

Abstract
The emergence of modern humans in the Late Pleistocene, whatever its phylogenetic history, was characterized by a series of behaviorally important shifts reflected in aspects of human hard tissue biology and the archeological record. To elucidate these shifts further, diaphyseal cross-sectional morphology was analyzed by using cross-sectional areas and second moments of area of the mid-distal humerus and midshaft femur. The humeral diaphysis indicates a gradual reduction in habitual load levels from Eurasian late archaic, to Early Upper Paleolithic early modern, to Middle Upper Paleolithic early modern hominids, with the Levantine Middle Paleolithic early modern humans being a gracile anomalous outlier. The femoral diaphysis, once variation in ecogeographically patterned body proportions is taken into account, indicates no changes across the pre-30,000 years B.P. samples in habitual locomotor load levels, followed by a modest decrease through the Middle Upper Paleolithic.
Keywords: Paleoanthropology, human origins, Late Pleistocene, postcrania
The middle of the Late Pleistocene, between approximately 100,000 and 30,000 years B.P., saw the emergence and establishment of a novel constellation of human biological characteristics. This evolutionary process, known as the “origins of modern humans,” led to the presence across the Old World by ca. 30 thousands of years (kyr) B.P. of a new biological complex to the exclusion of the one which, with evolutionary modification, had been present throughout archaic Homo for the previous 1.7+ million years. With minor evolutionary changes through time and space, this biological pattern remains in place in the present world.
Our comprehension of the evolutionary emergence of modern humans rests primarily on our ability to decipher from the paleoanthropological record the patterns and processes of change, whether adaptive or stochastic, regional or global, which enabled one biological pattern to replace a previously highly successful one in a relatively short period of geological time. This paleobiological and paleoanthropological problem, although dependent on neontological uniformitarian patterns for explanatory reference, can be resolved only through the analysis of the prehistoric record, both paleontological and archeological.
With this problem in mind, paleoanthropological research has included attempts to decipher the patterns and degrees of change of functionally relevant aspects of human biology during the Late Pleistocene. Given the nature of the hominid fossil record with its abundance of fossils from Europe and western Asia and the dearth of reasonably complete remains from elsewhere in the Old World, this research has been focused on the paleoanthropological record of the northwestern Old World. However, sufficient remains are now known from less well represented areas to indicate that, once normal stochastic and ecogeographical patterns of interregional variation within species of large-bodied mobile terrestrial mammals are taken into account, the northwestern Old World is generally representative of the more global patterns of human biology.
With this information in mind, we have been investigating patterns of Late Pleistocene hominid diaphyseal appendicular robusticity by using cross-sectional geometry (1–5). Given the high degree of plasticity of the mammalian diaphyseal cortical bone, especially during development (2, 6, 7), this approach provides a paleobiological window on the habitual activity levels of extinct hominid populations. Moreover, potentially contrasting patterns of upper vs. lower limb diaphyseal cortical hypertrophy allow insights into manipulative vs. locomotor activity levels, thus shedding light on two of the most important aspects of hominid behavioral evolution.
A Genealogical Digression
At the same time, the majority of the research on Late Pleistocene hominid evolution beyond philatelic concerns has been focused on the phylogenetic relationships of geographical groups of late archaic and early modern humans. And despite a century of debate on this issue with the progressive introduction of more diverse and higher-quality data and analytical techniques, combined with more global approaches to the problem, there is little consensus. Indeed, the current and ongoing debate on the phylogenetic aspects of modern human emergence appears to be more concerned with hypothesis confirmation than with hypothesis testing.
The past decade has seen the increasing application of human molecular data to issues of modern human origins. However, with one exception (8), the molecular data that have been brought to bear on the issue have no empirical time depth, only probabilistic inferential time depth dependent on both the nature of the data and the layered analytical assumptions behind the various quantitative techniques used to process those data. Moreover, all of these analyses assume a highly uniform stochastic accumulation of genetic change (i.e., a molecular clock that keeps accurate time throughout the last half-million years) and/or geographically uniform human demographic stability throughout the Middle and Late Pleistocene. These assumptions are simply untenable. Any reasonable assessment of molecular data, analytical techniques, and processes makes it highly unlikely that the standard errors of estimates of divergence times are sufficiently small to be useful. The geographical and demographic fluctuations of Pleistocene hominid populations, given both their foraging adaptive patterns and susceptibility to major Pleistocene climatic fluctuations, make any assumptions of uniform population size and distribution implausible, even for short periods of the last half-million years.
The fossil data as it pertains to strictly phylogenetic issues are not much better except in Atlantic Europe, a peripheral cul-de-sac where the transition was very late, relatively abrupt, and probably unrepresentative of more global patterns. Elsewhere across the Old World the human paleontological evidence is sufficiently ambiguous to be interpretable as indicating varying degrees of population continuity, replacement, and/or gene flow. Moreover, the biological bases and hence phylogenetic usefulness of most of the morphological traits commonly used are simply unknown, making it uncertain what is being analyzed.
The simple accumulation of additional neontological and paleontological data and its analysis by current techniques are insufficient for the resolution of these phylogenetic issues. For these reasons, it may well be scientifically more profitable, once one reasonably can define paleontological samples and their distributions in time and space, to look at changing patterns of biology and behavior, no matter what the original genealogical relationships were between the groups.
Materials and Methods
Paleontological Samples.
Given the dearth of associated partial skeletons and largely intact long bones from most of the Late Pleistocene of Africa and eastern Asia, the analysis here is focused on two primary samples, one of late archaic humans from the northwestern Old World and the other of early modern humans (defined on the basis of cranio-facial and nondiaphyseal postcranial morphology) from across Eurasia. Despite minor trends through time in facial gracilization among the mostly Middle Paleolithic-associated late archaic humans, they represent a similar group across this geographical area with apparent stasis in most aspects of postcranial morphology. The early modern human group, however, combines three groups. The first is of Levantine Middle Paleolithic hominids from middle of oxygen isotope stage 5, and it may well represent [based on body proportions (9, 10) and associated fauna (11)] a temporary dispersal into the region from northeastern Africa. The second is a small sample of European Early (pre-30 kyr B.P.) Upper Paleolithic humans, and the third is of east Asian, Near Eastern, and European Middle Upper Paleolithic-associated humans, from <30 kyr to 18 kyr B.P. Specimens with pathological lesions that appear to have altered habitual biomechanical load levels (e.g., Neandertal 1 humeri) were not included.
Methods.
To maximize the accuracy and biomechanical relevance of the analysis, the diaphyses of long bones were compared by using cross-sectional areas and second moments of area (also known as area moments of inertia), analyzed in the context of variation in body proportions. Experience has shown that analyses using only external measures of diaphyseal size and lacking the integration of ecogeographical patterning in body shape can provide misleading results. Appendicular robusticity therefore was assessed by computing cross-sectional geometric parameters of all of the available humeri and femora, here presented for the humeral mid-distal (35%) diaphysis and the femoral midshaft (50%). Given the near universality of right-side upper limb dominance among these extinct hominids, associated with variable levels of humeral asymmetry, only right humeri were considered.
Cross sections were reconstructed noninvasively by using transcribed molds of sub-periosteal contours combined with bi-planar radiography for the parallax-adjusted determination of cortical thicknesses, from which the endosteal contours were interpolated. Cross-sectional parameters (total and cortical area, anatomically oriented and maximum-minimum second moments of area, and the polar moment of area) were computed from digitized cross sections by using a PC version of slice (12, 13). In this framework, cortical area represents structural resistance to axial loading, second moments of area indicate resistance to bending in the plane in question, and polar moments of area approximate strength relative to torsional forces. Furthermore, because the polar moment of area is the sum of any two perpendicular second moments of area, it also provides an indication of overall biomechanical structural integrity.
The resultant parameters for the samples are compared graphically, for the humerus and initially for the femur by using lne-lne plots of the resultant values. For the humerus, given its normally non-weight-bearing role in humans, the logarithmic transformation appears to be adequate to adjust for allometric effects, especially of cross-sectional measures vs. bone length. For the femur, however, load levels are dependent on both body mass (weight and momentum) and beam characteristics. To correct for documented variance of ecogeographically patterned Late Pleistocene human body proportions (9, 10, 14–16), bi-iliac breadth was used to represent variance in body laterality, and femoral length was used for both beam length and to represent stature. Given the relative constancy of bi-iliac breadth within ecogeographically defined human groups (14), bi-iliac breadth was estimated from femoral length for specimens lacking sufficiently complete pelvic remains for direct determination of bi-iliac breadth. This calculation was done by using the mean associated bi-iliac breadth (BIB) and femoral length (FL) of the larger relevant sample, such that:
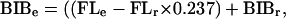 |
in which BIBr and FLr are the mean values for the reference sample and BIBe and FLe are the values for the specimen in question. From these data, the difference in relative body laterality (BIB/FL) between the specimen in question and a recent human reference sample [Pecos Pueblo Amerindians: mean BIB/FL = 0.63 (15)] (k) was determined. Because cortical area (CA) primarily reflects resistance to axial loads, it scales to FL3, or:
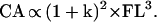 |
Second moments of area (Ii) and polar moments of area (J) scale to both an estimate of body mass (∝ FL3) and beam length (∝ FL), all raised to the 4/3 power (see ref. 1 for derivation and justification). Introducing variance in body breadth, this scaling becomes:
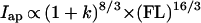 |
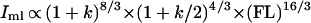 |
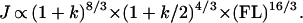 |
From these formulae, “adjusted femoral lengths” were computed, against which are plotted the raw values for individual second and polar moments of area.
Past research on Late Pleistocene human appendicular robusticity (1, 4, 17, 18) has provided conflicting assessments of the degree of contrast between these samples. Consequently, the distributions of measures of diaphyseal robusticity were statistically evaluated with an Ho of similarity. These degrees of similarity between the samples were assessed by using standard residuals from the pooled Late Pleistocene sample. They were compared between the late archaic and pooled early modern human samples by using unpaired Wilcoxon rank-sum tests and across combinations of multiple samples with Kruskal–Wallis tests.
Patterns of Humeral Robusticity
Assessments of Pleistocene diaphyseal robusticity frequently focus on percent cortical area. It sometimes is incorrectly referred to as a narrowing of the medullary canal (medullary stenosis), but it is usually a product of reduced endosteal resorption during development combined with greater subperiosteal deposition (3). The distribution of cortical area relative to total subperiosteal area for Late Pleistocene mid-distal humeri (Fig. 1), however, shows no separation of the samples. There is only a nonsignificant tendency (P = 0.104) for the late archaic humans to have greater relative cortical area. The three early modern human samples are indistinguishable (P = 0.209).
Figure 1.

Bivariate plots of humeral mid-distal diaphyseal cortical area vs. total area (Top) and humeral length (Middle), plus mid-distal polar moment of area vs. humeral length (Bottom). Solid pentagons, Eurasian late archaic humans; solid squares, Levantine Middle Paleolithic early modern humans; gray diamonds, European Early Upper Paleolithic early modern humans; empty triangles, Eurasian Middle Upper Paleolithic early modern humans.
Associated with this similarity in relative cortical area within the cross section, the late archaic human sample exhibits significantly greater cortical areas and polar moments of area relative to humeral length (P = 0.038 and 0.042, respectively) (Fig. 1). Given that humeral length can be taken as a surrogate measurement for overall body size and closely approximates upper arm beam length, these data indicate generally greater loading of the upper limb during manipulative activities among these late archaic humans.
Despite major differences in the associated technologies (Middle Paleolithic vs. Early Upper Paleolithic vs. Middle Upper Paleolithic), there is no significant difference across the early modern human samples in relative cortical area (P = 0.409) and only a tendency toward a significant difference (P = 0.067) in relative polar moment of area. Interestingly, however, it is the Levantine Middle Paleolithic-associated early modern humans who exhibit the most gracile humeri. Within the earlier Upper Paleolithic, the few Early Upper Paleolithic humeri cluster in the zone of overlap between the late archaic humans and the Middle Upper Paleolithic sample.
Consequently, it is possible to see a frequency shift in humeral diaphyseal robusticity from late archaic humans to Early Upper Paleolithic ones to Middle Upper Paleolithic ones, a pattern that follows the increasing sophistication and mechanical effectiveness of the associated technologies. The Middle Paleolithic Levantine sample, however, is anomalous with respect to both its time horizon (alternating through oxygen isotope stage 5 with late archaic humans) and its technological associations.
Patterns of Femoral Robusticity
As with the humerus, femoral midshaft cortical area vs. total area is a poor discriminator of Late Pleistocene human samples (Fig. 2), with an overall P = 0.099, an archaic/modern P = 0.104, and an early modern human P = 0.173. There is a visual tendency for the late archaic and Early Upper Paleolithic humans to have the relatively higher values and the Middle Upper Paleolithic ones to have some of the lower values, but the degree of overlap makes any inferences tentative.
Figure 2.

Bivariate plots femoral midshaft cortical area vs. total area (Top) and femoral length (Middle), plus midshaft polar moment of area vs. femoral length (Bottom). Symbols are as in Fig. 1.
When midshaft cortical area and polar moment of area are plotted against femoral length (Fig. 2), there is a highly significant difference between the late archaic and early modern human samples (both P < 0.001). However, there are no significant differences across the early modern human samples (P = 0.312 and 0.422 respectively). These patterns hold despite considerable within sample variation in both relative cortical area and relative polar moment of area, complete overlap between the early modern human samples, and some degree of overlap between the late archaic human remains and each of the early modern human ones.
However, as mentioned above, there are ecogeographically patterned contrasts in body proportions between these samples. In this regard, the late archaic humans exhibit hyperarctic body proportions (especially wide bi-iliac breadths), the Levantine early modern human sample has linear equatorial body proportions, and European earlier Upper Paleolithic human populations exhibit warm temperate to tropical proportions (9, 10, 14–16). Only some of the later (<20 kyr B.P.) Middle Upper Paleolithic humans exhibit proportions similar to modern populations in the same regions (10).
The adjustment of femoral length for body laterality, and hence for much of the variation in body proportions (see Methods), provides a rather different pattern (Fig. 3). With respect to cortical area vs. adjusted femoral length, there is only a suggestion of an overall difference between the samples and of the late archaic humans remaining more robust (P = 0.107 and 0.089, respectively). However, if the differences between only the three older samples (excluding the Middle Upper Paleolithic human remains) are evaluated, any tendency toward differentiation disappears (P = 0.758). In the mechanically more comprehensive polar moment of area, which combines both the quantity and distribution of bone in the cross sections, there is little difference across the samples (P = 0.380). Moreover, deletion of the Middle Upper Paleolithic sample reveals nearly identical proportions across the earlier three samples (P = 0.999). Consequently, once body proportions are included within the analysis, there is no change in lower limb diaphyseal hypertrophy across the two Middle Paleolithic samples (the late archaic human and Levantine early modern human ones) and the European Early Upper Paleolithic sample. It is only with the Middle Upper Paleolithic that one perceives a shift toward lower limb gracilization.
Figure 3.

Bivariate plots of femoral midshaft cortical area (Upper) and polar moment of area (Lower) vs. femoral length adjusted for variation in body core laterality. Symbols are as in Fig. 1.
At the same time, there is a clear shift between these late archaic and early modern humans in femoral diaphyseal cross-sectional shape (Fig. 4). The late archaic sample exhibits the pattern characteristic of all archaic Homo since 1.8 millions of years B.P. of having subcircular femoral midshafts with variable development of the linea aspera but lacking a pilaster (17, 19–21). In contrast, the majority of the early modern humans (all except some of the Middle Upper Paleolithic specimens) have femoral midshafts with prominent pilasters and the antero-posterior diameter greater than the medio-lateral one (17, 22). Even though details of the cross-sectional shapes may indicate contrasting developmental baselines for these two hominid grades, the differences in shape also may have biomechanical implications for lower limb loading patterns (as opposed to merely levels of habitual loading). It therefore is potentially profitable to evaluate the distributions of the antero-posterior (Iap) and medio-lateral (Iml) second moments of area of these femoral midshafts.
Figure 4.

Midshaft cross sections of late archaic and early modern human femora.
The plot of Iap vs. Iml (Fig. 5) reflects the subperiosteal pattern previously observed, namely the late archaic humans have highly significantly rounder femoral midshafts than the early modern human ones (P < 0.001), whereas the early modern human samples remain insignificantly different from each other (P = 0.683). However, if the individual second moments of area are plotted against femoral length adjusted for patterns of body laterality (Fig. 5), the pattern changes. In the plot of Iap vs. adjusted femoral length, the samples are not significantly different (P = 0.535), even though the late archaic humans are on average slightly less structurally reinforced in this plane than are the early modern human ones.
Figure 5.

Bivariate plots of femoral midshaft antero-posterior vs. medio-lateral second moments of area (Top), antero-posterior second moment of area vs. adjusted femoral length (Middle), and medio-lateral second moment of area vs. adjusted femoral length (Bottom). Symbols are as in Fig. 1.
In contrast to both the polar moment of area and Iap vs. adjusted femoral length, Iml vs. adjusted femoral length provides a highly significant difference between the late archaic and early modern human samples (P < 0.001) but no real contrasts across the latter samples (P = 0.669). These data indicate residual increased medio-lateral reinforcement of the femoral midshaft among the late archaic humans, even after adjustment for body laterality.
Discussion
Humeral Robusticity and Manipulation.
The pattern of changes in humeral diaphyseal robusticity is in good correlation with several other human biological indicators of the degree to which the human anatomy was directly used in manipulative behavior during the Late Pleistocene, despite contrasts with current interpretations of the associated Paleolithic archeological record.
Between late archaic humans and all of the considered early modern humans, these biological reflections of manipulative behavior include: marked decrease in the use of the anterior dentition for holding and processing objects, decrease in the robusticity of the more caudal cervical vertebral spinous processes, decrease in the size and rugosity of the proximal humeral insertions of the thoraco-humeral musculature, changes in orientation of the radial tuberosity reflecting biceps brachii moment arms, decrease in radial diaphyseal curvature related to pronation, marked reduction of palmar carpal tuberosities related to the size and mechanical advantages of both extrinsic and intrinsic hand musculature, changes in pollical phalangeal proportions related to pollical flexor muscle mechanical advantages, reduction of the crests for the pollical and digiti minimi opponens muscles, and decrease in the breadth of the pollical and middle phalangeal apical tufts (22–27). Although the degree of developmental plasticity of many of these shifts remains uncertain, all of them can be related biomechanically or attritionally to a reduction in the habitual load levels experienced by the upper limb and dentition for manipulative behavior. Humeral diaphyseal robusticity, which is developmentally highly plastic, patterns closely with these other paleontological indicators.
At the same time, there are archeological reflections of significant shifts of manipulative behavior, all related to technology, that correspond generally to these changes between the late archaic human and two Upper Paleolithic samples. These shifts involve increasing leptolithic exploitation of lithic raw materials, combined with the elaboration of the use of organic raw materials (especially bone and antler) into more effective composite technologies. This behavior is seen to some extent in the Early Upper Paleolithic, and it became far more standardized and elaborated through the Middle Upper Paleolithic (28–30). In addition, the Middle Upper Paleolithic saw the introduction of both ceramic and textile technologies (31, 32).
The anomaly remains the Levantine early modern humans. Their associated Middle Paleolithic archeological record is indistinguishable from that of late archaic humans, given normal variation within the Middle Paleolithic (33). The skeletal contrasts are unlikely to be merely stochastic, given both the multiple biological reflections of a contrast in manipulative behavior and the plasticity through the life cycle of several of these reflections, including diaphyseal hypertrophy, muscular attachment areas, and dental attrition. It is more likely that the behavioral dichotomy is not reflected in the currently analyzed aspects of the associated archeological record.
Femoral Robusticity and Locomotion.
Despite some previous assessments inferring a significant decrease in hominid locomotor levels with the emergence of modern humans, these data make it clear that any reduction in lower limb hypertrophy related to habitual adult locomotor levels is after ca. 30 kyr B.P. It therefore follows the establishment of early modern humans across the Old World. Other paleontological reflections of locomotor levels across these groups are more ambiguous in their implications. Gluteal tuberosity breadths change little between late archaic and Early Upper Paleolithic humans, but reduce in both Levantine Middle Paleolithic and Eurasian Middle Upper Paleolithic early modern human samples (23). Quadriceps femoris moment arms reduce uniformly between late archaic and early modern humans, as does pedal phalangeal diaphyseal hypertrophy (23, 34). However, femoral neck shaft angles (inversely reflecting locomotor levels during development) increase markedly in the Levantine early modern sample, less so in the Early Upper Paleolithic sample, but then decrease in the Middle Upper Paleolithic sample (24, 35).
Human foraging locomotor behavior is complex, because the resultant cumulative loads on the lower limb include distances traveled, intensity of activity, and levels and patterns of burden carrying. The archeological reflections of this behavior generally are ambiguous, because there is little necessary correlation between the distances materials have moved and the time, level and/or number of individual locomotor events responsible for the discard patterns on the prehistoric landscape. It is possible that the suggestions of increased social and hence task-related organizational complexity seen in the Middle Upper Paleolithic are functionally correlated with the shift in femoral robusticity, but any connection is likely to be very indirect and difficult to confirm.
The shift between late archaic and early modern humans in medio-lateral diaphyseal reinforcement is difficult to interpret. It is possible that it reflects decreases in pelvic and proximal femoral breadths relative to bone length, less pronounced than those between Early Pleistocene archaic Homo and recent humans (36) but still reflecting proportionately smaller bi-acetabular breadths and femoral biomechanical neck lengths. The alternative interpretation would be that there was a reduction in habitual medio-lateral loading of the lower limb during locomotion, an interpretation that other aspects of lower limb morphology may, but do not necessarily, support (34).
Summary
A consideration of the patterns of proximal appendicular diaphyseal cortical bone hypertrophy in Late Pleistocene Homo indicates a mosaic pattern of reduction in habitual load levels. There is a consistent contrast between late archaic and Middle Upper Paleolithic humans indicating a significant reduction in the habitual loads on both the upper and lower limbs. However, in the upper limb the greatest reduction is among the Levantine Middle Paleolithic-associated early modern humans, with the European Early Upper Paleolithic early modern humans exhibiting an intermediate level of hypertrophy. In the lower limb, in contrast, there is no overall difference in structural rigidity among the pre-30 kyr B.P. Late Pleistocene Homo samples. These morphologically plastic reflections of activity levels and patterns correlate well with other appendicular indications of manipulative and locomotor behaviors, but they are more difficult to correlate with the behaviorally often ambiguous Paleolithic archeological record. They nonetheless provide further insight into the nature, if not the genealogical history, of the evolutionary emergence of early modern humans.
Acknowledgments
I am grateful to the many curators who have permitted analysis of human remains in their care, to C.B. Ruff and S.E. Churchill for continued discussions and insight into these evolutionary processes, to T. Kimura, S.E. Churchill, and B. Holt for unpublished data, and to the L.S.B. Leakey Foundation, National Science Foundation, Wenner-Gren Foundation, and Centre National de la Recherche Scientifique for support.
ABBREVIATIONS
- kyr
thousands of years
- BIB
bi-iliac breadth
- FL
femoral length
References
- 1.Ruff C B, Trinkaus E, Walker A, Larsen C S. Am J Phys Anthropol. 1993;91:21–53. doi: 10.1002/ajpa.1330910103. [DOI] [PubMed] [Google Scholar]
- 2.Trinkaus E, Churchill S E, Ruff C B. Am J Phys Anthropol. 1994;93:1–34. doi: 10.1002/ajpa.1330930102. [DOI] [PubMed] [Google Scholar]
- 3.Ruff C B, Walker A, Trinkaus E. Am J Phys Anthropol. 1994;93:35–54. doi: 10.1002/ajpa.1330930103. [DOI] [PubMed] [Google Scholar]
- 4.Churchill S E. Am J Phys Anthropol. 1996;100:559–583. doi: 10.1002/(SICI)1096-8644(199608)100:4<559::AID-AJPA9>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 5.Trinkaus E. Anthropologie. 1996;34:101–110. [Google Scholar]
- 6.Lanyon L E. In: Bone in Clinical Orthropaedics. Sumner-Smith G, editor. Philadelphia: Saunders; 1982. pp. 273–304. [Google Scholar]
- 7.Meade J B. In: Bone Mechanics. Cowin S C, editor. Boca Raton, FL: CRC; 1989. pp. 211–251. [Google Scholar]
- 8.Krings M, Stone A, Schmitz R W, Krainitzki H, Stoneking M, Pääbo S. Cell. 1997;90:19–30. doi: 10.1016/s0092-8674(00)80310-4. [DOI] [PubMed] [Google Scholar]
- 9.Trinkaus E. In: Aspects of Human Evolution. Stringer C B, editor. London: Taylor & Francis; 1981. pp. 187–224. [Google Scholar]
- 10.Holliday T W. Ph.D. thesis. University of New Mexico; 1995. [Google Scholar]
- 11.Rabinovich R, Tchernov E. In: Archaeozoology of the Near East II. Buitenhuis H, Uerpmann H P, editors. Leiden, The Netherlands: Backhuys; 1995. pp. 5–44. [Google Scholar]
- 12.Nagurka M L, Hayes W C. J Biomech. 1980;13:59–64. doi: 10.1016/0021-9290(80)90008-1. [DOI] [PubMed] [Google Scholar]
- 13.Eschman P N. SLCOMM, Version 1.6. Albuquerque, NM: Eschman Archeological Services; 1992. [Google Scholar]
- 14.Ruff C B. J Hum Evol. 1991;21:81–105. [Google Scholar]
- 15.Ruff C B. Yearb Phys Anthropol. 1994;37:65–107. [Google Scholar]
- 16.Holliday T W. J Hum Evol. 1997;32:423–447. doi: 10.1006/jhev.1996.0111. [DOI] [PubMed] [Google Scholar]
- 17.Trinkaus E. Z Morphol Anthropol. 1976;67:291–319. [PubMed] [Google Scholar]
- 18.Hambücken A. Bull Mém Soc Anthropol Paris. 1995;7:37–47. [Google Scholar]
- 19.Weidenreich F. Palaeontol Sinica. 1941;5D:1–82. [Google Scholar]
- 20.Day M H. Nature (London) 1971;232:383–387. doi: 10.1038/232383a0. [DOI] [PubMed] [Google Scholar]
- 21.Grine F E, Jungers W L, Tobias P V, Pearson O M. Am J Phys Anthropol. 1995;97:151–185. doi: 10.1002/ajpa.1330970207. [DOI] [PubMed] [Google Scholar]
- 22.Trinkaus E. In: The Evolution and Dispersal of Modern Humans in Asia. Akazawa T, Aoki K, Kimura T, editors. Tokyo: Hokusen-Sha; 1992. pp. 277–294. [Google Scholar]
- 23.Trinkaus E. The Shanidar Neandertals. New York: Academic; 1983. [Google Scholar]
- 24.Trinkaus E, Jelínek J. J Hum Evol. 1997;33:33–82. doi: 10.1006/jhev.1997.0142. [DOI] [PubMed] [Google Scholar]
- 25.Trinkaus E, Churchill S E. Am J Phys Anthropol. 1988;75:15–21. doi: 10.1002/ajpa.1330750103. [DOI] [PubMed] [Google Scholar]
- 26.Trinkaus E, Villemeur I. Am J Phys Anthropol. 1991;84:249–260. doi: 10.1002/ajpa.1330840303. [DOI] [PubMed] [Google Scholar]
- 27.Villemeur I. La Main des Néandertaliens. Paris: CNRS Editions; 1994. [Google Scholar]
- 28.Hahn J. Aurignacien: Der Ältere Jungpaläolithikum in Mittel- und Osteuropa. Cologne, Germany: Böhlau; 1977. [Google Scholar]
- 29.Knecht H, Pike-Tay A, White R, editors. Before Lascaux. Boca Raton, FL: CRC; 1993. [Google Scholar]
- 30.Svoboda J, Ložek V, Vlček E. Hunters Between East and West. New York: Plenum; 1996. [Google Scholar]
- 31.Vandiver P B, Soffer O, Klíma B, Svoboda J. Science. 1989;246:1002–1008. doi: 10.1126/science.246.4933.1002. [DOI] [PubMed] [Google Scholar]
- 32.Adovasio J M, Hyland D C, Soffer O. In: Pavlov I–Northwest. Svoboda J, Sǩrdla P, editors. Brno: Akademie věd České Republiky; 1997. pp. 403–424. [Google Scholar]
- 33.Shea J J. In: The Human Revolution. Mellars P, Stringer C B, editors. Princeton: Princeton Univ. Press; 1989. pp. 611–625. [Google Scholar]
- 34.Trinkaus E, Hilton C E. J Hum Evol. 1996;30:399–425. [Google Scholar]
- 35.Trinkaus E. J Hum Evol. 1993;25:393–416. [Google Scholar]
- 36.Ruff C B. Am J Phys Anthropol. 1995;98:527–574. doi: 10.1002/ajpa.1330980412. [DOI] [PubMed] [Google Scholar]