

Abstract
Coiled bodies are nuclear organelles that contain components of at least three RNA-processing pathways: pre-mRNA splicing, histone mRNA 3′- maturation, and pre-rRNA processing. Their function remains unknown. However, it has been speculated that coiled bodies may be sites of splicing factor assembly and/or recycling, play a role in histone mRNA 3′-processing, or act as nuclear transport or sorting structures. To study the dynamics of coiled bodies in living cells, we have stably expressed a U2B"–green fluorescent protein fusion in tobacco BY-2 cells and in Arabidopsis plants. Time-lapse confocal microscopy has shown that coiled bodies are mobile organelles in plant cells. We have observed movements of coiled bodies in the nucleolus, in the nucleoplasm, and from the periphery of the nucleus into the nucleolus, which suggests a transport function for coiled bodies. Furthermore, we have observed coalescence of coiled bodies, which suggests a mechanism for the decrease in coiled body number during the cell cycle. Deletion analysis of the U2B" gene construct has shown that the first RNP-80 motif is sufficient for localization to the coiled body.
INTRODUCTION
The coiled body was first described by Ramon y Cajal (1903), who called it the “nucleolar accessory body” because of its association with the nucleolus. This nuclear organelle was later reidentified by electron microscopy and renamed the “coiled body” because of its appearance as loosely packed coiled fibrils (Monneron and Bernhard, 1969). Subsequent studies detected coiled bodies in animal and plant nuclei, showing that it is a conserved structure (Moreno Diaz de la Espina et al., 1980; Seite et al., 1982; Schultz, 1990).
Coiled bodies have been shown to contain splicing small nuclear ribonucleoproteins (snRNPs) and small nuclear RNAs (snRNAs), a subset of nucleolar components — including fibrillarin, Nopp140, NAP57, and U3 small nucleolar ribonucleoprotein (U3 snoRNP) — and the protein p80 coilin, which has been widely used as a marker for coiled bodies (Lamond and Earnshaw, 1998; Matera, 1998). The function of coiled bodies is still under debate, but several hypotheses have been proposed that are not necessarily mutually exclusive. Because coiled bodies do not contain DNA, nascent pre-mRNA, heterogeneous nuclear RNPs (hnRNPs), or the SC-35 splicing factor, which is required for splicing in vitro, it has been argued that they are not directly involved in transcription and pre-mRNA splicing (Lamond and Carmo-Fonseca, 1993). However, it has been speculated that coiled bodies may be sites of splicing factor assembly or recycling, may play a role in histone mRNA 3′ processing (Gall et al., 1995; Lamond and Earnshaw, 1998), or may be involved in all these activities. As coiled bodies are frequently observed at the nucleolar periphery and also in the nucleoplasm and within nucleoli (Malatesta et al. 1994; Ochs et al., 1994), they may also act as nuclear transport or sorting structures. It has been shown recently that coiled bodies are also involved in processing or transport of small nucleolar RNA (snoRNA) precursors in maize (Shaw et al., 1998).
Coiled bodies have been shown to be dynamic structures, largely based upon immunofluorescence and in situ studies. When transcription is inhibited, splicing snRNPs no longer concentrate in coiled bodies but, instead, aggregate in large clusters thought to be storage sites of splicing factors (Carmo-Fonseca et al., 1992). Changes have also been observed in the size and number of coiled bodies during the cell cycle (Andrade et al., 1993; Carmo-Fonseca et al., 1993; Chan et al., 1994; Ferreira et al., 1994; Beven et al., 1995), with smaller and more coiled bodies during G1 phase, and larger and fewer coiled bodies in S and G2 phases of the cell cycle. Upon entry into mitosis, most coiled bodies disappear, reappearing in G1 phase after reformation of nucleoli (Andrade et al., 1993; Ferreira et al., 1994). Changes in the number of coiled bodies as a function of the differentiation stage (Antoniou et al., 1993; Santama et al., 1996; Boudonck et al., 1998), and after virus infection (Fortes et al., 1995; Rebelo et al., 1996), have been described. Exposure of mammalian cells to low levels of the specific Ser/Thr protein phosphatase inhibitor, okadaic acid, results in the accumulation of p80 coilin and splicing snRNPs within nucleoli, which indicates that protein dephosphorylation is required for normal formation of nucleoplasmic coiled bodies (Lyon et al., 1997). Sleeman et al. (1998) expressed fusions of the spliceosomal proteins, SmE and U1A, and p80 coilin with green fluorescent protein (GFP) in human cell lines. This work confirmed earlier immunofluorescence and in situ studies that p80 coilin localizes to coiled bodies in a transcription-dependent manner and that a reversible protein phosphorylation mechanism is involved in regulating the interaction of snRNPs and coiled bodies with the nucleolus.
To investigate the dynamics of coiled bodies in living cells, we have generated a protein fusion between GFP and the U2B" spliceosomal protein that is, among others, concentrated in coiled bodies. Although U2B" is a component of the U2 snRNP complex, its precise role in pre-mRNA splicing is still unknown. It has been suggested that U2B" may not be required for the splicing reaction itself but may have a role in U2 snRNP biogenesis (Mattaj and De Robertis, 1985; Pan and Prives, 1989). The full-length U2B" cDNA has been cloned from potato (Simpson et al., 1991) and has been characterized at the molecular level (Simpson et al., 1991, 1995), largely by analogy with the human U2B" cDNA (Scherly et al., 1990; Bentley and Keene, 1991; Kambach and Mattaj, 1994). The potato U2B" protein contains two RNP-80 motifs separated by a central putative nuclear localization signal (NLS). The RNP motifs consist of 80–90 amino acids containing two short, highly conserved sequences (RNP1 and RNP2) and are found in numerous proteins involved in RNA processing events (Birney et al., 1993; Mattaj, 1993). It has been shown that only the N-terminal RNP motif of U2B" is required for specific binding to U2 snRNA (Kambach and Mattaj, 1994; Simpson et al., 1995) and that the interaction between U2B" and U2 snRNA requires the presence of a second protein, U2A′ (Scherly et al., 1990; Simpson et al., 1995).
In the present work we show directly for the first time, by time-lapse confocal fluorescence microscopy, that coiled bodies can move within the nucleus and can coalesce. These findings suggest a transport function for coiled bodies and a mechanism for the observed decrease in coiled body number during the cell cycle. We have shown that overexpression of the U2B"–GFP fusion affects neither the cell cycle and the number of coiled bodies in tobacco BY-2 cells nor development of Arabidopsis plants. Deletion analysis of the U2B" gene fusion has identified the N-terminal RNP-80 motif as sufficient for localization to the coiled body and the nucleus.
MATERIALS AND METHODS
Construction of the U2B"–GFP Fusion Protein
The full-length 693-base pair (bp) sequence of the potato U2B" cDNA (Simpson et al., 1991) was PCR amplified with specific primers introducing NcoI sites (underlined) at both termini for further cloning steps. At the same time the second methionine codon, the stop codon and an internal NcoI site near the 3′-end of the U2B" gene, were mutated (base changes shown in lowercase).
primer 1: 5′-AGCGCCCC ATG gTG CTT ACA GGA GAC A-3′
primer 2: 5′-GGTACGCCATggg TTT CTT GGC ATA GGT GAT TGC CAT tGG ATT TTG GG-3′
The amplified U2B" gene was fused in frame to the 5′-end of sGFPS65T (Chiu et al., 1996) by use of the NcoI site. The U2B"::GFP cassette was cut out and inserted into the pRTL2 vector (Carrington et al., 1991), creating the pRTL2::U2B"::GFP vector. The new U2B"::GFP construct was checked by sequence analysis. For stable transformation experiments, the expression cassette (2× En CaMV 35S promotor, TEV 5′-nontranslated sequence, U2B"::GFP coding sequence, and CaMV polyadenylation site) from the vector pRTL2::U2B"::GFP was excised, using HindIII, and inserted into the HindIII site of the vector pGreen0229, which carries the bar gene (http://intranet.jic.bbsrc.ac.uk/INFOSERV/DEPART/appgen/pgreen/a_cst_ fr.htm by Roger Hellens), creating the pGreen0229::U2B"::GFP vector.
Construction of the U2B"del::GFP Deletion Cassettes
We constructed several U2B"::GFP fusions with different deletions in the U2B" gene (Figure 10). For a first deletion construct we PCR amplified the first 276 bp of the U2B" gene, corresponding to the N-terminal RNP-80 motif, introducing NcoI sites (underlined) at both termini for further cloning steps. Primers 1 and 3 were used for PCR amplification.
Figure 10.

Stable U2B"::GFP expression in Arabidopsis roots. Projections of series of confocal sections through the root meristem are shown. (A) The full-length U2B"::GFP fusion shows expression in the nucleoplasm and in the coiled bodies (bright spots) in all root nuclei in the epidermis. The nucleolus is largely devoid of labeling (dark centers). Root cap nuclei are labeled very brightly (small nuclei). (B and C) Deletion constructs U2B"del1::GFP and U2B"del2::GFP label the nucleoplasm and the coiled bodies. In addition, cytoplasmic strands emerging from the nucleus and plastid-like structures are clearly labeled in the root meristem of U2B"del1::GFP transformants. (D) Fusion of the U2B" putative NLS to GFP results in labeling of the cytoplasm and somewhat brighter labeling of the nucleoplasm. There is no labeling of coiled bodies. Bar, 20 μm.
primer 3: 5′-GGTACGCCATGGG TGA CTT TGA TTT AGC-3′
For a second deletion construct we PCR amplified the first 453 bp of the U2B" gene, corresponding to the N-terminal RNP-80 motif plus a putative NLS, introducing NcoI sites at both termini for further cloning steps. Primers 1 and 4 were used for PCR amplification.
primer 4: 5′-GGTACGCCatgGG ATC TTG AGC ACT TGG-3′
For a third deletion construct, we PCR amplified nucleotide (nt) 286–453 of the U2B" gene, corresponding to a putative NLS, introducing NcoI sites at both termini for further cloning steps and introducing a new start codon at the 5′-end (lowercase). Primers 4 and 5 were used for PCR amplification.
primer 5: 5′-AGCGCCCC atg GTT GCT AAG GCA G-3′
The amplified U2B" deletion fragments were fused in frame to the 5′-terminus of sGFPS65T using the NcoI site in the pRTL2::GFP vector, creating vectors pRTL2::U2B"del1::GFP, pRTL2::U2B"del2::GFP, and pRTL2::U2B"del3::GFP, respectively. The three gene fusions were checked by sequence analysis. For stable transformation experiments, the expression cassettes (2× En CaMV 35S promotor, TEV 5′-nontranslated sequence, U2B"del::GFP coding sequence, and CaMV polyadenylation site) from the vectors pRTL2::U2B"del::GFP were excised using HindIII and inserted into the HindIII site of the vector pGreen0229, which carries the bar gene, creating the pGreen0229::U2B"del::GFP vectors.
Stable Transformation of Tobacco BY-2 Cells and Arabidopsis thaliana
The pGreen0229::U2B"::GFP vectors were transformed into Agrobacterium strain C58C1 pGV3101 (pMP90) (Koncz and Schell, 1986), carrying the helper plasmid pJIC Sa_RepA. BY-2 tobacco cells were transformed by Agrobacterium essentially as described by Ito et al. (1998) and An (1985, 1987). After transformation, BY-2 cells were plated onto BY-2 medium (per liter: 4.3 g Murashige and Skoog salts; 30 g sucrose; 0.1 g inositol; 1 mg thiamine; 0.2 mg 2,4-dichlorophenoxyacetic acid (2,4-D); 0.2 g KH2PO4) + 0.4% Gelrite, containing carbenicillin (500 mg/l) to kill Agrobacteria and phosphinotricin (10 mg/l) to select transformed BY-2 cells. After selection for 3–4 wk, transformed BY-2 cells were maintained as suspension cultures in BY-2 medium and grown at 25°C in the dark.
Transgenic Arabidopsis plants, ecotype Columbia, were generated by Agrobacterium transformation using vacuum infiltration as described by Bechtold and Pelletier (1998). Transgenic T1 plants, carrying the bar gene, were selected on soil by spraying with the herbicide Challenge (2.5 ml Challenge/l water) (Duchefa, The Netherlands).
Time-Lapse Fluorescence Microscopy
For time-lapse confocal microscopy, cells from GFP-expressing BY-2 cultures, 5 d after subculture, were spun down and transferred onto BY-2 medium + 1% Difco Bacto agar in Petri dishes. A coverslip was placed on top. For confocal microscopy of GFP-expressing T2 Arabidopsis seedlings, seeds were germinated and grown as described by Boudonck et al. (1998). Three-day-old seedlings were mounted on slides in liquid AT growing medium (per liter: 4.4 g Murashige and Skoog salts, 30 g sucrose, 0.05 mg kinetin, 0.5 mg α-naphthaleneacetic acid (NAA), pH 5.8). A coverslip was placed on top and sealed with nail varnish. Optical sections were collected on living BY-2 cells or Arabidopsis seedlings using a confocal microscope (MRC 1024, Bio-Rad, Richmond, CA; or TCS SP, Leica, Deerfield, IL) with 488-nm excitation line. Images were transferred to a Macintosh computer and assembled into composite images using Adobe Photoshop and NIH Image, a program for the Macintosh available via http://rsb.info.nih.gov/nih-image. Images were printed on a Pictrography 3000 printer.
For heat shock treatment of BY-2 cells, cells were imaged before heat shock. Petri dishes were then transferred to 42°C for 1 h. The same cells were then relocated and imaged over time. For inhibition of transcription in BY-2 cells by actinomycin D, 50 μl of 2 mM actinomycin D (Sigma Chemical, St. Louis, MO) was applied to the cells on the Petri dish. For treatment with okadaic acid, 50 μl of 100 nM okadaic acid were applied to the cells on the Petri dish.
Immunofluorescence on BY-2 Cells
Five-day old BY-2 cells were fixed for 1 h in 4% (wt/vol) formaldehyde in PEM buffer (50 mM piperazine-N,N′-bis(2-ethanesulfonic acid), 5 mM EGTA, 5 mM MgSO4; pH 6.9). The fixed cells were washed in PEM for 30 min followed by permeabilization with 2% (wt/vol) cellulase in TBS (25 mM Tris-HCl [pH 7.4], 140 mM NaCl, 3 mM KCl) for 1 h. Cells were washed four times with TBS and finally dried down on to glutaraldehyde-activated APTES (γ-aminopropyltriethoxysilane)-coated multiwell slides. A blocking step for 1 h, with PEM/0.2% NP40/3% BSA followed. The slides were incubated for 1.5 h at 37°C with a 1:10 dilution of primary antibody 4G3 (Euro-diagnostica B.V., Apeldoorn, Netherlands) or a 1:500 dilution of human autoantibody against p80 coilin (a gift from Professor Angus Lamond, University of Dundee, Scotland) in PEM/3% BSA. The primary antibody was detected by cy3-conjugated secondary antibody (Jackson ImmunoResearch Laboratories, West Grove, PA). The slides were mounted in Vectashield (Vector laboratories, Burlingame, CA). GFP kept its fluorescence through this treatment.
RESULTS
U2B"-GFP Is a Good Marker for the Coiled Body
No homologue of the p80 coilin gene has yet been identified in plants. However, the U2B" protein is a well characterized plant spliceosomal protein that has been shown to be a good marker for plant coiled bodies (Beven et al., 1995; Boudonck et al., 1998). To study the dynamics of coiled bodies in detail, therefore, we generated a translational fusion between GFP and the full-length plant U2B" protein and made stable transgenic lines of both BY-2 tobacco suspension culture cells and Arabidopsis plants expressing this fusion. BY-2 suspension culture cells have large nuclei, contain many prominent coiled bodies, and are particularly well suited for microscopy. To confirm that the U2B"–GFP fusion protein is expressed in coiled bodies in the BY-2 cells, we combined fluorescence imaging of the GFP with immunofluorescence labeling using an autoantibody against p80 coilin. Figure 1A shows that the bright GFP spots do indeed correspond to coiled bodies and demonstrates that all the coiled bodies contain the U2B"–GFP fusion protein. Figure 1B confirms our previous observations that the anti-U2B" antibody, 4G3 (Habets et al., 1989), is a good marker for coiled bodies in plants. In these cells, coiled bodies are most frequently found at the nucleolar periphery, but can also be found throughout the nucleoplasm and inside the nucleolus as well as in the central nucleolar cavity.
Figure 1.

(A) Single confocal optical section of BY-2 cells expressing U2B"-GFP, double labeled with GFP (left panel) and autoantibody against p80 coilin (right panel). Three nuclei are shown, and the bright GFP spots colocalize with bright foci of anti-coilin labeling. There is some labeling of the cytoplasm by anti-p80 coilin. (B) Single confocal optical section of BY-2 cells expressing U2B"-GFP, double labeled with GFP (left panel) and 4G3 antibody (right panel). Three nuclei are shown. Most coiled bodies are in the nucleoplasm, but occasionally are seen in the nucleolus (arrows). All coiled bodies that contain U2B" also express the U2B"–GFP fusion. Bars, 5 μm.
To determine whether overexpression of the U2B"–GFP protein fusion affected the numbers of coiled bodies in BY-2 cells, we labeled cells expressing U2B"-GFP, and untransformed BY-2 cells, with the 4G3 antibody. Table 1 shows that there is no difference in the number of coiled bodies between BY-2 cells transformed or not transformed with the U2B"::GFP fusion. To investigate whether the expression of the U2B"–GFP fusion affected the cell cycle duration, we set up fresh subcultures and measured the optical density (OD600) for 8 successive days for BY-2 cells transformed or not transformed with the U2B"::GFP fusion. In five such time courses we found no significant difference between the growth curves for cells transformed with the U2B"::GFP construct and wild-type BY-2 cells (our unpublished results).
Table 1.
Coiled body numbers in tobacco BY-2 cells
| Mean CB number ± SEM (no. of cells counted) | Min. CBa | Max. CBa | |
|---|---|---|---|
| Untransformed BY-2 | 8.88 ± 2.22 (60) | 4 | 15 |
| U2B"-GFP BY-2 | 8.97 ± 2.19 (60) | 4 | 15 |
Minimum and maximum coiled body number observed in these cells.
We also found no effect of the U2B::GFP transgene on development or fertility of Arabidopsis compared with wild type. The 1.5 times difference in coiled body number between root hair and hairless files in the Arabidopsis root epidermis, as described by Boudonck et al. (1998), was maintained in lines transformed with U2B::GFP (e.g., see Figure 10).
Coiled Bodies Move and Coalesce in Plant Cells
We used time-lapse confocal microscopy of U2B"-GFP–expressing BY-2 cells to investigate the mobility of coiled bodies. We were able to collect three-dimensional (3-D) data sets consisting of ∼15 focal sections at intervals of 30–60 min for periods up to 15 h, without unacceptable fading of the fluorescence. We examined ∼60 cells in this way, and 70% of these cells showed movements of some or all of the coiled bodies over an average time range of 6 h. Figure 2 shows 5 of the 18 confocal sections through a single cell at one time point, together with projections of the entire confocal stack. In order to present four-dimensional data in most subsequent figures, we show only a projection of the 3-D data at each time point, but the interpretation of the data was made from the original 3-D image stacks.
Figure 2.

Confocal fluorescence microscopy of U2B"-GFP in a living BY-2 cell. (A) Single confocal optical sections of a BY-2 cell nucleus are shown, with 2.6 μm between consecutive sections, followed by a projection of the 3-D data stack. Coiled bodies can be found in the nucleoplasm and nucleolus, but most frequently just inside the nucleolar periphery. (B) Stereo projection of the confocal series shown in panel A. Bar, 5 μm.
The movements of coiled bodies that we observed in BY-2 cells can be categorized into a number of distinct classes. The most frequent were movements of coiled bodies within the nucleolus and the nucleoplasm, which occurred in 70% (42 cells) of the 60 cells examined. Coiled bodies were also seen to move along the boundary between nucleolus and nucleoplasm (Figure 3), in more than 20% of the 42 cells that showed coiled body dynamics. The most striking observations were unidirectional movements of coiled bodies from the nuclear periphery to the nucleolar periphery (Figure 4) in more than 25% of the 42 cells that showed coiled body dynamics. The velocity of these movements ranged from fractions of 1 μm/h to ∼10 μm/h. We frequently observed very small coiled bodies within the nucleolus, which were also mobile, but were difficult to image clearly because of their small size and rapid photobleaching of GFP. In more than 20% of the 42 cells with coiled body dynamics, two or more coiled bodies moved toward one another and then fused together (Figure 4). Although we saw movements of coiled bodies toward one another in all regions of the nucleus, we observed fusions only at the nucleolar periphery.
Figure 3.

Time-lapse confocal microscopy of U2B"-GFP in living BY-2 cells. Projections of three or four confocal optical sections are shown at each time point. Coiled body marked with an arrow demonstrates that coiled bodies can move around in the nucleoplasm and nucleolus, and often move along the periphery of the nucleolus. Other, unmarked coiled bodies show smaller movements. Bar, 5 μm.
Figure 4.

Time-lapse confocal microscopy of U2B"-GFP in living BY-2 cells. Projections of series of confocal optical sections are shown at each time point. Coiled body marked with an arrow moves from the periphery of the nucleoplasm to the periphery of the nucleolus, after which it moves along the periphery of the nucleolus and finally fuses with another coiled body. Other coiled bodies also show movements. Bar, 5 μm.
Figure 5 shows a pair of cells at late telophase/early G1. As G1 progressed, the size of the nuclei increased and the number of coiled bodies concomitantly decreased. This decrease in coiled body number was accompanied by an increase in the size of the coiled bodies. The number of coiled bodies and the complexity of their distribution made it impossible to follow their dynamics in detail.
Figure 5.

Time-lapse confocal microscopy of U2B"-GFP in living BY-2 cells. Projections of series of confocal optical sections are shown at each time point. Late telophase/early G1 cells have many small coiled bodies. As the cells progress through the cell cycle, the nuclei enlarge and the number of coiled bodies decreases. Concomitantly, the size of the coiled bodies increases. Bar, 10 μm.
Figure 6 shows that Arabidopsis root epidermal cells display coiled body dynamics similar to those of BY-2 cells. These images show movements of coiled bodies in the nucleoplasm and at the periphery of the nucleolus. Coiled bodies frequently coalesced, as we observed earlier for BY-2 cells (Figure 6, arrows). This image sequence also shows two cells undergoing mitosis (starred cells). In this sequence, mitosis, accompanied by redistribution of the U2B"-GFP throughout the cytoplasm, was followed by reformation of the coiled bodies in the G1 daughter cells in the space of 2 h.
Figure 6.

Time-lapse confocal microscopy of U2B"-GFP in stable Arabidopsis transformants. Projections of series of confocal optical sections through the root epidermis are shown at each time point. Many coiled bodies are clearly mobile. Several coiled bodies coalesce (arrows). Two cells underwent division during this sequence, and the newly divided cells decreased their coiled body numbers quickly (asterisk). Bar, 10 μm.
Dynamics of Coiled Bodies Are Affected by Regulators of Transcription and Phosphorylation in Plant Cells
Heat shock is known to inhibit transcription of most RNA pol II genes and disrupts pre-mRNA splicing in both plants and animals (Bond, 1988; Christensen et al., 1992). Transfer of U2B"-GFP BY-2 cells to 42°C for 1 h resulted in the disappearance of all coiled bodies (Figure 7). After heat shock the coiled bodies reformed throughout the nucleus within hours. The heat shock could be repeated on the same cells for a second time, with subsequent second reformation of coiled bodies. There was an indication that some of the coiled bodies reformed near previous positions (Figure 7, arrows).
Figure 7.

Heat shock on U2B"-GFP–expressing BY-2 cells. Projections of series of confocal optical sections are shown at each time point. (A) Cells before heat shock. (B) After 1 h heat shock, at 42°C, all coiled bodies have disappeared. (C) 19.5 h after heat shock, coiled bodies have reformed. (D) A second heat shock, at 42°C, for 1 h, results in the disappearance of all coiled bodies. (E) 2.5 h after heat shock, coiled bodies are already present. Several coiled bodies reappear near places where they were before heat shock (arrows). Bar, 5 μm.
Treatment of U2B"-GFP BY-2 cells with 2 mM actinomycin D, a potent inhibitor of transcription, resulted in the redistribution of some coiled bodies as elongated structures around the nucleolus, while other coiled bodies remained in the nucleoplasm (Figure 8). The elongated structures were dynamic, moving around the nucleolus and changing in size.
Figure 8.

Actinomycin D treatment of U2B"-GFP–expressing BY-2 cells. Projections of series of confocal optical sections are shown at each time point. After addition of 2 mM actinomycin D, at time point 0 h, U2B"-GFP forms elongated structures around the nucleolus (arrow). These elongated structures change their relative positions around the nucleolus and can increase or decrease in size. Bar, 5 μm.
We tested the effect of okadaic acid, a specific Ser/Thr protein phosphatase inhibitor that has been shown to result in the accumulation of p80 coilin and splicing snRNPs within nucleoli in mammalian cells (Lyon et al., 1997; Sleeman et al., 1998). Figure 9 shows that 19 h after addition of 100 nM okadaic acid to U2B"-GFP BY-2 cells, all the coiled bodies in most cells were located within the nucleolus. This indicates that one possible mechanism for nucleolar localization of coiled bodies is through a phosphorylation/dephosphorylation mechanism.
Figure 9.
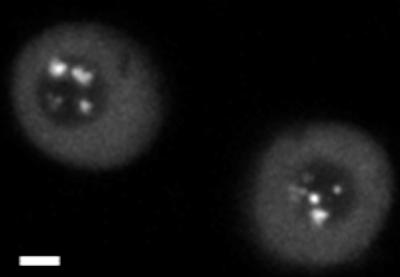
Okadaic acid treatment of U2B"-GFP–expressing BY-2 cells. A single confocal section is shown. After 19 h treatment with 100 nM okadaic acid, coiled bodies are mainly found in the nucleolus in BY-2 cells. Bar, 5 μm.
We also tested the effect of melittin and mastoparan, which have been shown to activate the plant phosphoinositide-signaling system (Drobak and Watkins, 1994). Addition of these drugs did not affect the dynamics or location of coiled bodies, which suggests that the phosphoinositide-signaling pathway is not involved in the movements of coiled bodies (our unpublished results).
N-terminal U2B" RNP-80 Motif Is Sufficient for Localization to the Coiled Body
Stable expression of the full-length U2B"::GFP fusion in Arabidopsis thaliana resulted in nucleoplasmic localization with clear GFP expression in coiled bodies (Figure 10A). This localization is the same as described previously using antibodies against U2B" (Boudonck et al., 1998). To investigate which part of the U2B" protein is required for localization to the coiled body, we made three additional stably transformed Arabidopsis lines using three deletion constructs: U2B"del1::GFP, U2B"del2::GFP and U2B"del3::GFP. U2B"del1-GFP, which contains the first U2B" RNP-80 motif, was localized to the coiled bodies (Figure 10B) and also to fluorescent strands emerging from the nucleus and to plastid-like structures in the root cap and in the hypocotyl (our unpublished results). U2B"del2-GFP, which contains the RNP-80 motif plus the putative NLS sequence, showed the same localization pattern as the full-length U2B"-GFP protein fusion (Figure 10C). U2B"del3-GFP, which only contains the putative NLS from U2B", was distributed throughout the cytoplasm and within the nucleoplasm, with no labeling of coiled bodies and weaker labeling of the nucleolus (Figure 10D). All the transformant seedlings examined showed GFP fusion protein expression in roots, hypocotyl, and cotyledons. In summary, these data demonstrate that the U2B"–GFP fusion protein is localized to coiled bodies throughout the plant and that the first part of the U2B" protein is sufficient for localization to the coiled body.
DISCUSSION
We have shown, using lines stably expressing the U2B"::GFP fusion construct, that coiled bodies are mobile organelles in living plant cells. We have observed both dramatic movements of coiled bodies from the nuclear periphery to the nucleolus and also smaller movements within both the nucleoplasm and nucleolus. Furthermore, we have seen frequent events in which coiled bodies fuse together. We have demonstrated that overexpression of the U2B"–GFP fusion does not affect the cell cycle duration or viability of BY-2 cells or the growth and development of Arabidopsis plants. Therefore, the observed movements of coiled bodies in cells transformed with U2B"::GFP are likely to faithfully reflect the natural movements in untransformed cells.
Although other researchers have observed coiled bodies in living cells, our results are the first to show clear movements over time. Almeida et al. (1998) detected no movements of coiled bodies in HeLa cells after injection of fluorescent-labeled antibodies to coilin. This inconsistency with our results could be due to the immobilization of coiled bodies by antibody binding, or inhibition of movement by phototoxicity. Sleeman et al. (1998) also visualized coiled bodies in living human cells by fusing a number of snRNP proteins and p80 coilin to GFP, but only observed small movements of coiled bodies (Sleeman, personal communication). It is entirely possible that we have been able to observe more and larger coiled body movements because plant cells are more resistant to photodamage. However, the possibility remains that coiled bodies are less mobile in animal than plant cells.
We have shown that coiled bodies can move unidirectionally from the nucleoplasm to the nucleolar periphery. This shows that nucleoplasmic and nucleolar coiled bodies are one and the same. Ochs et al. (1994) reported that nucleolar coiled bodies differed in composition from those found in the nucleoplasm. This suggests that some components of a coiled body can be modified or replaced and may be either the result or the cause of differential localization of coiled bodies. So far, we have observed unidirectional movement of coiled bodies only from the nucleoplasm into the nucleolus. Such a directional movement might indicate a transport function for coiled bodies (Lamond and Earnshaw, 1998). It could provide a mechanism for the import of factors involved in nucleolar transcript processing, such as fibrillarin, into the nucleolus, or it might provide a mechanism for shuttling of proteins such as Nopp140 and NAP57 between the nucleoplasm and nucleolus. Alternatively, the coiled body might be the initial site for assembly of processing complexes, which are then sorted to other locations in the nucleus or nucleolus where the actual RNP processing takes place, as suggested earlier by Gall et al. (1995).
While most coiled bodies in a given nucleus undergo constant small movements, a coiled body will occasionally undergo a sudden dramatic movement over longer distances. This suggests that these large movements of coiled bodies are triggered by a biochemical modification. We currently have no detailed quantitative data on the maximum speed and the time scale of such sudden movements, as we cannot predict them, and imaging at time points too close together causes bleaching of the GFP. The accumulation of coiled bodies in the nucleolus upon addition of okadaic acid shows that phosphorylation is likely to be an important factor for localization (Lyon et al., 1997; Sleeman et al., 1998) and that the mobile coiled bodies may differ in phosphorylation state from the more static bodies. Addition of actinomycin D, an inhibitor of transcription, results in the accumulation of U2B"–GFP protein into elongated structures around the nucleolus, which are mobile and change in size. This suggests a link between transcriptional activity and snRNP location, which may either be a direct effect or the result of changes in chromatin or other nuclear organization.
Previous studies have shown that the nucleus is organized into chromosome territorial domains and interchromosomal channels (Cremer et al., 1988; Lichter et al., 1988; Abranches et al., 1998). It will be instructive to investigate whether coiled bodies move through interchromosomal channels, or along an underlying nuclear matrix, and whether specific motor proteins are required. Alternatively, as some coiled bodies have been found associated with specific chromosomal loci, including gene clusters that encode U snRNAs and histone mRNAs (Frey and Matera, 1995; Smith et al., 1995), and as there is direct evidence that chromosome movements take place in several cell types (Li et al., 1998; review by Zink and Cremer, 1998), it is possible that the movements of coiled bodies are related to underlying movements of chromosome loci to which they are attached. On the other hand, as Frey et al. (1999) showed that the frequency of colocalization of coiled bodies with artificial U1 and U2 snRNA genes depends on the transcriptional activity of the gene array, the movement of a coiled body might be the result of the termination of transcription and the subsequent release of the coiled body from a particular chromosome locus.
We have also shown directly that coiled bodies can coalesce. This fusion mechanism explains the observed decrease in coiled body number and the increase in size through the cell cycle that has been reported for several organisms (Andrade et al., 1993; Chan et al., 1994; Boudonck et al., 1998). It is noteworthy that all coiled body fusions that we observed took place at the nucleolar periphery. The elongated structures observed at the nucleolar periphery on treatment with actinomycin D might be the result of the fusion of several coiled bodies into elongated structures. Other nuclear bodies may also coalesce. Boddy et al. (1997) demonstrated that nuclear matrix-associated promyelocytic leukemia bodies are often located together and linked in pairs, as though in the process of fusing. However, pairing of the promyelocytic leukemia nuclear bodies could also be explained as budding or replication, since no studies have been carried out in living cells to date.
The biogenesis of coiled bodies remains an open question. It has been suggested that coiled bodies are assembled at the nucleolar periphery (Bohmann et al., 1995), and coiled bodies are often found adjacent to nucleoli or physically attached to nucleoli, as though in the process of emergence or fusion (Lafarga et al., 1983; Ferreira and Carmo-Fonseca, 1995; Matera, 1998). In most BY-2 cells, we observed very small coiled bodies inside the nucleolus, which were either too small or photobleached too quickly to follow during time-lapse experiments. It is possible that these small coiled bodies are newly formed in the nucleolus; however we saw no sign of such nucleolar coiled bodies emerging into the nucleoplasm. On the other hand, the reappearance of U2B"–GFP in coiled bodies in similar locations on recovery from heat shock shows that coiled bodies can reform in the nucleoplasm. However, it is possible that heat shock leaves a remnant of the coiled body structure and U2B"–GFP and other coiled body components simply reaccumulate in preexisting coiled body structures when transcription restarts after the heat shock. Supporting evidence for a biogenesis model in which coiled bodies form throughout the nucleus comes from the study of coiled bodies in late telophase/early G1 cells. Coiled bodies are first visible as several tens of coiled bodies or their precursors located all over the nucleus (Beven et al., 1995; present study). We have seen no evidence of any concentration of coiled bodies around the nucleolus at this stage, as might be expected if it was the site of coiled body biogenesis.
Mutation analysis of the human U2B" gene has previously shown which part of the sequence localizes U2B" to the nucleus (Scherly et al., 1990; Bentley and Keene, 1991; Kambach and Mattaj, 1994), but not which part of the sequence is responsible for localization to the coiled body. We have shown that fusion of the first N-terminal RNP-80 motif of the potato U2B" protein to GFP is sufficient for localization of the fusion protein to the coiled body. As Simpson et al. (1995) showed that the plant N-terminal RNP-80 motif is responsible for binding to the U2 snRNA and that this binding is mediated by the U2A′ protein, our deletion analysis results demonstrate that localization of the U2B" protein to the coiled body is mediated by binding of the U2B" protein to the U2A′ protein and U2 snRNA, and that the central NLS domain of the U2B" protein alone is not sufficient for localization to the coiled body.
As we have produced stable transgenic Arabidopsis lines that express the U2B"::GFP fusion in the coiled bodies of all cells, we are now able to screen for coiled body mutants with defects in their size, number, or organization. Such studies should shed new light on the function of these intriguing nuclear organelles.
ACKNOWLEDGMENTS
This work was supported by the European Commission (Training and Mobility of Researchers grant ERBFMBICT961250) and by the Biotechnology and Biological Sciences Research Council of the United Kingdom.
REFERENCES
- Abranches R, Beven AF, Arágon-Alcaide L, Shaw PJ. Transcription sites are not correlated with chromosome territories in wheat nuclei. J Cell Biol. 1998;143:5–12. doi: 10.1083/jcb.143.1.5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Almeida F, Saffrich R, Ansorge W, Carmo-Fonseca M. Microinjection of anticoilin antibodies affects the structure of coiled bodies. J Cell Biol. 1998;142:899–912. doi: 10.1083/jcb.142.4.899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- An G. High efficiency transformation of cultured tobacco cells. Plant Physiol. 1985;79:568–570. doi: 10.1104/pp.79.2.568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- An G. Binary Ti vectors for plant transformation and promotor analysis. Methods Enzymol. 1987;153:292–305. [Google Scholar]
- Andrade LEC, Tan EM, Chan EKL. Immunocytochemical analysis of the coiled body in the cell cycle and during cell proliferation. Proc Natl Acad Sci USA. 1993;90:1947–1951. doi: 10.1073/pnas.90.5.1947. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Antoniou M, Carmo-Fonseca M, Ferreira J, Lamond AI. Nuclear organization of splicing snRNPs during differentiation of murine erythroleukemia cells in vitro. J Cell Biol. 1993;123:1055–1068. doi: 10.1083/jcb.123.5.1055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bechtold N, Pelletier G. In planta Agrobacterium-mediated transformation of adult Arabidopsis thaliana plants by vacuum infiltration. Methods Mol Biol. 1998;82:259–266. doi: 10.1385/0-89603-391-0:259. [DOI] [PubMed] [Google Scholar]
- Bentley RC, Keene JD. Recognition of U1 and U2 small nuclear RNAs can be altered by a 5-amino-acid segment in the U2 small nuclear ribonucleoprotein particle (snRNP) B" protein and through interactions with U2 snRNP-A′ protein. Mol Cell Biol. 1991;11:1829–1839. doi: 10.1128/mcb.11.4.1829. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beven AF, Simpson GG, Brown JWS, Shaw PJ. The organization of spliceosomal components in the nuclei of higher plants. J Cell Sci. 1995;108:509–518. doi: 10.1242/jcs.108.2.509. [DOI] [PubMed] [Google Scholar]
- Birney E, Kumar S, Krainer AR. Analysis of the RNA-recognition motif and RS and RGG domain: conservation in metazoan premRNA splicing factors. Nucleic Acids Res. 1993;21:5803–5816. doi: 10.1093/nar/21.25.5803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boddy MN, Duprez E, Borden KLB, Freemont P. Surface residue mutations of the PML RING finger domain alter the formation of nuclear matrix-associated PML bodies. J Cell Sci. 1997;110:2197–2205. doi: 10.1242/jcs.110.18.2197. [DOI] [PubMed] [Google Scholar]
- Bohmann K, Ferreira J, Santama N, Weis K, Lamond AI. Molecular analysis of the coiled body. J Cell Sci. 1995;19,suppl:107–113. doi: 10.1242/jcs.1995.supplement_19.16. [DOI] [PubMed] [Google Scholar]
- Bond U. Heat shock but not other stress inducers leads to the disruption of a subset of snRNPs and inhibition of in vitro splicing in HeLa cells. EMBO J. 1988;7:3509–3518. doi: 10.1002/j.1460-2075.1988.tb03227.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boudonck K, Dolan L, Shaw PJ. Coiled body numbers in the Arabidopsis root epidermis are regulated by cell type, developmental stage and cell cycle parameters. J Cell Sci. 1998;111:3687–3694. doi: 10.1242/jcs.111.24.3687. [DOI] [PubMed] [Google Scholar]
- Carmo-Fonseca M, Ferreira J, Lamond AI. Assembly of snRNP-containing coiled bodies is regulated in interphase and mitosis-evidence that the coiled body is a kinetic nuclear structure. J Cell Biol. 1993;120:841–852. doi: 10.1083/jcb.120.4.841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carmo-Fonseca M, Pepperkok R, Carvalho MT, Lamond AI. Transcription-dependent colocalization of the U1, U2, U4/U6 and U5 snRNPs in coiled bodies. J Cell Biol. 1992;117:1–14. doi: 10.1083/jcb.117.1.1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Carrington JC, Freed DD, Leinicke AJ. Bipartite signal sequence mediates nuclear translocation of the plant potyviral NIa protein. Plant Cell. 1991;3:953–962. doi: 10.1105/tpc.3.9.953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chan EKL, Takano S, Andrade LEC, Hamel JC, Matera AG. Structure, expression and chromosomal localization of human p80-coilin gene. Nucleic Acids Res. 1994;22:4462–4469. doi: 10.1093/nar/22.21.4462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chiu W-I, Niwa Y, Zeng W, Hirano T, Kobayashi H, Sheen J. Engineered GFP as a vital reporter in plants. Curr Biol. 1996;6:325–330. doi: 10.1016/s0960-9822(02)00483-9. [DOI] [PubMed] [Google Scholar]
- Christensen AH, Sharrock RA, Quail PH. Maize polyubiquitin genes: structure, thermal perturbation of expression and transcript splicing and promotor activity following transfer to protoplast by electroporation. Plant Mol Biol. 1992;18:675–689. doi: 10.1007/BF00020010. [DOI] [PubMed] [Google Scholar]
- Cremer T, Lichter P, Borden J, Ward DC, Manuelidis L. Detection of chromosome aberrations in metaphase and interphase tumor cells by in situ hybridization using chromosome-specific library probes. Hum Genet. 1988;80:235–246. doi: 10.1007/BF01790091. [DOI] [PubMed] [Google Scholar]
- Drobak BK, Watkins PAC. Inositol(1,4,5)triphos-phate production in plant cells: stimulation by the venom peptides, melittin and mastoparan. Biochem Biophys Res Commun. 1994;205:739–745. doi: 10.1006/bbrc.1994.2727. [DOI] [PubMed] [Google Scholar]
- Ferreira J, Carmo-Fonseca M. The biogenesis of the coiled body during early mouse development. Development. 1995;121:601–612. doi: 10.1242/dev.121.2.601. [DOI] [PubMed] [Google Scholar]
- Ferreira JA, Carmo-Fonseca M, Lamond AI. Differential interaction of splicing snRNPs with coiled bodies and interchromatin granules during mitosis and assembly in daughter nuclei. J Cell Biol. 1994;126:11–23. doi: 10.1083/jcb.126.1.11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fortes P, Lamond AI, Ortin J. Influenza virus NS1 protein alters the subnuclear localization of cellular splicing components. J Gen Virol. 1995;76:1001–1007. doi: 10.1099/0022-1317-76-4-1001. [DOI] [PubMed] [Google Scholar]
- Frey MR, Bailey AD, Weiner AM, Matera AG. Association of snRNA genes with coiled bodies is mediated by nascent snRNA transcripts. Curr Biol. 1999;9:126–135. doi: 10.1016/s0960-9822(99)80066-9. [DOI] [PubMed] [Google Scholar]
- Frey MR, Matera AG. Coiled bodies contain U7 small nuclear RNA and associate with specific DNA sequences in interphase human cells. Proc Natl Acad Sci. 1995;92:5915–5919. doi: 10.1073/pnas.92.13.5915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gall JG, Tsvetkov A, Wu Z, Murphy C. Is the sphere organelle/coiled body a universal nuclear component? Dev Genet. 1995;16:25–35. doi: 10.1002/dvg.1020160107. [DOI] [PubMed] [Google Scholar]
- Habets WJ, Hoet MH, De Jong BAW, Van der Kemp A, van Venrooij WJ. Mapping of B cell epitopes on small nuclear ribonucleoproteins that react with human autoantibodies as well as with experimentally induced mouse monoclonal antibodies. J Immunol. 1989;143:2560–2566. [PubMed] [Google Scholar]
- Ito M, Iwase M, Kodama H, Lavisse P, Komamine A, Nishihama R, Machida Y, Watanabe A. A novel cis-acting element in promoters of plant B-type cyclin genes activates M phase-specific transcription. Plant Cell. 1998;10:331–341. doi: 10.1105/tpc.10.3.331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kambach C, Mattaj IW. Nuclear transport of the U2 snRNP-specific U2B" protein is mediated by both direct and indirect signaling mechanisms. J Cell Sci. 1994;107:1807–1816. doi: 10.1242/jcs.107.7.1807. [DOI] [PubMed] [Google Scholar]
- Koncz C, Schell J. The promotor of TL-DNA gene 5 controls the tissue-specific expression of chimeric genes carried by a novel type of Agrobacterium binary vector. Mol Gen Genet. 1986;204:383–396. [Google Scholar]
- Lafarga M, Hervas JP, Santa-Cruz MC, Villegas J, Crespo D. The accessory body of Cajal in the neuronal nucleus. A light and electron microscopic approach. Anat Embryol. 1983;166:19–30. doi: 10.1007/BF00317942. [DOI] [PubMed] [Google Scholar]
- Lamond AI, Carmo-Fonseca M. The coiled body. Trends Cell Biol. 1993;3:198–204. doi: 10.1016/0962-8924(93)90214-l. [DOI] [PubMed] [Google Scholar]
- Lamond AI, Earnshaw WC. Structure and function in the nucleus. Science. 1998;280:547–553. doi: 10.1126/science.280.5363.547. [DOI] [PubMed] [Google Scholar]
- Li G, Sudlow G, Belmont AS. Interphase cell cycle dynamics of a late-replicating, heterochromatic homogeneously staining region: precise choreography of condensation/decondensation and nuclear positioning. J Cell Biol. 1998;140:975–989. doi: 10.1083/jcb.140.5.975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lichter P, Cremer T, Borden J, Manuelidis L, Ward DC. Delineation of individual human-chromosomes in metaphase and interphase cells by in situ suppression hybridization using recombinant DNA libraries. Hum Genet. 1988;80:224–234. doi: 10.1007/BF01790090. [DOI] [PubMed] [Google Scholar]
- Lyon CE, Bohmann K, Sleeman J, Lamond AI. Inhibition of protein dephosphorylation results in the accumulation of splicing snRNPs and coiled bodies within the nucleolus. Exp Cell Res. 1997;230:84–93. doi: 10.1006/excr.1996.3380. [DOI] [PubMed] [Google Scholar]
- Malatesta M, Zancanaro C, Martin TE, Chan EKL, Amalric F, Lührmann R, Vogel P, Fakan S. Is the coiled body involved in nucleolar functions? Exp Cell Res. 1994;211:415–419. doi: 10.1006/excr.1994.1106. [DOI] [PubMed] [Google Scholar]
- Matera AG. Of coiled bodies, gems, and salmon. J Cell Biochem. 1998;70:181–192. [PubMed] [Google Scholar]
- Mattaj IW. RNA recognition: a family matter? Cell. 1993;73:837–840. doi: 10.1016/0092-8674(93)90265-r. [DOI] [PubMed] [Google Scholar]
- Mattaj IW, De Robertis EM. Nuclear segregation of U2 snRNA requires binding of specific snRNP proteins. Cell. 1985;40:111–118. doi: 10.1016/0092-8674(85)90314-9. [DOI] [PubMed] [Google Scholar]
- Monneron A, Bernhard W. Fine structural organization of the interphase nucleus in some mammalian cells. J Ultrastruct Res. 1969;27:266–288. doi: 10.1016/s0022-5320(69)80017-1. [DOI] [PubMed] [Google Scholar]
- Moreno Diaz de la Espina S, Sanchez-Pina MA, Risueño MC, Medina FJ, Fernández-Gómez ME. The role of plant coiled bodies in the nuclear RNA metabolism. Electron Microsc. 1980;2:240–241. [Google Scholar]
- Ochs RL, Stein TW, Tan EM. Coiled bodies in the nucleolus of breast cancer cells. J Cell Sci. 1994;107:385–399. doi: 10.1242/jcs.107.2.385. [DOI] [PubMed] [Google Scholar]
- Pan Z-Q, Prives C. U2 snRNA sequences that bind U2-specific proteins are dispensable for the function of U2 snRNP in splicing. Genes & Dev. 1989;3:1887–1898. doi: 10.1101/gad.3.12a.1887. [DOI] [PubMed] [Google Scholar]
- Ramon y Cajal SR. Un sencillo metodo de coloracion seletiva del reticulo protoplasmico y sus efectos en los diversos organos nerviosos de vertebrados y invertebrados. Trab Lab Invest Biol. 1903;2:129–221. [Google Scholar]
- Rebelo L, Almeida F, Ramos C, Bohmann K, Lamond. AI, Carmo-Fonseca M. The dynamics of coiled bodies in the nucleus of adenovirus-infected cells. Mol Biol Cell. 1996;7:1137–1151. doi: 10.1091/mbc.7.7.1137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Santama N, Dotti CG, Lamond AI. Neuronal differentiation in the rat hippocampus involves a stage-specific reorganization of subnuclear structure both in vivo and in vitro. Eur J Neurosci. 1996;8:892–905. doi: 10.1111/j.1460-9568.1996.tb01576.x. [DOI] [PubMed] [Google Scholar]
- Scherly D, Boelens W, Dathan NA, van Venrooij WJ, Mattaj IW. Major determinants of the specificity of interaction between small nuclear ribonucleoproteins U1A and U2B" and their cognate RNAs. Nature. 1990;345:502–506. doi: 10.1038/345502a0. [DOI] [PubMed] [Google Scholar]
- Schultz MC. Three structures associated with the nucleolus in male rat germinal cells: round body, coiled body and “nubecula” and general presence of round body at male meiosis. Am J Anat. 1990;189:11–23. doi: 10.1002/aja.1001890103. [DOI] [PubMed] [Google Scholar]
- Seite R, Pebusque MJ, Vio-Cigna M. Argyrophilic proteins on coiled bodies in sympathetic neurons identified by Ag-NOR procedure. Biol Cell. 1982;46:97–100. [Google Scholar]
- Shaw PJ, Beven AF, Leader DJ, Brown JWS. Localization and processing from a polycistronic precursor of novel snoRNAs in maize. J Cell Sci. 1998;111:2121–2128. doi: 10.1242/jcs.111.15.2121. [DOI] [PubMed] [Google Scholar]
- Simpson GG, Clark GP, Rothnie HM, Boelens W, van Venrooij W, Brown JWS. Molecular characterization of the spliceosomal proteins U1A and U2B" from higher plants. EMBO J. 1995;14:4540–4550. doi: 10.1002/j.1460-2075.1995.tb00133.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Simpson GG, Vaux P, Clark G, Waugh R, Beggs JD, Brown JWS. Evolutionary conservation of the spliceosomal protein, U2B". Nucleic Acids Res. 1991;19:5213–5217. doi: 10.1093/nar/19.19.5213. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sleeman J, Lyon CE, Platani M, Kreivi J-P, Lamond AI. Dynamic interactions between splicing snRNPs, coiled bodies and nucleoli revealed using snRNP protein fusions to the green fluorescent protein. Exp Cell Res. 1998;243:290–304. doi: 10.1006/excr.1998.4135. [DOI] [PubMed] [Google Scholar]
- Smith KP, Carter KC, Johnson CV, Lawrence JB. U2 and U1 snRNA gene loci associate with coiled bodies. J Cell Biochem. 1995;59:473–485. doi: 10.1002/jcb.240590408. [DOI] [PubMed] [Google Scholar]
- Zink D, Cremer T. Cell nucleus: chromosome dynamics in nuclei of living cells. Curr Biol. 1998;8:321–324. doi: 10.1016/s0960-9822(98)00198-5. [DOI] [PubMed] [Google Scholar]