
Figure 7. Models for cell cycle regulation of DSB repair and checkpoint activation.
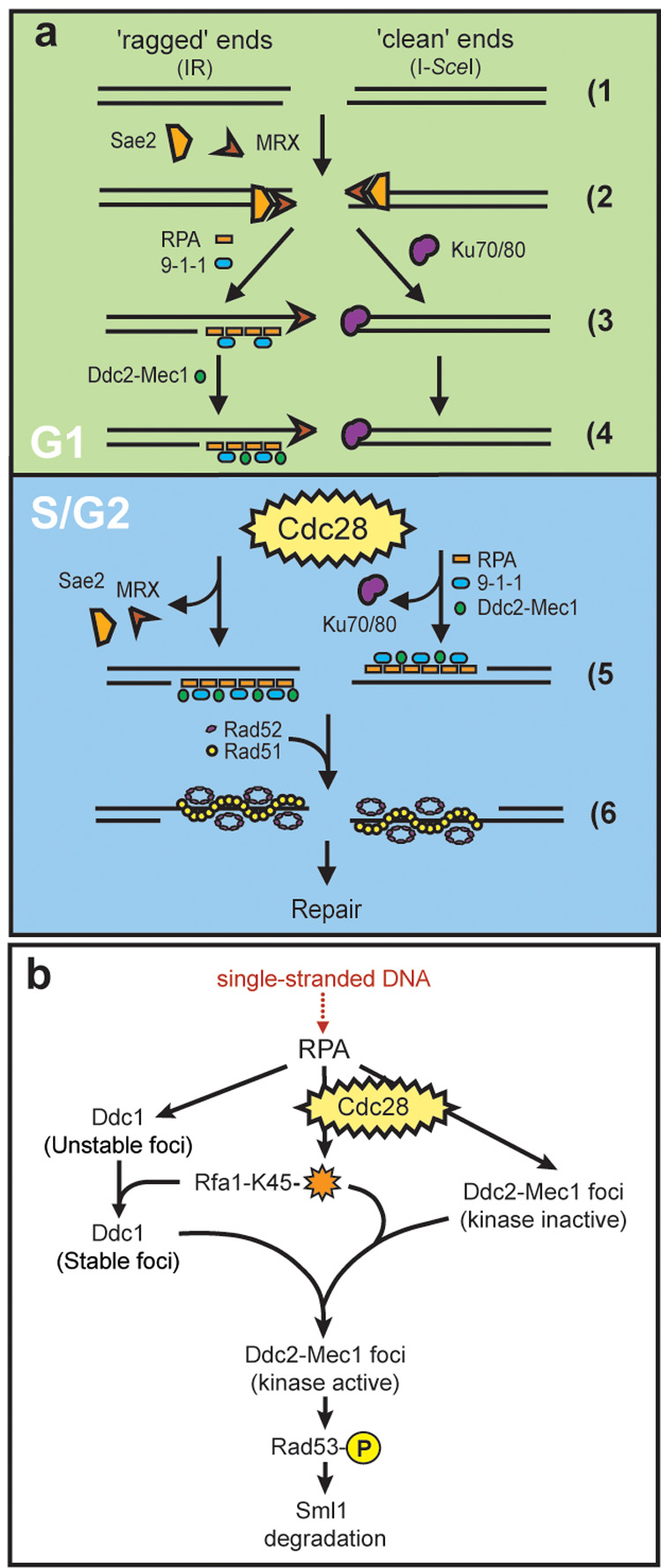
(a) Cell cycle regulation of DSB repair. When a DSB is generated (1), ends may be either ‘ragged’ as those produced following exposure to IR (left) or ‘clean’ such as those resulting from an endonuclease-mediated cleavage (right). The MRX complex recognizes both kinds of ends (2) while during G1, clean ends are preferentially bound by Yku70/Yku80 (3). Once Yku70/Yku80 is bound, the ends are not released until ligation is complete or until the cell has entered into S phase (4, 5). Upon entering S phase, Yku70/Yku80 dissociates from the ends, freeing them for digestion by nucleases resulting in ssDNA. RPA binds the ssDNA, independently recruiting the 9-1-1 (Ddc1-Mec3-Rad17) and Ddc2-Mec1 checkpoint complexes (5, right). ‘Ragged’ DNA ends, on the other hand, are not bound by Yku70/Yku80 during G1. Instead, these free ends are processed by one or more nucleases into 3’ ssDNA tails, recognized and bound by RPA. RPA then recruits Ddc1-Mec3-Rad17, which in turn is required for Ddc2-Mec1 focus formation in G1. (4, left). The homologous recombination machinery, here shown as Rad52 and Rad51, cannot be recruited however, until the cells have entered into S phase (6).
(b) Ddc2-Mec1 checkpoint activation. In G1 cells, 9-1-1(Ddc1-Mec3-Rad17) is absolutely required for recruitment of Ddc2 into foci and subsequent Rad53 phosphorylation. In S/G2, Cdc28 activity and RPA modification on Lysine 45 (orange star) act in conjunction with the 9-1-1 complex to promote Ddc2 focus formation and downstream checkpoint events. See discussion for details.