

Abstract
Telomerase is an essential enzyme that maintains telomeres on eukaryotic chromosomes. In mammals, telomerase is required for the lifelong proliferative capacity of normal regenerative and reproductive tissues and for sustained growth in a dedifferentiated state. Although the importance of telomeres was first elucidated in plants 60 years ago, little is known about the role of telomeres and telomerase in plant growth and development. Here we report the cloning and characterization of the Arabidopsis telomerase reverse transcriptase (TERT) gene, AtTERT. AtTERT is predicted to encode a highly basic protein of 131 kDa that harbors the reverse transcriptase and telomerase-specific motifs common to all known TERT proteins. AtTERT mRNA is 10–20 times more abundant in callus, which has high levels of telomerase activity, versus leaves, which contain no detectable telomerase. Plants homozygous for a transfer DNA insertion into the AtTERT gene lack telomerase activity, confirming the identity and function of this gene. Because telomeres in wild-type Arabidopsis are short, the discovery that telomerase-null plants are viable for at least two generations was unexpected. In the absence of telomerase, telomeres decline by approximately 500 bp per generation, a rate 10 times slower than seen in telomerase-deficient mice. This gradual loss of telomeric DNA may reflect a reduced rate of nucleotide depletion per round of DNA replication, or the requirement for fewer cell divisions per organismal generation. Nevertheless, progressive telomere shortening in the mutants, however slow, ultimately should be lethal.
Keywords: transfer DNA, telomeres, plant, callus
The groundbreaking studies of Barbara McClintock (1, 2) and Hermann Muller (3) demonstrated that genome stability depends on the integrity of the telomere complex at the ends of eukaryotic chromosomes. Although alternative strategies have been reported (4), telomere synthesis by telomerase is the primary mechanism for sustaining chromosome ends in eukaryotes. Telomeres and their maintenance by telomerase comprise a biological clock that influences cellular lifespan in mammals (5). Telomerase expression is confined primarily to the germ line and permanently regenerating tissues of the adult soma. In other cells, telomerase is turned off and telomeres progressively shorten with each division. Once telomeres shorten below a critical length, a DNA damage checkpoint is activated, leading to cellular senescence and death. Telomerase is reactivated in about 85% of all human tumors and telomere function is maintained indefinitely (6).
Telomerase is a ribonucleoprotein reverse transcriptase (7). The RNA subunit contains a templating sequence complementary to the G-rich strand of the telomere, whereas the telomerase reverse transcriptase (TERT) harbors the catalytic activity for telomere repeat synthesis. Characterization of TERT subunits from a variety of protozoa, yeasts, and mammals has revealed several distinct reverse transcriptase motifs that comprise the polymerase active site as well as a telomerase-specific motif (8–12).
TERT mRNA levels parallel telomerase activity in both mice and humans (9, 13–15). Further regulation is achieved posttranscriptionally by alternative splicing (16, 17) and perhaps by differential protein stability (18). Ectopic expression of TERT in telomerase-negative tissue culture cells activates telomerase and leads to telomere lengthening and increased lifespan (19–22). Thus, TERT is a key regulator of the telomere clock in mammals (23).
Characterization of telomerase-null mice established a critical role for this enzyme in mammalian growth and development. Phenotypically normal for the first five generations, telomerase-null mice exhibit inexorable telomere shortening at a rate of approximately 5 kbp per generation (24). In the sixth generation, the proliferative capacity of self-renewing tissues decreases, abnormal karyotypes are observed, and the male reproductive system collapses (25). The delay in detecting a detrimental phenotype in Mus musculus is attributed to the extremely long telomeres in this species (40–150 kbp) (26, 27), which must be eroded before chromosome ends become uncapped.
Plants have a more plastic pattern of development than animals. Although this plasticity might suggest a more flexible profile of telomerase regulation, telomerase expression actually is tightly controlled in plants and closely associated with cell proliferation and dedifferentiation (28). Telomerase is not detectable in vegetative tissues, but is highly expressed in reproductive (refs. 29–33; M.S.F. and D.E.S., unpublished work) and dedifferentiated tissues, including crown gall tumors of tobacco (E. Grace, T.D.M., and D.E.S., unpublished results) and callus cultures of barley (34), Arabidopsis (35), carrot, and soybean (29).
We are using Arabidopsis thaliana as a model to define the role of telomerase in plant growth and development and to address fundamental questions concerning telomere function in higher eukaryotes. One advantage of Arabidopsis over mouse is that it contains significantly shorter telomeres (2–4 kb) (36) and hence perturbations in the telomere maintenance machinery should be readily detected as proportionally large changes in a short sequence. Here we report the cloning of the Arabidopsis TERT gene (AtTERT) and demonstrate that its expression is an accurate marker for telomerase activity. We also show that a homozygous transfer DNA (T-DNA) insertion in the AtTERT gene abolishes telomerase activity. Despite the short telomeres in Arabidopsis, the telomerase deficiency is not immediately lethal and mutant plants survive for at least two generations.
Materials and Methods
Plant Growth and Callus Induction.
A. thaliana (Columbia ecotype) was grown at 21°C in an environmental growth chamber under a 24-hr photoperiod. Callus was initiated by placing excised hypocotyls of 4-day-old plants on solid MS medium (37) supplemented with 2 mg 2,4-dichlorophenoxyacetic acid and 0.05 mg kinetin per liter. Callus was maintained in the dark at 25°C on the same medium.
Preparation of Telomerase Extracts and Telomere Repeat Amplification Protocol (TRAP) Assays.
Extracts were prepared from rosette and cauline leaves, inflorescence bolts, flowers, and siliques as described (29). Telomerase was detected by a modified version of the TRAP (29). Products were resolved on 6% sequencing gels that were dried and subjected to autoradiography.
Cloning and Sequence Analysis of the AtTERT Genomic Clone and cDNA.
The peptide sequence of human TERT was used to query all Arabidopsis DNA sequences in GenBank by the tblastn algorithm (38). A 624-bp region at the end of one bacterial artificial chromosome clone (T17O4) was identified as encoding a protein with a high degree of similarity to human TERT. Primers 1235 (CTTCATTGCAGCCAACAGAAA) and 1233 (GACTACACAAGGTCTGCCTCA) were used to generate a PCR probe to identify a genomic clone from an A. thaliana (Landsberg erecta) lambda library, kindly provided by Terry Thomas (Texas A & M University). Six- and nine-kilobase SalI fragments were subcloned into pGEM-3Zf, and both strands of the gene were sequenced. Exon/intron junctions were predicted by the splicepredictor program (39). The position of introns was verified by sequence analysis of AtTERT cDNA. Three segments of AtTERT mRNA from callus were amplified by reverse transcription–PCR (RT-PCR) using the Access RT-PCR kit (Promega). The products were cloned into the pBAD vector (Invitrogen) and sequenced. Multiple sequence alignments were performed by the program clustal w (version 1.74) using a blosum 62 matrix under default parameters (40).
RNA Extraction and RT-PCR Analysis of AtTERT mRNA.
Arabidopsis leaf and callus mRNA was isolated by using the FastTrack mRNA isolation kit (Invitrogen). RT-PCRs were carried out with 100 ng of mRNA by using the Assess RT-PCR system (Promega) following the manufacturer's instructions. Reactions were performed with three sets of primers: 6 (GGACATATCCATCAAGGGC) and 7 (GGAAGCTGTATTGCACG); 5 (GCCCTTGATGGATATGTCC) and 48 (CCAACTGCAGCATGTTGTTC); 10 (GTCGTTCCGGACTTCAATGC) and 11 (CTGCTCTGATTCAAAGCTCC). RT was conducted in a RoboCycler (Stratagene) for 45 min at 48°C and then the enzyme was inactivated by incubation at 96°C for 2 min. PCR was carried out for 20 cycles under the following conditions: 94°C for 45 sec, 63°C for 45 sec, 72°C for 1 min followed by a final elongation period of 7 min at 72°C. A 10-μl aliquot of the PCR was resolved on a 1.5% agarose gel and subjected to Southern blot analysis using probes generated from each PCR product. As a quantitation control, RT-PCR was performed on leaf and callus mRNA with primers specific for the cystosolic glyceraldehyde-3-phosphate dehydrogenase gene (forward: GACCTTACTGTCAGACTCGAG and reverse: CGGTGTATCCAAGGATTCCCT). Signals were quantitated on a STORM PhosphorImager (Molecular Dynamics).
Identification of an AtTERT Null Plant.
The PCR-based method of McKinney et al. (41) was used to screen T-DNA insertion lines for a disruption of the AtTERT gene. Pooled DNA samples from 12,000 T-DNA insertion lines (CD5–7 and CD6–7) were obtained from the Arabidopsis Biological Resource Center (ABRC). DNA was provided in 12 pools of 1,000 individual lines. PCR was performed in a Stratagene RoboCycler using 50-μl reactions containing 25 ng of pooled DNA, 1 unit of Taq polymerase (Promega), 0.1 mM dNTPs, and 250 μM of primers for the left or right border of the T-DNA (provided by ABRC) along with primer 1 (GTTTATGTGCTAAGCGACCTGCGTTTGCG) or primer 4 (GAATACATCCGCAACCACAAAGTACAAAG) derived from the AtTERT sequence. PCR conditions were 3 min at 94°C, followed by 40 cycles (94°C, 45 sec; 65°C, 1 min; 72°C, 2 min) and a final extension at 72°C for 7 min. Southern blot analysis was used to assess the validity of PCR products.
A plant heterozygous for the T-DNA disruption was self-pollinated and its progeny were screened for a mutation in both alleles by PCR. DNA was extracted from young leaves of individual progeny (42) and 1 μl was added to a PCR mixture containing 1.5 mM MgCl2, 200 μM dNTPs, 800 μM primer 6, 500 μM primer 7 primer, 400 μM T-DNA LB (CD6) primer, and 1 unit of Taq polymerase (Promega). PCR conditions were 2 min at 94°C, then 35 cycles (45 sec at 94°C, 45 sec at 60°C, and 1 min at 72°C) and a final extension of 5 min at 72°C. A 457-bp product was generated from the wild-type allele, and a 729-bp product was generated with the disrupted allele. Plants with a homozygous disruption were self-pollinated to obtain subsequent generations.
DNA Extraction and Terminal Restriction Fragment (TRF) Analysis.
DNA was extracted according to Li and Chory (43) from rosette leaves 3 weeks after germination or from stems with inflorescences and siliques 5 weeks after germination. A 1-μg aliquot of DNA from each sample was cleaved with Tru9I (Promega), resolved on a 1% agarose gel, and subjected to Southern blot analysis as described (32). [32P] 5′ end-labeled (T3AG3)4 was used as a probe. Radioactive signals were scanned by a STORM PhosphorImager (Molecular Dynamics), and the data were analyzed by imagequant software (Molecular Dynamics).
Results
Identification of the AtTERT Gene.
The gene encoding the TERT from Arabidopsis was identified by using the human TERT protein (9) as the query sequence in a blast search of the Arabidopsis database. We found a short region at the end of a bacterial artificial chromosome (BAC T17O4) encoding a protein with strong similarity to conserved RT domains in human and yeast TERTs. A probe generated from the BAC was used to retrieve the corresponding region from a lambda genomic library.
A 5,618-bp region of genomic DNA spanning the putative TERT gene was sequenced (GenBank accession no. AF172097). Intron organization was predicted by the splicepredictor program (39) and confirmed by sequence analysis of three cDNA clones (Fig. 1A). The gene harbors 11 introns and encodes a single basic protein (pI = 9.62) of 131 kDa. The predicted peptide sequence is 24% identical and 62% similar to the human TERT and carries all of the motifs conserved in other TERTs (Fig. 1B). Amino acid residues in the T motif and RT motifs 1, 2, A, and C shown to be essential for telomerase activity in yeast (8), and human TERTs (44, 45) also are conserved in the Arabidopsis protein. Based on its similarity to other TERT genes, we designated the Arabidopsis gene, AtTERT. Southern analysis indicated that AtTERT is single-copy in the Arabidopsis genome (data not shown).
Figure 1.

Identification of the AtTERT gene. (A) Schematic representation of the AtTERT gene. Open boxes represent exons, black lines, introns, and gray boxes, conserved TERT motifs. The position of PCR primers (arrows) and a T-DNA insertion are indicated. (B) Alignment of AtTERT with TERT proteins from human (hTERT) (9), mouse (mTERT) (18), Schizosaccharomyces pombe (SpTERT) (9), Saccharomyces cerevisiae (ScTERT) (8), Euplotes aediculatus (EaTERT) (8), Oxytricha trifallax (OtTERT) (12), and Tetrahymena thermophila (TtTERT) (11, 12).
Expression of AtTERT mRNA Correlates with Telomerase Activity.
We tested whether the AtTERT gene expression correlates with telomerase activity. TRAP assays were performed on various vegetative and reproductive organs from Arabidopsis. As shown for other plants (29, 30, 32, 33), telomerase was undetectable in leaves (Fig. 2A, lanes 2 and 3), but strongly expressed in cells with high proliferation capacity (reproductive organs, seeds, and callus) (Fig. 2A, lanes 1, 4 and 7). To determine whether AtTERT mRNA levels correlate with enzyme activity, RT-PCR experiments were conducted with callus (telomerase positive) and rosette leaf (telomerase negative) RNA samples. Primer pair 6 and 7 does not span an intron and is expected to generate a 457-bp product, whereas primer set 10 and 11 flanks an intron and should generate a 237-bp product in the spliced mRNA (Fig. 1A). Although the PCRs yield products of the predicted size in both callus and leaf mRNA samples, the level was 10- to 20-fold higher in callus (Fig. 2B). In telomerase-negative human cells, human TERT mRNA is alternatively spliced to eliminate an essential RT motif in the protein (16, 17, 46). The ninth intron in the AtTERT gene lies between motifs A and B near but not precisely in the same position as the alternatively spliced intron in human TERT. To test whether this intron is alternatively spliced in AtTERT mRNA, RT-PCR experiments were performed with primers 5 and 48 (Fig. 1A). Although the intensity of the products was reduced in leaves relative to callus, the expected 606-bp product was generated in both reactions, indicating that this intron spliced in the same manner in both tissues (Fig. 2B).
Figure 2.

AtTERT mRNA levels correlate with telomerase activity. (A) Telomerase regulation in Arabidopsis. TRAP results from different organs. Elongation ladders correspond to the addition of TTTAGGG repeats. Silque (seed pod); inflor. (inflorescence) bolt. (B) RT-PCR products from callus (C) and leaf (L) mRNA. Primer pairs used to generate AtTERT products (*) are indicated. Bottom band represents the glyceraldehyde-3-phosphate dehydrogenase quantitation control.
Disruption of the AtTERT Gene by T-DNA Insertion Eliminates Telomerase Activity.
The identity and function of the AtTERT gene were confirmed by examining a plant with this gene disrupted by a T-DNA insertion. To obtain an AtTERT-deficient plant, we screened 12,000 independently transformed Arabidopsis lines by using a PCR-based procedure (reviewed in ref. 47). A plant was identified harboring a T-DNA inserted in the ninth exon of AtTERT, 228 nt upstream of the region encoding the T-motif (Fig. 1A).
PCR with primers designed to detect the insertion or a wild-type exon showed that the original plant was heterozygous for the T-DNA insertion. This plant was allowed to self-pollinate, and its progeny were screened by PCR to find plants with a homozygous disruption of AtTERT. The T-DNA insert was inherited in a Mendelian manner, and one-fourth of the progeny (6/23) were homozygous for the AtTERT disruption. TRAP assays conducted on floral buds from the knockout lines confirmed that AtTERT is a single-copy gene and is essential for telomerase activity in Arabidopsis (Fig. 3, lane 6).
Figure 3.
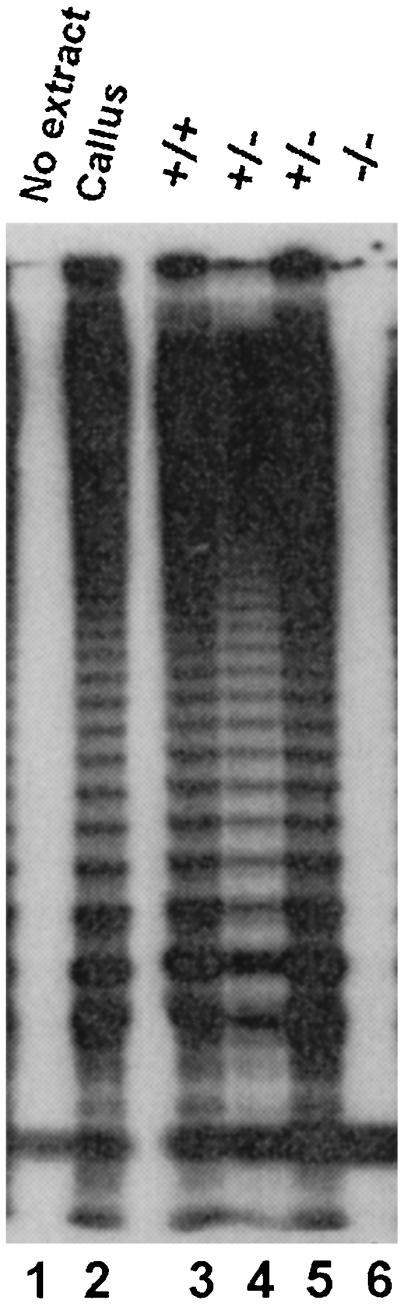
Plants with a homozygous disruption in the AtTERT gene lack telomerase. TRAP assays were performed on the floral buds from plants that are wild type (+/+), heterozygous (+/−), or homozygous (−/−) for the T-DNA disruption in AtTERT.
Telomere Shortening in Telomerase-Deficient Plants.
Plants with an AtTERT disruption do not exhibit obvious phenotypic changes in the first (G1) or second (G2) generation. Because a temporal lag between telomere loss and diminished cell viability is characteristic of telomerase-null yeast (48, 49) and mice (25), we investigated the status of telomeres in mutant plants using TRF analysis of DNA from leaves, floral buds, and siliques (seed pods). Tru9I digestion of DNA from wild-type plants and plants heterozygous for the AtTERT disruption generated a classic telomere smear between 2 and 4 kbp (Fig. 4A, lanes 1, 2, and 5). Interestingly, the size of the telomeric DNA tracts in rosette leaves (Fig. 4A, lane 1) was the same as in floral buds and sliques (Fig. 4A, lane 2), demonstrating that Arabidopsis telomeres do not undergo the dramatic shortening characteristic of telomeres in barley development (34) and aging mammalian cells (50).
Figure 4.

Progressive telomere shortening in telomerase-deficient plants. (A) TRF analysis. Shown are results with Tru9I digestion of DNA from rosette leaves (lane 1) or floral buds and siliques (lane 2) of wild-type plants and rosette leaves of heterozygous plants (lane 5). Results with rosette leaf DNA from first-generation (G1) and second-generation (G2) plants homozygous AtTERT disruption are shown in lanes 3 and 4. (B) Quantitative analysis of TRF data from wild-type (wt) and G2 homozygous mutant plants digested with Tru9I or HaeIII (C) are shown. * indicate interstitial DNA with homology to the telomeric repeat. The rate of telomere shortening was calculated by comparing the size of the smallest telomere fragments in each generation of the mutants with telomeres from wild-type plants.
DNA samples from homozygous mutant plants exhibited a strikingly different profile. Starting in G1, the telomeric smear in AtTERT-deficient plants was replaced by several sharp bands (Fig. 4A, lane 3). By G2 at least 11 discrete telomeric fragments were visible (Fig. 4A, lane 4; Fig. 4B). The Tru9I restriction enzyme (recognition sequence: TTAA) cleaves degenerate telomere repeat sequences adjacent to the terminal T3AG3 tract (36). We tested whether the discrete banding profile would be evident in DNA samples treated with enzymes that cut at more internal sites on the chromosome. A similar banding profile was obtained with HaeIII (Fig. 4C) or AluI digestion (data not shown). Several of the higher molecular weight telomeric bands in G1 and G2 homozygous mutants comigrated with the smear of wild-type DNA and one band actually appeared to increase in size in G2. Overall, however, the profile showed a dramatic downward shift from G1 to G2 (Fig. 4A, lanes 3 and 4). PhosphorImager analysis of the smallest telomere fragments indicated that the telomeres were shortening at a rate of approximately 500 bp per generation (Fig. 4 B and C). This rate of depletion is 10-fold slower than in telomerase-null mice (24).
Telomerase Expression Is Not a Prerequisite for Dedifferentiation.
Human cells stop dividing in culture within approximately 50 cell divisions, and immortalized lines are generated only if senescence is bypassed by oncogenic viral infection or multiple genetic mutations (51). In contrast, most cells in cultured plant explants achieve immortality. To investigate whether telomerase activation is a prerequisite for dedifferentiation and immortalization in plants, we asked whether callus could be generated from telomerase-null plants. In wild-type plants, callus is easily induced from Arabidopsis hypocotyls (see Materials and Methods). Callus was readily generated from G1 telomerase-null plants and is morphologically indistinguishable from wild-type callus (data not shown). TRAP assays confirmed that telomerase was not reactivated (data not shown). The callus has been successfully cultivated for 4 months, and future studies will reveal whether it is dedifferentiated but mortal, a novel combination of traits for plant cells.
Discussion
Plant and animal lineages evolved independently before multicellularization and this divergence is reflected in their distinctive patterns of development. Expression of telomerase and, in particular, the TERT subunit of telomerase, is intimately tied to cell proliferation programs, dedifferentiation, and immortalization in mammals. To elucidate the role of telomerase in plant growth and development, we cloned the gene encoding the catalytic subunit of the Arabidopsis telomerase and confirmed its identity by demonstrating that plants with a T-DNA insertion in this gene lack telomerase activity. The structure of AtTERT is remarkably similar to other TERTs and like the mammalian counterparts, AtTERT expression is an accurate marker for enzyme activity. While this work was under review, Oguchi et al. (52) reported the cloning of AtTERT cDNA and provided further evidence that AtTERT expression correlates with telomerase activity.
It is not surprising that telomerase is active in reproduction in both plant and animal lineages where it helps ensure that progeny inherit a full complement of chromosomes. However, that both plants and animals independently evolved mechanisms to repress telomerase in most nonreproductive organs is noteworthy. It may be that constitutive expression of telomerase has an adverse impact on fitness. A nonadaptive model for the loss of expression in nonreproductive organs is also feasible. Stochastic inactivation of telomerase in nonreproductive organs probably has no deleterious consequences, and over time most evolutionary lineages may have lost the ability to express telomerase except in tissues where it is essential.
One interesting outcome of our study was the observation that telomerase expression is not a prerequisite for dedifferentiation in Arabidopsis. Likewise, fibroblasts derived from telomerase-deficient mice grow in culture beyond 500 population doublings and form tumors as efficiently as wild-type cells (24). Cytogenetic studies revealed telomere shortening occurs at different rates on individual mouse chromosomes, and some telomeres actually increase in length, implying that an alternative telomere maintenance mechanism (ALT) has been activated (53). In yeast (54) and probably in mammals (4), ALTs are mediated by recombination. Because plants induce recombination pathways at a higher frequency than animals (55, 56), ALTs may be readily activated in telomerase-null plants and callus. One piece of data supporting this idea is the observation that at least one of the telomeric bands from the mutants appears to increase in size in second-generation null plants (see below).
Given the short telomeres of Arabidopsis (57), it was unclear whether plants carrying a homozygous disruption of AtTERT would be viable. That these plants were recovered at expected frequencies in the progeny of the initial heterozygous plant indicates a lack of detrimental effect on gametogenesis and embryogenesis. These plants currently are growing in their third generation. Although the homozygous mutants display no obvious growth defects, TRF analysis demonstrated that their telomeres are significantly shorter than wild-type plants. Progressive telomere shortening in subsequent generations ultimately should lead to profound defects in growth and development, if telomerase action is essential for genome stability.
TRF experiments uncovered a remarkable qualitative change in the structure of telomeres in telomerase-null plants, which provides insight into the mechanism of telomere maintenance. Telomeres typically appear as a smear of DNA representing variations in telomere length at individual chromosome ends and between cells. Instead of a telomere smear, DNA samples from plants homozygous for the AtTERT disruption exhibited a series of discrete bands.
These bands could represent amplification of subtelomeric sequences, analogous to the accumulation of telomere-associated Y′ elements, an ALT that allows a small subset of yeast cells to survive without telomerase (54). Y′ elements contain stretches of telomeric DNA repeats, and accumulation of these internal telomeric repeats is thought to restore telomere function. Three lines of evidence argue against this mechanism in telomerase-null plants. First, the telomeric bands appear in the first generation of telomerase-deficient plants and completely replace the normal profile of telomeric DNA. The Mendelian inheritance of the T-DNA disruption indicates that telomerase-deficient gametes or embryos are not selected against, and hence the TRF pattern is not derived from rare cells that activated an ALT. Second, the Tru9I restriction enzyme cleaves within a tract of variant telomeric sequence adjacent to the perfect array of terminal T3AG3 repeats. Accordingly, telomeric smears should still be detectable in DNA samples from telomerase-null plants. Finally, the majority of telomeric fragments decrease in size in successive generations of the mutants, a finding consistent with erosion of terminal DNA tracts.
We postulate that the discrete telomeric bands represent individual chromosome ends. Arabidopsis embryos begin life as a single cell with 20 telomeres, each with a defined size. During embryogenesis and subsequent development of wild-type plants, telomerase extends the telomeric DNA tract whereas other activities such as incomplete DNA replication and possibly telomere-specific nucleases (58) shorten the tract. Together, these activities broaden the size distribution of all 20 telomeres, generating the typical smear on Southern blots. If telomere lengthening by telomerase is primarily responsible for generating size heterogeneity, then telomeres in telomerase-deficient plants should remain at a discrete, but shortened, size in all descendant cells. The striking homogeneity of the fragments implies that the rate of telomere shortening is the same for a given chromosome in every cell, a conclusion supported by fluorescence in situ hybridization analysis of cells derived from telomerase-deficient mouse embryos (54). When the Arabidopsis genome project is completed next year (59), specific probes for all the telomeres can be generated to test this model and to address whether ALTs are acting on particular chromosome ends.
At only 500 bp per generation, the rate of telomere loss in telomerase-null plants is 10 times less than the rate reported for telomerase-deficient mice (24). We envision two possible explanations for this discrepancy. First, fewer nucleotides may be lost from Arabidopsis telomeres per round of DNA replication. Arabidopsis may be similar to yeast wherein telomeres in telomerase-negative cells decline by only 2–4 bp per cell division (48) instead of the 50- to 100-bp loss in mammalian cells (50, 60). Second, there may be fewer cell generations per organismal generation in Arabidopsis than in mice. Approximately 50 cell generations are required per mouse generation (61, 62). Establishing the corresponding number for Arabidopsis is problematic because mitotic activity primarily is concentrated in a few meristematic zones, and cell division rates vary widely within a single meristem (63). Whatever the cause, the slow rate of telomeric DNA loss in conjunction with the global qualitative changes that occur in telomeres of telomerase-null mutants indicate that Arabidopsis will be a useful model for investigating telomere maintenance mechanisms in higher eukaryotes.
Acknowledgments
We thank Alan Pepper for supplying the T17O4 bacterial artificial chromosome clone, the Gene Technology Laboratory at Texas A&M for running DNA sequencing gels, the Arabidopsis Stock Center for supplying DNA and seeds from T-DNA lines, and Jeff Kapler and Craig Nessler for critically reading the manuscript. This work was funded by a grant from the Advanced Texas Research Program to D.E.S. (999902–141) and T.D.M. (010366–141).
Abbreviations
- TRAP
telomere repeat amplification protocol
- TERT
telomerase reverse transcriptase
- AtTERT
Arabidopsis TERT gene
- TRF
terminal restriction fragment
- T-DNA
transfer DNA
- RT-PCR
reverse transcription–PCR
- ALT
alternative telomere maintenance mechanism
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. AF172097).
References
- 1.McClintock B. Proc Natl Acad Sci USA. 1939;25:405–416. doi: 10.1073/pnas.25.8.405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.McClintock B. Genetics. 1941;26:234–282. doi: 10.1093/genetics/26.2.234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Muller H J. Collect Net. 1938;13:181–195. [Google Scholar]
- 4.Colgin L M, Reddel R R. Curr Opin Genet Dev. 1999;9:97–103. doi: 10.1016/s0959-437x(99)80014-8. [DOI] [PubMed] [Google Scholar]
- 5.Chiu C P, Harley C B. Proc Soc Exp Biol Med. 1997;214:99–106. doi: 10.3181/00379727-214-44075. [DOI] [PubMed] [Google Scholar]
- 6.Shay J W, Bacchetti S. Eur J Cancer. 1997;33:787–791. doi: 10.1016/S0959-8049(97)00062-2. [DOI] [PubMed] [Google Scholar]
- 7.Greider C W. In: Telomeres. Blackburn E H, Greider C W, editors. Plainview, NY: Cold Spring Harbor Lab. Press; 1995. pp. 35–68. [Google Scholar]
- 8.Lingner J, Hughes T R, Shevchenko A, Mann M, Lundblad V, Cech T R. Science. 1997;276:561–567. doi: 10.1126/science.276.5312.561. [DOI] [PubMed] [Google Scholar]
- 9.Nakamura T M, Morin G B, Chapman K B, Weinrich S L, Andrews W H, Lingner J, Harley C B, Cech T R. Science. 1997;277:955–959. doi: 10.1126/science.277.5328.955. [DOI] [PubMed] [Google Scholar]
- 10.Nakamura T M, Cooper J P, Cech T R. Science. 1998;282:493–496. doi: 10.1126/science.282.5388.493. [DOI] [PubMed] [Google Scholar]
- 11.Collins K, Gandhi L. Proc Natl Acad Sci USA. 1998;95:8485–8490. doi: 10.1073/pnas.95.15.8485. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Bryan T M, Sperger J M, Chapman K B, Cech T R. Proc Natl Acad Sci USA. 1998;95:8479–8484. doi: 10.1073/pnas.95.15.8479. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Meyerson M, Counter C M, Eaton E N, Ellisen L W, Steiner P, Caddle S D, Ziaugra L, Beijersbergen R L, Davidoff M J, Liu Q, et al. Cell. 1997;90:785–795. doi: 10.1016/s0092-8674(00)80538-3. [DOI] [PubMed] [Google Scholar]
- 14.Greenberg R A, Allsopp R C, Chin L, Morin G B, DePinho R A. Oncogene. 1998;16:1723–1730. doi: 10.1038/sj.onc.1201933. [DOI] [PubMed] [Google Scholar]
- 15.Takakura M, Kyo S, Kanaya T, Tanaka M, Inoue M. Cancer Res. 1998;58:1558–1561. [PubMed] [Google Scholar]
- 16.Kilian A, Bowtell D D L, Abud H E, Hime G R, Venter D J, Keese P K, Duncan E L, Reddel R R, Jefferson R A. Human Mol Gen. 1997;6:2011–2019. doi: 10.1093/hmg/6.12.2011. [DOI] [PubMed] [Google Scholar]
- 17.Ulaner G A, Hu J-F, Vu T H, Giudice L C, Hoffmann A R. Cancer Res. 1998;58:4168–4172. [PubMed] [Google Scholar]
- 18.Martin-Rivera L, Herrera E, Albar J P, Blasco M A. Proc Natl Acad Sci USA. 1998;95:10471–10476. doi: 10.1073/pnas.95.18.10471. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Bodnar A G, Ouellette M, Frolkis M, Holt S E, Chiu C P, Morin G B, Harley C B, Shay J W, Lichtsteiner S, Wright W E. Science. 1998;279:349–352. doi: 10.1126/science.279.5349.349. [DOI] [PubMed] [Google Scholar]
- 20.Nakayama J I, Tahara H, Tahara E, Saito M, Ito K, Nakamura H, Nakanishi T, Tahara E, Ide T, Ishikawa F. Nat Genet. 1998;18:65–68. doi: 10.1038/ng0198-65. [DOI] [PubMed] [Google Scholar]
- 21.Vaziri H, Benchimol S. Curr Biol. 1998;8:279–282. doi: 10.1016/s0960-9822(98)70109-5. [DOI] [PubMed] [Google Scholar]
- 22.Counter C M, Meyerson M, Eaton E N, Ellisen L W, Caddle S D, Haber D A, Weinberg R A. Oncogene. 1998;16:1217–1222. doi: 10.1038/sj.onc.1201882. [DOI] [PubMed] [Google Scholar]
- 23.de Lange T, DePinho R A. Science. 1999;283:947–949. doi: 10.1126/science.283.5404.947. [DOI] [PubMed] [Google Scholar]
- 24.Blasco M A, Lee H, Hande M, Samper E, Lansdorp P M, DePinho R A, Greider C W. Cell. 1997;91:25–34. doi: 10.1016/s0092-8674(01)80006-4. [DOI] [PubMed] [Google Scholar]
- 25.Lee H-W, Blasco M A, Gottlieb G J, Horner J W, Greider C W, DePinho R A. Nature (London) 1998;392:569–574. doi: 10.1038/33345. [DOI] [PubMed] [Google Scholar]
- 26.Kipling D, Cooke H J. Nature (London) 1990;347:400–402. doi: 10.1038/347400a0. [DOI] [PubMed] [Google Scholar]
- 27.Starling J A, Naule J, Hastie N D, Allshire R C. Nucleic Acids Res. 1990;18:6881–6888. doi: 10.1093/nar/18.23.6881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Shippen D E, McKnight T D. Trends Plant Sci. 1998;3:126–130. [Google Scholar]
- 29.Fitzgerald M S, McKnight T D, Shippen D E. Proc Natl Acad Sci USA. 1996;93:14422–14427. doi: 10.1073/pnas.93.25.14422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Heller K, Kilian A, Piatyszek M A, Kleinhofs A. Mol Gen Genet. 1996;252:342–345. doi: 10.1007/BF02173780. [DOI] [PubMed] [Google Scholar]
- 31.Fajkus J, Kovarik A, Kralovics R. FEBS Lett. 1996;391:307–309. doi: 10.1016/0014-5793(96)00757-0. [DOI] [PubMed] [Google Scholar]
- 32.Riha K, Fajkus J, Siroky J, Vyskot B. Plant Cell. 1998;10:1691–1698. doi: 10.1105/tpc.10.10.1691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Kilian A, Heller K, Kleinhofs A. Plant Mol Biol. 1998;37:621–628. doi: 10.1023/a:1005994629814. [DOI] [PubMed] [Google Scholar]
- 34.Kilian A, Stiff C, Kleinhofs A. Proc Natl Acad Sci USA. 1995;92:9555–9559. doi: 10.1073/pnas.92.21.9555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.McKnight T D, Fitzgerald M S, Shippen D E. Biochemistry (Moscow) 1997;62:1224–1231. [PubMed] [Google Scholar]
- 36.Richards E J, Chao S, Vongs A, Yang J. Nucleic Acids Res. 1992;20:4039–4046. doi: 10.1093/nar/20.15.4039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Murashige T, Skoog F. Physiol Plant. 1962;15:493–497. [Google Scholar]
- 38.Altschul S F, Madden T L, Schaffer A A, Zhang J H, Miller W, Lipman D J. Nucleic Acids Res. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Brendel V, Kleffe J. Nucleic Acids Res. 1998;26:4748–4757. doi: 10.1093/nar/26.20.4748. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Thompson J D, Higgins D G, Gibson T J. Nucleic Acids Res. 1994;22:4673–4680. doi: 10.1093/nar/22.22.4673. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.McKinney E C, Ali N, Traut A, Feldmann K A, Belostetsky D A, McDowell J M, Meagher R B. Plant J. 1995;8:613–622. doi: 10.1046/j.1365-313x.1995.8040613.x. [DOI] [PubMed] [Google Scholar]
- 42.Wang H, Qi M, Ctler A J. Nucleic Acids Res. 1993;21:4153. doi: 10.1093/nar/21.17.4153. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Li J, Chory J. In: Methods in Molecular Biology: Arabidopsis Protocols. Martnez-Zapater J M, Salinas J, editors. Totowa, NJ: Humana; 1998. pp. 55–60. [Google Scholar]
- 44.Harrington L, McPhail T, Mar V, Zhou W, Oulton R, Bass M B, Arruda I, Robinson M O. Science. 1997;275:973–977. doi: 10.1126/science.275.5302.973. [DOI] [PubMed] [Google Scholar]
- 45.Weinrich S L, Pruzan R, Ma L, Ouellette M, Tesmer V M, Holt S E, Bodnar A G, Lichtsteiner S, Kim N W, Trager J B, et al. Nat Genet. 1997;17:498–502. doi: 10.1038/ng1297-498. [DOI] [PubMed] [Google Scholar]
- 46.Wick M, Zubov D, Hagen G. Gene. 1999;232:97–106. doi: 10.1016/s0378-1119(99)00108-0. [DOI] [PubMed] [Google Scholar]
- 47.Azpiroz-Leehan R, Feldmann K A. Trends Genet. 1997;13:152–156. doi: 10.1016/s0168-9525(97)01094-9. [DOI] [PubMed] [Google Scholar]
- 48.Lundblad V, Szostak J W. Cell. 1989;57:633–643. doi: 10.1016/0092-8674(89)90132-3. [DOI] [PubMed] [Google Scholar]
- 49.McEachern M J, Blackburn E H. Nature (London) 1995;376:403–409. doi: 10.1038/376403a0. [DOI] [PubMed] [Google Scholar]
- 50.Harley C B, Futcher A B, Greider C W. Nature (London) 1990;345:458–460. doi: 10.1038/345458a0. [DOI] [PubMed] [Google Scholar]
- 51.Hahn W C, Counter C M, Lundberg A S, Beijersbergen R L, Brooks M W, Weinberg R A. Nature (London) 1999;400:464–468. doi: 10.1038/22780. [DOI] [PubMed] [Google Scholar]
- 52.Oguchi K, Liu H, Tamura K, Takahashi H. FEBS Lett. 1999;457:465–469. doi: 10.1016/s0014-5793(99)01083-2. [DOI] [PubMed] [Google Scholar]
- 53.Hande M P, Samper E, Lansdorp P, Blasco M A. J Cell Biol. 1999;144:589–601. doi: 10.1083/jcb.144.4.589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Lundblad V, Blackburn E H. Cell. 1993;73:347–360. doi: 10.1016/0092-8674(93)90234-h. [DOI] [PubMed] [Google Scholar]
- 55.Tovar J, Lichtenstein C. Plant Cell. 1992;1992:319–332. doi: 10.1105/tpc.4.3.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Lebel E G, Masson J E, Bogucki A, Paszkowski J. Proc Natl Acad Sci USA. 1993;90:422–426. doi: 10.1073/pnas.90.2.422. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Richards E R, Ausubel F M. Cell. 1988;53:127–136. doi: 10.1016/0092-8674(88)90494-1. [DOI] [PubMed] [Google Scholar]
- 58.Wellinger R J, Ethier K, Labrecque P, Zakian V A. Cell. 1996;85:423–433. doi: 10.1016/s0092-8674(00)81120-4. [DOI] [PubMed] [Google Scholar]
- 59.Meinke D W, Cherry J M, Dean C, Rounsley S D, Koorneef M. Science. 1998;282:678–682. doi: 10.1126/science.282.5389.662. [DOI] [PubMed] [Google Scholar]
- 60.Counter C M, Avilion A A, LeFeuvre C E, Stewart N G, Greider C W, Harley C B, Bacchetti S. EMBO J. 1992;11:1921–1929. doi: 10.1002/j.1460-2075.1992.tb05245.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Drost J B, Lee W R. Environ Mol Mutagen. 1995;25:48–64. doi: 10.1002/em.2850250609. [DOI] [PubMed] [Google Scholar]
- 62.Lansdorp P M. J Cell Biol. 1997;139:309–312. doi: 10.1083/jcb.139.2.309. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Laufs P, Grandjean O, Jonak C, Kieu K, Traas J. Plant Cell. 1998;10:1375–1389. doi: 10.1105/tpc.10.8.1375. [DOI] [PMC free article] [PubMed] [Google Scholar]