 4 hr in paraformaldehyde, permeabilized for 10 min in acetone (−20°C), rinsed in phosphate-buffered saline/0.1% Tween 20 (PBST, four 5-min washes), and exposed to proteinase K (20 μg/ml in PBST) for 1 hr. Embryos were then washed for 30 min in 2 M HCl, and rinsed in PBST (four 10-min washes), and antibody was added (anti-BrdU monoclonal, Becton Dickinson) at 1:50 dilution in blocking solution (PBST/1% goat serum) and incubated overnight at 4°C. After rinses in PBST, embryos were incubated overnight at 4°C in a secondary anti-mouse horseradish peroxidase (HRP)-conjugated antibody diluted 1:200 in blocking solution. Animals were then prepared for HRP staining, embedded into Epon-Araldite (as above), and sectioned for analysis.
RESULTS
Effects of RA on Rod and Cone Photoreceptor Development.
Transverse sections taken through the optic nerve of normal 5-day-old zebrafish reveal differences in the distribution of rods and cones within both the dorsal and ventral retina (Fig. 1). In more central regions dorsally and ventrally, short single cones are morphologically distinct from other cone types as they lie vitreal to a homogenous population of slender longer single cones or rods. Fig. 1A is a section taken in the midperipheral region of the dorsal retina. Short single cones are obvious more centrally (to the right) but less so peripherally (to the left). Rods are observed throughout this region of the retina (arrows). No rod photoreceptors are found immediately adjacent to the optic nerve at this age (data not shown) as noted earlier by Branchek and Bremiller (7).
Figure 1.
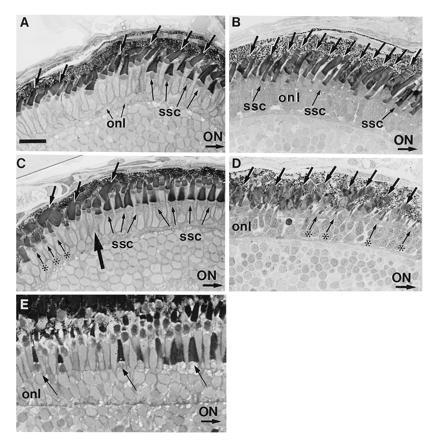
(A–D) Transverse sections of normal (A and C) and RA-treated (B and D) embryos on day 5 pf. In these sections, the optic nerve (ON) is to the right. (A) In the midperipheral region of the dorsal retina of control embryos, short single cones (ssc) with broad outer segments lie vitreal to rods and longer single cones. The short single cones are more obvious to the right, i.e., more centrally. Rod outer segments average 6 μm in length and are observed across all of this retinal region (arrows). (B) After a 3-day treatment with RA, the number of observable rods (arrows) has increased within the dorsal retina, and the short single cones (ssc) are smaller and less obvious. (C) The ventral retina in control eyes is partitioned into two compartments: a cone-dominated region (right of large arrow) adjacent to the optic nerve and a peripheral region (left of large arrow) dominated by rods (arrows). Mature short single cones (ssc) lie vitreal to a layer of longer single cones in the cone-dominated region. In the rod-dominated region, a population of miniature cones (asterisks) are seen. (D) After RA treatment beginning at day 2 pf, the cone-dominated region within the ventral retina is not observed. Rather large rod outer segments (arrows) are observed extending across the entire ventral region rather than being restricted to the periphery. These rod outer segments appear less organized than those of controls and average ≈10 μm in length, which is ≈2 μm longer than those observed in the ventral region of controls. Small cones (asterisks) resembling the miniature cones located within the ventral periphery in controls extend across the entire ventral region. (E) Transverse section of the peripheral retina from a normal retina at 21 days pf. Arrows identify maturing short single cones (from left to right). onl, Outer nuclear layer. (A–D, bar = 16 μm; E, bar = 9 μm.)
The retina ventral to the optic nerve is sharply partitioned into a cone-dominated region immediately adjacent to the optic nerve (to the right of large arrow in Fig. 1C) and a rod-dominated region located somewhat more peripherally (left of large arrow, Fig. 1C). The cone-dominated region is characterized by a distinct row of short single cones possessing broad well-formed outer segments. As in the dorsal retina, these short single cones lie vitreal to a homogenous population of longer slender single cones. In the far periphery of the ventral retina, rod outer segments up to ≈8 μm in length are seen. In addition, a population of tiny single cones is observed prominently in this peripheral region (asterisks). These tiny cones possess outer segments almost as long as adult cones, but they are much thinner than more mature cones. Similar miniature cones are observed in the peripheral retina of adult teleosts (15) where new photoreceptors and other retinal cells are continually being formed during life (Fig. 1E). We believe these tiny cones to be immature cones.
RA treatment between 2 and 5 days pf alters photoreceptor development and maturation. Transverse sections through the optic nerve taken from RA-treated zebrafish reveal a significant increase in the number of recognizable rod photoreceptors throughout both the dorsal and ventral retina. Fig. 1B shows a retinal area from a RA-treated animal comparable to the area shown in Fig. 1A. The increase in numbers of observable rods (large arrows) is striking, and rod outer segments average ≈8 μm in length, which is ≈2 μm longer than those of controls. Furthermore, the distinct layer of short single cones seen in Fig. 1A is not observed
Ventral to the optic nerve (Fig. 1D), the prominent cone-dominated region adjacent to the optic nerve seen in controls is no longer apparent. Instead, numerous rods with large outer segments extend throughout the ventral retina (large arrows), including the region immediately adjacent to the optic nerve. Ventrally, rod outer segments in RA-treated animals are broader than those found dorsally and they average ≈10 μm in length, which is ≈2 μm greater than that of controls. Although the cone-dominated region is no longer observed, a number of tiny cones with well-formed outer segments are present in this area (asterisks in Fig. 1D). The size and shape of these tiny cones strongly resembles the tiny cones located within the ventral periphery of control animals (Fig. 1E), suggesting that RA has inhibited the maturation of these cones.
Effect of RA on Opsin Expression.
Rods. To analyze in more detail the effects of RA on rod development, a RNA probe marker to the rhodopsin gene product was used for whole mount in situ hybridization studies. Rhodopsin mRNA expression is localized primarily to the inner segments of the rod photoreceptors. In 3-day control animals, rhodopsin mRNA expression was observed exclusively within the ventral-most region of the eye. The whole mount photographs in Fig. 2 show the ventral surface of the eye. The rhodopsin-expressing cells are organized as a band extending longitudinally along the nasal-temporal axis of the ventral retina. This staining pattern enlarges and increases in density at both day 4 (Fig. 2B) and day 5 (Fig. 2C).
Figure 2.
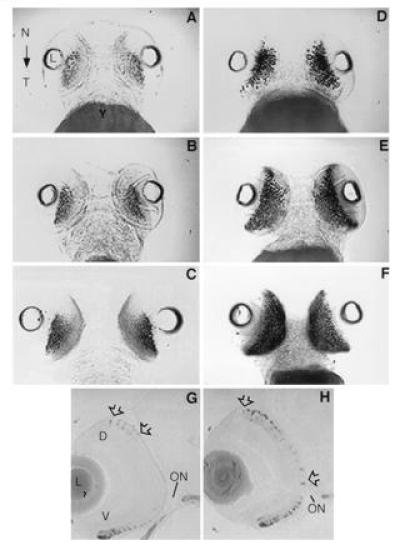
Control embryos (A–C) and RA-treated embryos (D–F) in ventral view showing the localization of rhodopsin mRNA expression by whole mount in situ hybridization. In controls, the level of rhodopsin mRNA expression increases modestly within the ventral retina during development through 3 (A), 4 (B), and 5 (C) days pf. After a 24-hr RA treatment beginning at day 2, staining within the ventral region is more robust indicating the level of rhodopsin expression has increased significantly (D). Not only is rhodopsin expression greater but it extends further into the nasal and temporal regions of the retina as compared with controls. After 2 and 3 days of RA treatment, the level of rhodopsin expression is further increased relative to the controls and extends throughout the ventral region as far as the nasal and temporal margins at days 4 (E) and 5 pf (F), respectively. The localization of rhodopsin transcripts by in situ hybridization at day 4 in transverse sections (5 μm) in control (G) and RA-treated (H) embryos. (G) On day 4, rhodopsin expression is evident within the periphery of the ventral retina (V). Weak expression is also observed near the dorsal (D) periphery (arrows). (H) After 2 days of RA treatment, rhodopsin expression extends throughout the ventral retina. Furthermore, expression is considerably more robust and extensive within the dorsal retina (arrows). ON, optic nerve; N, nasal region; T, temporal region; L, lens; Y, yolk.
A 24-hr exposure to RA significantly expanded the number of cells showing rhodopsin mRNA expression and also increased the density of staining in individual cells within the ventral region of the eye at day 3 pf (Fig. 2D). A 24-hr exposure of embryos to similar concentrations of retinol or retinal did not have this effect. Prolonged exposure of zebrafish embryos to RA (for 48 and 72 hr) further increased mRNA expression at days 4 and 5 pf (Fig. 2 E and F), particularly along the nasal-temporal axis of the ventral retina. An RA-induced increase in the number of stained photoreceptors was also observed in whole mount antibody preparations at 5 days of development with the use of the zebrafish-rod-specific monoclonal antibody 1D1 (data not shown). Immunohistochemical staining patterns were similar to the rhodopsin mRNA expression patterns in RA-treated embryos.
In sections of control embryos, rod photoreceptors expressing the rhodopsin gene product can be observed initially within the dorsal region of the developing retina at day 4 pf (Fig. 2G). Those cells expressing rhodopsin within the dorsal retina are localized to the dorsal-most third of the developing retina in controls. A 48-hr exposure of embryos to RA beginning at day 2 pf expanded significantly the spatial expression pattern of the rod photoreceptor marker within the dorsal region of the retina (Fig. 2H). Indeed, cells expressing rod opsin were now seen throughout half of the dorsal retina in day 4 embryos (Fig. 2G). A further expansion of rod opsin expression is seen at day 5 pf (data not shown). An increase in rod staining is observed also in the ventral retina at 4 days pf (Fig. 2H).
Cones.
To determine the effects of RA on cone development, cell-specific RNA probes for the ultraviolet (UV), blue, and red opsin mRNAs were used for whole mount in situ hybridization. RA was given between 2 and 5 days pf, and embryos were examined at day 5. Opsin expression was significantly decreased in the eyes of RA-treated embryos hybridized with the UV-opsin probe, particularly within the ventral retina (Fig. 3 A and B). The size of the UV-cones was also reduced in this retinal region (data not shown). Significant differences were not observed in whole mount staining patterns for the red or blue opsins. However, in sections taken from whole mount preparations hybridized with both the red and blue opsin RNA probes, differences could be observed, particularly in sections stained with the blue opsin RNA probe. (Fig. 3 C and D). The blue cones appeared to be smaller and opsin expression was weaker, particularly in the ventral region of the retina. In all cases, the staining pattern in treated embryos for red, blue, and UV opsins was graded along the dorsal-ventral axis of the eye; levels of opsin expression were greatest near the dorsal periphery and gradually diminished ventrally. Sections taken from control embryos did not display this graded pattern of expression.
Figure 3.
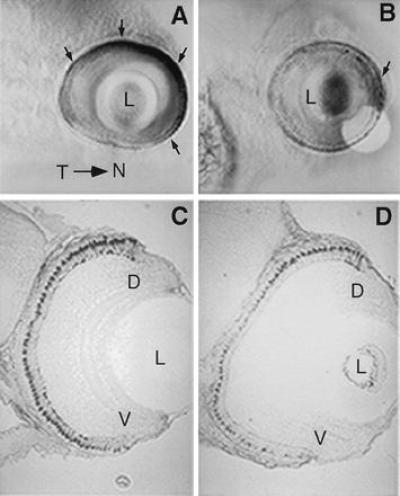
Localization of mRNA transcripts for ultraviolet opsin by whole mount in situ hybridization at day 5 pf in control embryos (A) and RA-treated embryos (B) in lateral view. (A) In controls, ultraviolet opsin expression is most robust within the nasal retina and faint expression is also observed in the temporal retina (arrows indicate regions of high expression). (B) After three days of RA treatment, ultraviolet opsin expression is significantly decreased at day 5 in RA-treated embryos throughout most of the retina. The only region of dense staining is found within the nasal retina (arrow). The localization of blue opsin transcripts by in situ hybridization at day 5 in transverse sections (5 μm) in control (C) and RA-treated (D) embryos. Blue opsin staining is generally weaker in RA-treated embryos (D). Furthermore, staining patterns of cells expressing the blue opsin appears graded in RA-treated embryos along the dorsal-ventral axis of the retina. The size of cones expressing the blue-sensitive opsin is decreased, particularly in regions ventral to the dorsal retina. D, dorsal retina; V, ventral retina; L, lens; Y, yolk.
Effects of RA on Other Retinal Neurons and Eye Shape.
Morphological analysis revealed no changes within populations of other retinal cells in embryos treated with RA between 2 and 5 days pf. Cell markers specific to zebrafish ganglion and amacrine cells were used to search for RA-induced alterations in these cell populations. No abnormalities in number or location of ganglion and amacrine cells were detected within RA-treated retinas. Although all cell layers of the retina except for the photoreceptor cell layers appeared unaffected by prolonged RA treatments, the general size and shape of the eye was slightly altered. The eyes of embryos treated with RA between 2 and 5 days pf were slightly smaller and appeared more rounded than the eyes of controls (compare Fig. 3 A and B).
Effects of RA Inhibition on the Ventral Retina.
In the zebrafish retina, RA synthesis is mediated in the dorsal retina by a class-1 aldehyde dehydrogenase and in the ventral retina by a different more effective retinaldehyde dehydrogenase (2). This arrangement results in substantially higher RA levels in the eye ventrally. By exposing 2-day zebrafish embryos to citral, a competitive inhibitor of the dehydrogenase that converts endogenous retinaldehyde to RA within the ventral region of the retina, a dose-dependent inhibition of rod opsin expression was observed within the ventral region of the developing eye (at day 3). Citral treatments of 3 μM greatly reduced the number of cells expressing the rod-specific opsin marker (compare Fig. 4 A and B) but did not have observable effects of cone cell types expressing the ultraviolet-, blue-, or red-sensitive opsins (data not shown). Other retinal cell types within the ventral retina (analyzed by light microscopy) were also not altered by this treatment. Higher concentrations (6 μM) of citral (Fig. 4C) further decreased the number of rhodopsin-expressing cells within the ventral retina, but retinas exposed to 6, 12, or 18 μM citral exhibited toxic effects upon development. These embryos appeared smaller and retinal tissue was less developed with these higher citral concentrations. A dose-dependent increase in cell death was observed in the retinas and other brain regions of animals exposed to increased citral concentrations.
Figure 4.
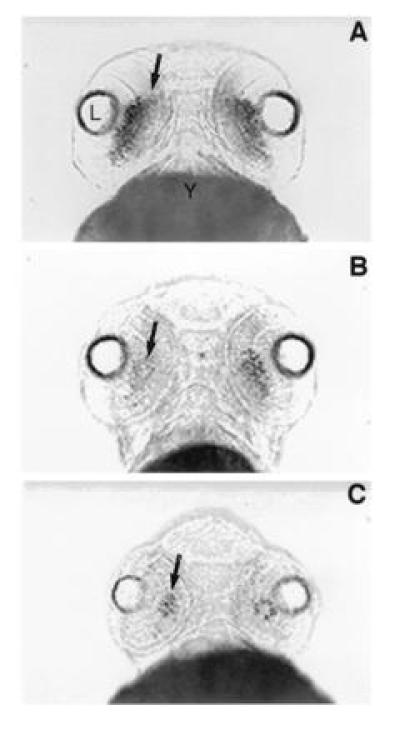
Ventral views of rhodopsin expression in whole mount in situ preparations at day 3 pf in control embryos (A) and embryos treated with the RA-synthetic competitive inhibitor citral (B and C). (A) Rhodopsin expression (arrow) extends across the ventral retina in control embryos at day 3. (B) After treatment with 3 μM citral between days 2 and 3 pf, the level of rhodopsin expression (arrow) is significantly reduced. (C) After treatment with 6 μM citral, rhodopsin expression is further reduced such that only a small number of rods (arrow) are observed within the retina at day 3 pf. L, lens; Y, yolk.
Effects of RA on Retinal Cell Proliferation.
To determine if the effects of RA were upon postmitotic cells, inducing them to differentiate into rod photoreceptors, or if RA exposure increased the proliferation and, therefore, the number of cells differentiating into rods, embryonic yolk sacks were injected with BrdU prior to RA exposure. The presence of BrdU within the developing embryo throughout the 3 days of treatment labeled all cells in S phase of the cell cycle during this period. The exposure of embryos to RA for 3 consecutive days did not increase the number of cells within the retina containing the BrdU label. There were few cells in sectioned control and RA-treated embryos (one or two per section) containing the BrdU label within the photoreceptor layer of the retina. BrdU nuclear labeling was similar in control and RA-treatment groups and was localized primarily to the proliferative zone of the retina at the extreme margins of the eyecup.
DISCUSSION
In this study we demonstrate that RA induces a precocious development of rod photoreceptors within the developing zebrafish retina. This effect is seen initially at day 3 in the ventral region of the retina after 24 hr of RA treatment. By day 4, an RA-induced acceleration in rod development is observed also within the dorsal retina after a 48-hr exposure to RA. These effects of RA on rod development were observed both by histological examination of the developing retina and by in situ hybridization studies of rod opsin expression. Our data indicate that RA promotes rod photoreceptor development throughout the retina but that the ventral retina responds more rapidly to exogenously applied RA than does the dorsal retina.
An earlier study demonstrated that RA is synthesized within the dorsal and ventral regions of the zebrafish embryo by two distinct dehydrogenases, both of which convert retinaldehyde to RA (2). The dehydrogenase located within the ventral retina arises earlier in neurogenesis (13–15 hpf) than does the dorsal enzyme, which appears approximately 10 hr later (≈24 hpf). In addition, the activity of the ventral enzyme exceeds that of the dorsal enzyme throughout development. This produces an asymmetric distribution of RA in the retina with high levels of RA located ventrally in the developing eye and lower levels of RA dorsally. The differentiation of rods within the ventral and dorsal regions of the developing zebrafish retina may correlate with the levels of RA found in these locations; rod photoreceptors first appear within the ventral retina at 50 hpf and they differentiate there at a rapid rate thereafter. Rods in the dorsal retina, on the other hand, are not seen until about ≈12 hr later (≈62 hpf) and are added more slowly subsequently than are rods in the ventral regions of the retina. Why ventral receptors develop earlier and are more responsive to RA than dorsal retinal cells is not clear but could relate to the higher endogenous levels of RA found in the ventral retina.
The effect of RA on cones is to delay their maturation. This effect is seen clearly on day 5, after 3 days of RA treatment. Mature cones are not observed in the central regions of 5-day-old RA-treated retinas as is the case in control retinas. Instead, many small miniature cones are observed throughout the retina of such RA-treated animals. These miniature cones are presumed to be cone photoreceptors arrested in their development. In the adult retina, such small miniature cones are observed only in the extreme periphery of the retina where new cones (and other retinal cells) are continuously being formed (15). In untreated embryos at day 5, miniature cones are observed most prominently in the ventral regions of the eye where endogenous RA levels are high.
As was the case for rods, RA appears to have greater effects on cones in the ventral retina than in the dorsal retina. In RA-treated retinas, the largest and most mature cones were observed in the most peripheral regions of the dorsal retina. Cones became increasingly more reduced in size moving toward the ventral retinal periphery. These histological observations were supported by gene expression studies; levels of cone opsin expression were greater dorsally than ventrally in the retinas of RA-treated animals Again, cells in the ventral retina may be more responsive to RA because of the high endogenous level of RA found there.
Inhibition of RA synthesis in the eye by citral, an inhibitor of the ventral retinal enzyme, clearly suppressed rod opsin expression in the ventral region of the eye. This was a dose-dependent effect, although at higher citral concentrations, toxic effects of the citral were noted. We did not observe effects of citral on cone opsin expression, but this may reflect the fact that the effects of RA on rod opsin expression were generally more robust than the effects of RA on cone opsin expression.
How RA induces precocious rod development in zebrafish is not clear. A retinoic acid response element (RARE) has been identified in the promoter region of the bovine rhodopsin gene (D. Zack, personal communication) and RAREs may also be present in human red and green cone opsin promoter regions (A. Goodman, personal communication). Activation of RAREs could explain the effects of RA on both rod and cone opsin expression in zebrafish.
In the larval and adult teleost retina, rod photoreceptors are generated by a population of rod precursor cells that remain mitotic throughout life (16, 17). One possibility we considered was that RA not only accelerates rod development but also promotes the proliferation of these precursor cells, thus increasing the number of rods in the retina. However, embryos labeled with BrdU prior to RA treatment were identical to labeled control embryos. Thus, we conclude that the cells that respond to RA are postmitotic and are specified to become rod photoreceptors. This conclusion is supported by our finding that other retinal cell populations (ganglion and amacrine cells) were not affected by RA treatments; that is, precursors of these cells were not recruited by RA to become rod cells as was the case in rat cell cultures (5).
A number of factors, in addition to RA, have been shown to promote rod photoreceptor cell differentiation in retinal cell cultures, including cilliary neurotrophic factor (18), acidic fibroblast growth factor (19), basic FGF (20), and taurine (21). It could be that expression of several of these factors is enhanced by RA in the retina as RA is known to induce the expression of several growth factors in various tissues. RA has also been shown to affect certain cone types, particularly UV-cones, in trout retinas (22). RA causes a precocious loss of UV-cones in young trout but then induces the reappearance of these cone cells in older fish. Our results, taken together with earlier reports, indicate that photoreceptor development is regulated by a variety of diffusible factors and RA is likely to be one of these factors.
Acknowledgments
We thank Ursula Drager for reviewing the manuscript and for helpful comments. We also thank Bill McCarthy for providing zebrafish embryos from the Harvard University zebrafish facility. This work was supported by National Institutes of Health Grant EY-00811 to J.E.D., a grant from the National Retinitis Pigmentosa Foundation, Inc., a Fellowship from Knights Templar Foundation to E.A.S., and by National Institutes of Health Grant EY-06554 to J.M.F.
Footnotes
Abbreviations: RA, retinoic acid; pf, after fertilization; hpf, hours pf.
References
- 1.Hyatt G A, Schmitt E A, Marsh-Armstrong N R, Dowling J E. Proc Natl Acad Sci USA. 1992;89:8293–8297. doi: 10.1073/pnas.89.17.8293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Marsh-Armstrong N, McCaffery P, Gilbert W, Dowling J E, Drager U C. Proc Natl Acad Sci USA. 1994;91:7286–7290. doi: 10.1073/pnas.91.15.7286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Kastner P, Grondona J M, Mark M, Gansmuller A, LeMeur M, Decimo D, Vonesch J-L, Dolle P, Chambon P. Cell. 1994;78:987–1003. doi: 10.1016/0092-8674(94)90274-7. [DOI] [PubMed] [Google Scholar]
- 4.Hyatt G A, Schmitt E A, Marsh-Armstrong N R, McCaffery P, Drager U C, Dowling J E. Development (Cambridge, UK) 1996;122:195–204. doi: 10.1242/dev.122.1.195. [DOI] [PubMed] [Google Scholar]
- 5.Kelley M W, Turner J K, Reh T A. Development (Cambridge, UK) 1994;120:2091–2102. doi: 10.1242/dev.120.8.2091. [DOI] [PubMed] [Google Scholar]
- 6.Stencamp D L, Gregory J K, Adler R. Invest Ophthalmol Vis Sci. 1993;34:2425–2436. [PubMed] [Google Scholar]
- 7.Branchek T, Bremiller R. J Comp Neurol. 1984;224:107–115. doi: 10.1002/cne.902240109. [DOI] [PubMed] [Google Scholar]
- 8.Robinson J, Schmitt E A, Harosi F I, Reece R J, Dowling J E. Proc Natl Acad Sci USA. 1993;90:6009–6012. doi: 10.1073/pnas.90.13.6009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Kljavin I. J Comp Neurol. 1987;260:461–471. doi: 10.1002/cne.902600311. [DOI] [PubMed] [Google Scholar]
- 10.Raymond P A, Barthel L K, Curran G A. J Comp Neurol. 1995;359:537–550. doi: 10.1002/cne.903590403. [DOI] [PubMed] [Google Scholar]
- 11.Schmitt E A, Dowling J E. J Comp Neurol. 1996;371:222–234. doi: 10.1002/(SICI)1096-9861(19960722)371:2<222::AID-CNE3>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- 12.Schmitt E A, Dowling J E. J Comp Neurol. 1993;344:532–542. doi: 10.1002/cne.903440404. [DOI] [PubMed] [Google Scholar]
- 13.Oxtoby E, Jowett T. Nucleic Acids Res. 1993;21:1087–1095. doi: 10.1093/nar/21.5.1087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Puschel A W, Westerfield M, Dressler G R. Mech Dev. 1992;38:197–208. doi: 10.1016/0925-4773(92)90053-m. [DOI] [PubMed] [Google Scholar]
- 15.Muller H. Zoologisches Jarbuch. 1952;63:275–324. [Google Scholar]
- 16.Sandy J T, Blaxter J H. J Mar Biol Assoc UK. 1980;60:59–71. [Google Scholar]
- 17.Johns P R, Fernald R D. Nature (London) 1981;293:141–142. doi: 10.1038/293141a0. [DOI] [PubMed] [Google Scholar]
- 18.Fuhrmann S, Kirsch M, Hofmann H-D. Development (Cambridge, UK) 1995;121:2695–2706. doi: 10.1242/dev.121.8.2695. [DOI] [PubMed] [Google Scholar]
- 19.Hicks D, Courtois Y. FEBS Lett. 1988;234:475–479. doi: 10.1016/0014-5793(88)80141-8. [DOI] [PubMed] [Google Scholar]
- 20.Hicks D, Courtois Y. J Neurosci. 1992;12:2022–2033. doi: 10.1523/JNEUROSCI.12-06-02022.1992. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Altshuler D, Lo Turco J J, Rush J, Cepko C. Development (Cambridge, UK) 1993;119:1317–1328. doi: 10.1242/dev.119.4.1317. [DOI] [PubMed] [Google Scholar]
- 22.Browman H I, Hawryshyn C W. J Exp Biol. 1994;193:191–207. doi: 10.1242/jeb.193.1.191. [DOI] [PubMed] [Google Scholar]