

Abstract
Anti-4-1BB and cisplatin showed synergistic anti-cancer effects in the CT-26 colon carcinoma model, producing complete regression in >60% of mice with either preventive or therapeutic treatment. The tumor-free mice formed long-lasting CD8+ T-dependent tumor-specific memory. Anti-4-1BB induced rapid repopulation of T and B cells from cisplatin-mediated lymphopenia and differentiation and expansion of IFN-γ+CD11c+CD8+ T cells. Cisplatin facilitated expansion of naïve, effector, and memory CD8+ T cells; combination therapy produced almost twice as many lymphoid cells as anti-4-1BB alone. Cisplatin increased 4-1BB on antigen-primed T cells and induced 4-1BB de novo on kidney tubular epithelium. Cross-linking of 4-1BB protected the T cells and kidney epithelium from cisplatin-mediated apoptosis by increasing expression of anti-apoptotic molecules. Thus cisplatin-induced 4-1BB provided a mechanism for amelioration of the lymphopenia and nephrotoxicity inherent in cisplatin treatment. We concluded that chemo-immunotherapy with anti-4-1BB and cisplatin is synergistic in tumor killing and prevention of organ-specific toxicity.
Introduction
Immunotherapy can complement anti-cancer chemotherapy (1-6). Agonistic anti-4-1BB (CD137), an immunotherapeutics, displays a strong anticancer effect (7-9) and is undergoing clinical trials against various tumors. Genotoxic agents, γ-irradiation and chemotherapeutics, increased 4-1BB expression on T cells, and cross-linking of the 4-1BB enhanced T-cell survival (10). We, therefore, hypothesized that the chemotherapeutics which enhanced 4-1BB expression could potentiate the anti-cancer activities of an agonistic anti-4-1BB.
Cisplatin is widely used as a chemotherapeutic agent to treat a variety of tumors (11). The efficacy of cisplatin is dose-dependent, although it induces thrombocytopenia and leukocytopenia (12). Nephrotoxicity frequently hinders the use of higher doses that would maximize its antineoplastic effects (13, 14).
Here, we investigated whether anti-4-1BB augmented the anti-cancer effect of cisplatin and ameliorated cisplatin-induced loss of leukocytes and renal dysfunction. The results revealed that combination therapy with agonistic anti-4-1BB and cisplatin enhanced primary and memory anti-tumor immune responses, which were mediated by a preferential enhancement of antigen-specific CD8+ T cells, especially CD11c+CD8+ T cells. Moreover, anti-4-1BB treatment attenuated cisplatin-induced nephrotoxicity by protecting renal tubular epithelial cells from cisplatin-induced apoptosis.
Materials and Methods
Mice
Six to eight week-old BALB/c, C57BL/6, and OT-1 mice (Jackson Laboratory, Bar Harbor, ME), and Rag2γc-/- (CIEA, Japan) were housed in a specific pathogen-free facility at the Immunomodulation Research Center (IRC) at the University of Ulsan. All animal experiments were performed in accordance with the approved protocols and guidelines of the IRC’s Institutional Animal Care and Use Committee.
Antibodies and peptides
Anti-m4-1BB hybridoma cells (3E1) were a gift from Dr. Robert Mittler (Emory University, Atlanta, GA). Anti-mCD4 (GK1.5) and anti-mCD8 (2.43) hybridomas were obtained from ATCC (Manassas, VA). Purified rat IgG was obtained from Sigma-Aldrich (St Louis, MO). Flow cytometric mAbs were purchased from BD PharMingen (San Diego, CA). H-2Kb/OVA257-264-PE pentamers were purchased from ProImmune Ltd. (Oxford, United Kingdom). OVA257-264 (SIINFEKL) was synthesized by Peptron (Dae-Jeon, Korea).
Tumor challenge and treatment
Mice were injected s.c. with 3 × 105 CT26 cells on the dorsal surface (PI day 0). As a standard treatment, tumor-challenged mice were injected i.p. with 100 μg of rat IgG or anti-4-1BB mAb (3E1) every 5 days for five times, and a single injection of 10 mg/kg cisplatin on PI day 0. To examine therapeutic effects of combined therapy, the mice received the standard therapy beginning 6 days PI. For tumor-rechallenge, tumor-free mice were injected s.c. in the opposite flank 100 days after the primary tumor inoculation with CT26 or RENCA cells (3, 6, or 12 × 105 cells / mouse). Mice were injected i.p. with 400 μg of anti-CD4 or anti-CD8, or 100 μg of anti-asialo-GM1 antibody every 5 days for five cycles. The efficacy of immune subset depletion (>99 %) was confirmed by flow cytometry.
Flow cytometry
Lymphocytes were incubated with the Fc blocker 2.4G2 for 10 minutes at 4°C and stained with specific antibodies to surface markers. To identify IFN-γ-producing cells, TDLN (tumor draining lymph node) cells were first stimulated with 50 ng/ml PMA and 500 ng/ml ionomycin for 6 hours in a medium containing Brefeldin A. Cells were then stained with FITC-anti-CD8 or anti-CD4, and intracellularly stained PE-anti-IFN-γ by using Cytofix/cytoperm kit. All samples were analyzed using a FACScalibur (BD Biosciences, San Jose, CA).
Assessment of renal function
Blood samples were obtained from before (0 h) and at 24, 48, and 72 h after treatment with cisplatin and/or anti-4-1BB mAb. Serum creatinine was measured as a marker of renal dysfunction using a Roche Cobas Rara automated system (Roche, Nutley, NJ) with a Creatinine kit (R&D Systems, Minneapolis, MN).
Histological examination
The kidney paraffin block was cut into 4 μm sections and stained with haematoxylin-eosin (H&E). Renal tubular damage was assessed using a semiquantitative scale. A pathologist who was blinded to the experiments scored the degree of tubular damage. The magnitude of tubular epithelial cell loss and necrosis was scored on the basis of the percentage of affected tubules in a high-power field under a light microscope, as follows: 0, normal; 0.5, < 10%; 1, 10 to 25%; 2, 25 to 50%; 3, 50 to 75%; and 4, > 75%. For anti-4-1BB immunostaining of the kidney, thin cryosections from kidneys were stained with FITC-anti-m4-1BB (3E1) mAb, and visualized with an Olympus FV500 confocal laser scanning microscope.
RT-PCR
Total RNA was extracted from cells and tissues using TRIzol (Invitrogen Life Technologies) and reverse-transcribed into cDNA using a SuperScriptII™ (Invitrogen Life Technologies). Gene-specific primers used were as follows: mouse 4-1BB, forward 5′-tgacatttcgccatgggaaacaactgt-3′, reverse 5′-tcccggtcttaagcacagaccttccgt-3′; human 4-1BB, forward 5′-cgcgtggatccacaagatcattgcaggatcc-3′, reverse 5′-ttgagctcgaggatctgcggagagtgtcctg-3′; GAPDH, forward 5′-gaacgggaagcttgtcatcaa-3′, reverse 5′-ctaagcagttggtggtgcag-3′. PCR conditions were as follows: for 4-1BB, 94°C for 5 min, followed by 35 cycles of 30 sec at 94°C, 30 sec at 55°C, and 1 min at 72°C with a final extension at 72°C for 5 min; for the GAPDH RT-PCR, conditions were identical except that 30 cycles were used instead of 35.
Real-time PCR
SYBR Green I-based real-time quantitative PCR was carried out on a continuous fluorescence detection system (Opticon DNA Engine, MJ Research Inc., Waltham, MA) with first-strand cDNA of kidney. Gene-specific primers used were as follows: mouse Bcl-2, forward 5′-tgggatgcctttgtggaactat-3′, reverse 5′-agagacagccaggagaaatcaaac-3′; mouse Bcl-XL, forward 5′-ctgggacacttttgtggatctct-3′, reverse 5′-gaagcgctcctggccttt-3′; mouse Bfl-1, forward 5′-gcagattgccctggatgtat-3′, reverse 5′-agaaaagtcagccagccacta-3′; mouse Bax, forward 5′-gagatgaactggacagcaat-3′, reverse 5′-atcccaggaaaatgtcataa-3′; GAPDH, forward 5′-gaacgggaagcttgtcatcaa-3′, reverse 5′-ctaagcagttggtggtgcag-3′. The relative gene expression was determined using the comparative CT method and normalized to the housekeeping gene GAPDH. To simplify the representation of Bcl-2, Bcl-XL, Bfl-1, and Bax expression, the gene expression data normalized for GAPDH are shown as fold increases compared to levels in the rat IgG group and are means ± SD of triplicate experiments.
Kidney epithelial cells
Kidneys were removed, sliced, and digested with collagenase type IV in Hank’s balanced salt solution for 30 min at 37°C. The digest was centrifuged, and the cell pellet was resuspended in a cold culture medium with 10% FBS. The cell suspension was plated onto tissue culture dishes. Cells were allowed to adhere for 2 h. Unbound cells were removed by washing three times with PBS. Only adherent cells were maintained as primary cultures at 37°C for 7 days; these cells were then used for experiments.
Western blot
Samples of 20 μg kidney protein were diluted with 4 × SDS sample buffer, applied to SDS-PAGE gels, fractionated, and transferred to nitrocellulose membranes (Millipore, Bedford, MA). The membranes were incubated with anti-4-1BB mAb (3E1) and anti-rat IgG-horseradish peroxidase (HRP), and bound Ab was detected by enhanced chemiluminescence (ECL) (Amersham Pharmacia Biotech, Little Chalfont, U.K.).
SiRNA transfection and treatment
The cultured kidney epithelial cells from Rag2γc- or 4-1BB-deficient mice were transfected with either gene-specific siRNA against Bcl-XL, Bfl-1, and Bax or nontargeting control siRNA according to the manufacturer’s directions (Santa Cruz Biotechology). Transfection efficiencies were assessed by RT-PCR analysis 36 h after gene transfer. Two days after the transfection, cells were treated with 2 μg/ml cisplatin and/or 10 μg/ml rat IgG or anti-4-1BB. The absolute numbers of viable cells were measured in primary cultures of mouse kidney epithelial cells at the indicated time points.
Statistics
Student’s t test and one-way ANOVA were performed using the Prism software program (GraphPad Software).
Results and Discussion
We screened a variety of anti-cancer chemotherapeutics to select one that enhanced anti-cancer activity of the agonistic anti-4-1BB. A synergistic anti-cancer effect was observed when cisplatin was used in combination with anti-4-1BB in B16 melanoma and CT26 carcinoma models. Mice receiving combination therapy seldom showed the hunched posture, ruffled fur, and diarrhea often seen with cisplatin alone.
We examined mechanisms involved in this anti-cancer synergy and reduced nephrotoxicity in the CT26 model. Mice were treated with anti-4-1BB and/or cisplatin beginning on PI (post-tumor injection) day 0 or 6: >50% of 4-1BB-treated mice showed delayed tumor growth and 10-30% were tumor-free, whereas rat IgG- or cisplatin-alone-treated mice were dead or sacrificed by PI day 35 due to the large tumor burden (Fig. 1A and B). Anti-4-1BB plus cisplatin resulted in 60-70% tumor-free mice in both preventive and therapeutic models. Mice tumor-free for >100 days after combination treatment were re-challenged with CT26 or RENCA cells. CT26 did not grow, whereas RENCA did (Fig. 1C). Depletion of CD8+ T cells from the tumor-free mice followed by re-challenge with CT26 produced tumor growth; depletion of CD4 or NK cells did not (Fig. 1C). These results suggest that cisplatin enhances 4-1BB-mediated anti-cancer effects and that combination therapy leads to CD8+ T cell-mediated long lasting memory. Anti-4-1BB increased the numbers of IFN-γ+CD4+ and CD8+ T cells including AH-1-specific CD8+ T cells, and cisplatin provided further enhancement (Fig. 1D, and Supplementary Fig. 1). CD11c+CD8+ T cells (9) were the major source of IFN-γ-producing CD8+ T cells (Fig. 1D and Supplementary Fig. 2).
Figure 1.

Combination therapy with anti-4-1BB and cisplatin synergistically induces anti-tumor immune responses. (A) BALB/c mice were challenged s.c. with 3 × 105 CT26 colon carcinoma cells, received a single i.p. injection of 200 μg cisplatin, and injected i.p. with 100μg of rat IgG or anti-4-1BB mAb (3E1) every 5 days for five times from PI day 0. (B) Six days after tumor challenge, mice were treated i.p with PBS or 200 μg cisplatin and injected with rat IgG or anti-4-1BB mAb every 5 days for five times. The mice were monitored everyday for tumor growth and survival. (C) Tumor-free mice from the anti-4-1BB- or combination-treated groups were rechallenged s.c. with the indicated number of CT26 or RENCA cells on PI day 100 and monitored for tumor growth. In separated experiment, tumor-free mice on PI day 100 were challenged s.c. with CT26 and treated i.p. with 100μg of anti-asialo-GM1, 400μg of GK1.5, or 400μg of 2.43 mAb to deplete NK, CD4+ T, or CD8+ T cells every 5 days for five times. (D) On PI day 17, single cell suspensions of TDLNs were stained with PE-anti-CD11c and PE-Cy5-anti-CD8. TDLN cells were also stained with FITC-anti-CD4 or -anti-CD8, and further intracellularly stained with PE-anti-IFN-γ, and subsequently analyzed using FACscan (BD Biosciences). Absolute numbers were calculated by multiplying percentages measured by flow cytometry by total numbers of viable cells. Each group contained 10 mice (A-B) or 5 mice (C-D). The results are representative of two independent experiments. All plotted data are means ± SD.
Combination therapy showed enhanced expansion of all three phenotypes of CD8+ (Fig. 2A) and CD4+ T cells (Supplementary Fig. 3) — naïve (CD62LhighCD44low), effector (CD62LlowCD44high), and memory (CD62LlowCD44high) — compared with other monotherapies. Anti-4-1BB alone expanded effector and memory CD8+ T cells significantly compared with IgG or cisplatin alone. 4-1BB effectively expanded T cells following cisplatin-induced lymphopenia and expansion after combination therapy was more vigorous than with 4-1BB alone (Fig. 2A). Reduction of Foxp3+ Treg cells in TDLN and spleen was observed at 2-7 days following cisplatin-treatment (Supplementary Fig. 4). This may contribute to the cisplatin-mediated anti-cancer effect.
Figure 2.

Treatment with anti-4-1BB rescues lymphocytes from cisplatin-mediated apoptosis. Tumor-challenged BALB/c mice were injected i.p. with rat IgG or anti-4-1BB mAb as well as single injection of cisplatin on PI day 0 as described above. On PI day 17, single cell suspensions of TDLNs were stained with FITC-anti-CD62L, PE-anti-CD44, and PE-Cy5-anti-CD8 (A). (B) Five days after the treatment, TDLN cells were counted and stained with FITC-anti-CD3, -anti-B220, -anti-CD4 or -anti-CD8, and further stained with PE-anti-Annexin-V to examine the apoptotic cells. Samples were subsequently analyzed using FACscan and absolute numbers of annexin V-negative cells were calculated as described above. (C) To create a system that determine the 4-1BB expression on both Ag-specific and Ag-nonspecific CD8+ T cells simultaneously, OVA-specific CD8+ T cells were isolated from OT-I mice and adoptively transferred into C57BL/6 mice (5 × 106 cells per mouse). The mice were immunized s.c. with 10 μg of OVA in incomplete Freund’s adjuvant (IFA), and injected i.p. with 200 μg of cisplatin (filled symbol) or PBS (open symbol) as a control. Draining lymph node (DLN) cells were stained with FITC-anti-4-1BB, PE-Kb/OVA tetramer, and PE-Cy5-anti-CD8 on the indicated days. Data dictated the percentage of 4-1BB+CD8+ T cells in gated Kb/OVA tetramer-negative or Kb/OVA tetramer-positive CD8+ T cells. (D) Total RNA was extracted from CD8+ T cells of four different groups of mice on PI day 5 and first-strand cDNA was synthesized with 0.5 μg total RNA. SYBR Green I-based real-time quantitative PCR was carried out on a continuous fluorescence detection system (Opticon DNA Engine, MJ Research Inc, Waltham, MA). Relative gene expression was determined using the comparative CT method and normalized to the housekeeping gene GAPDH. To simply represent Bcl-2, Bcl-XL, Bfl-1, and Bax expression, the gene expression data normalized for GAPDH are shown as the fold increase compared to levels in the rat IgG-treated group. Each group contained three mice and the results are representative of two independent experiments. All plotted data are means ± SD (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
Absolute numbers of viable cells in the TDLNs on PI day 5 showed that cisplatin reduced total lymphoid cells by >78%, T cells >70%, and B cells >90% (Fig. 2B). Combination therapy enhanced lymphoid cells by PI day 5 compared with cisplatin alone: total numbers of TDLN cells increased >11-fold, T cells >7-fold (CD4, >5-fold; CD8, >11-fold), and B cells >54-fold. Combination therapy was also more effective in repopulating lymphoid cells than anti-4-1BB alone (total cells, 1.7-fold; B cells, 2.5-fold) (Fig. 2B). Analysis of TDLN cells on PI day 17 showed a 4-fold increase in CD8+ T cells and a 2-fold increase in CD4+ T cells with combination therapy, compared to rat IgG treatment (Supplementary Fig. 5).
We hypothesized that cisplatin enhanced the expression of 4-1BB on the T cell surface, cross-linking of which produced more anti-apoptotic molecules, resulting in increased lymphocytes after combination therapy. To create a system that measure 4-1BB expression in both antigen-primed and naïve CD8+ T cells simultaneously, OT-1 CD8+ T cells were adoptively transferred into congenic C57BL/6 mice, which were treated with OVA and/or cisplatin. Cisplatin increased expression of 4-1BB on OVA/H2Kb-positive CD8+ T cells from day 1 of antigen-injection; the higher levels persisted until day 6-8, in which 4-1BB in cisplatin plus OVA-treated mice approximately 2-fold higher than in OVA alone group (Fig. 2C). In separate experiments, we showed that enhanced release of tumor antigen by cisplatin did not contribute to the enhanced expression of 4-1BB and expansion of CD8+ T cells (Supplementary Fig. 6).
Interestingly, cross-linking of 4-1BB in the presence of cisplatin increased expression of Bcl-2 and Bfl-1 almost 2-fold compared with anti-4-1BB alone. Expression of Bax, a molecule known to induce cisplatin-mediated apoptosis of kidney epithelium (15), did not change (Fig. 2D). Cisplatin-mediated enhancement of 4-1BB expression on the T cell surface and concomitant enhancement of anti-apoptotic molecules with anti-4-1BB treatment may contribute to the rapid expansion of T cells and enhanced tumor killing activity.
Cisplatin increased renal tubular damage dose-dependently; damage was limited and localized at 200 μg, but widespread in all parts of the cortex at 400 or 800 μg. However, high-dose tubular damage was ameliorated with anti-4-1BB (Supplementary Fig. 7). Combination treatment reduced the semiquantitative scores for renal tubular epithelial cell necrosis (Supplementary Fig. 8) and loss by 2-fold compared with cisplatin alone (Fig. 3A). The concentration of serum creatinine increased dose-dependently with cisplatin and decreased with addition of anti-4-1BB (Fig. 3B).
Figure 3.

4-1BB triggering ameliorates cisplatin-mediated nephrotoxicity. BALB/c mice were injected i.p. with various doses of cisplatin and simultaneously injected with rat IgG or anti-4-1BB mAb. (A) Semiquantification of mouse renal tubular damage. Mouse kidneys at 72hrs after cisplatin injection were embedded in paraffin, cut into 4 mm sections, and stained with haematoxylin-eosin (H&E). Renal tubular damage was assessed using a semiquantitative scale. A pathologist who was blinded to the experiments scored the degree of tubular damage. The magnitude of tubular epithelial cell necrosis and loss was scored on the basis of the percentage of affected tubules in a high-power field under a light microscope, as follows: 0, normal; 0.5, < 10%; 1, 10 to 25%; 2, 25 to 50%; 3, 50 to 75%; and 4, > 75%. (B) Blood samples were obtained from before (0 hour) and at 24, 48, and 72 h after treatment with cisplatin and/or anti-4-1BB mAb. Serum creatinine was measured as a marker of renal dysfunction using a Roche Cobas Rara automated system (Roche, Nutley, NJ) with a Creatinine kit (R&D Systems, Minneapolis, MN). (C) 4-1BB expression in renal tubular epithelial cells. I. Rag2γc- or 4-1BB-deficient mice received a single injection of 100, 200 or 400 μg cisplatin. RT-PCR for 4-1BB was performed with cDNA of kidney cells 24 hr after cisplatin administration. II. 4-1BB proteins were detected in kidney samples by performing western blotting with anti-4-1BB mAb (3E1 clone). (D) For anti-4-1BB immunostaining of the kidney, frozen sections (4 μm thickness) were prepared from kidney 24 h after cisplatin administration and stained with FITC-conjugated anti-m4-1BB mAb. Sections were viewed and photographed using a laser-scanning confocal fluorescence microscope system (Fluoview FV500 Olympus, Center Valley, PA; original magnification ×10). Each group contained three mice and the results are representative of three independent experiments. All plotted data are means ± SD (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
To explain 4-1BB-mediated amelioration of cisplatin-induced nephrotoxicity, expression of 4-1BB (16) was examined in renal tissue following cisplatin treatment in ex vivo. To minimize infiltration of 4-1BB-expressing immune cells into the kidney, cisplatin was injected into Rag2γc DKO mice at doses indicated. 4-1BB was detected only in the kidneys from mice receiving 400 μg or more cisplatin, but not in those receiving lower doses (Fig. 3C-I and -II). 4-1BB-positive cells were located in the kidney cortex where renal tubular damage was increased by cisplatin (Fig. 3D).
4-1BB expression was detected in cisplatin-treated in vitro cultured kidney epithelium, but not in PBS-treated cells (Fig. 4A). Human embryonic kidney cells (HEK 293) constitutively expressed 4-1BB, which was marginally increased by cisplatin (Supplementary Fig. 9). We tested whether cross-linking of 4-1BB on cultured kidney epithelium enhanced the expression of anti-apoptotic molecules as with T cells (17, 18). Cisplatin decreased mRNA of anti-apoptotic molecules such as Bcl-2, Bcl-XL, and Bfl-1, only the decrease of Bfl-1 mRNA was statistically significant. Conversely, combination with anti-4-1BB significantly increased the expression of Bfl-1. The pro-apoptotic gene Bax increased with cisplatin treatment and decreased with anti-4-1BB in vivo (Fig. 4B).
Figure 4.
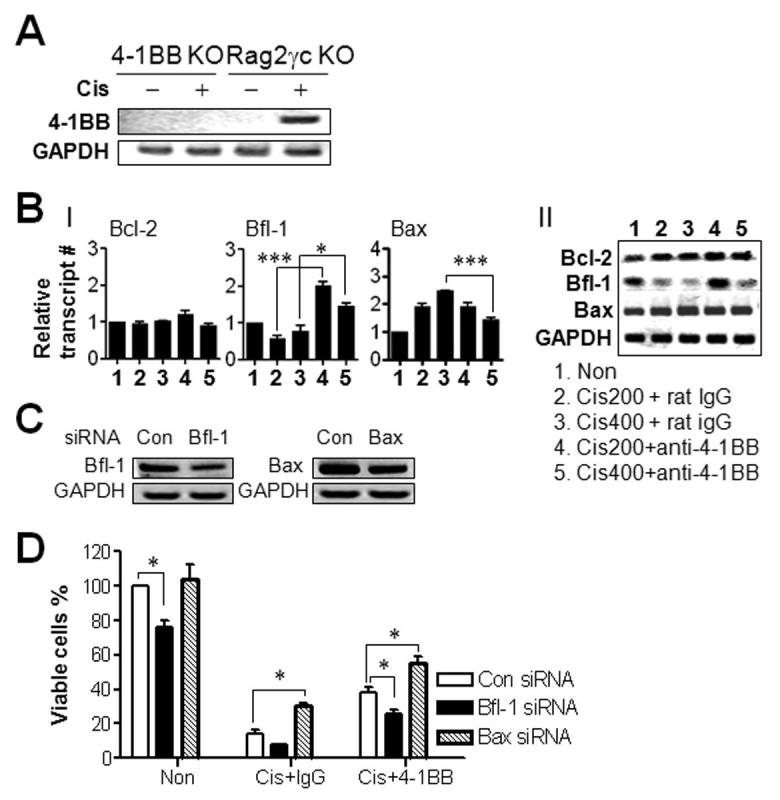
4-1BB triggering enhances survival of kidney epithelium in vivo. (A) Kidney epithelial cells from 4-1BB KO BALB/c or Rag2γc KO mice were cultured in vitro for 7 days and treated with 2 μg/ml cisplatin or PBS for 24hrs. RT-PCR was performed with cDNA of the cultured cells to determine 4-1BB transcripts. (B) Rag2γc KO mice were injected i.p. with indicated doses of cisplatin and also single-injected with 100μg of rat IgG or anti-4-1BB mAb. Complementary DNA was synthesized with total RNA from kidney 72 hrs after cisplatin injection. Real-time PCR (I) and RT-PCR (II) were performed to assess Bcl-2, Bfl-1, and Bax expression as describe above. Relative gene expression was normalized to GAPDH and shown as the fold increase compared to that of rat IgG-treated group. (C, D) Kidney epithelial cells from Rag2γc DKO or 4-1BB KO mice were transfected with either gene-specific siRNA against Bfl-1, Bax or nontargeting control siRNA according to manufacturer’s instruction (Santa Cruz Biotechology). (C) Decrease in Bfl-1 or Bax mRNA upon corresponding siRNA transfection was determined by RT-PCR. (D) Cells were treated with 2 μg/ml cisplatin and/or 10 μg/ml of rat IgG or anti-4-1BB, and the absolute numbers of viable cells were determined at the indicated time points. Each group contained three mice and the results are representative of three independent experiments. All plotted data are means ± SD (*, P < 0.05; **, P < 0.01; ***, P < 0.001).
We tested whether the 4-1BB-mediated survival of kidney epithelium was related to the changes in Bfl-1 and Bax expression levels using gene-specific small interfering RNA (siRNA). Transfection of siRNA specific to Bfl-1 or Bax decreased mRNA levels of Bfl-1 or Bax by 40 to 50% of control siRNA in primary cultures of kidney epithelium (Fig. 4C). Bfl-1 siRNA reduced anti-4-1BB-mediated survival of kidney epithelium by 32.5% compared with the control, whereas Bax siRNA enhanced the survival of kidney epithelium by 2.14-fold compared with cisplatin alone and by 1.46-fold compared with cisplatin plus anti-4-1BB (Fig. 4D). We concluded that cisplatin directly enhances 4-1BB expression in renal tubular epithelial cells, which is involved in the anti-4-1BB-mediated protection of kidney epithelial cells by regulating the expression of anti-apoptotic and pro-apoptotic molecules.
It appears that ciplatin does not produce any tumor-killing activities against CT26 (Fig. 1); the therapeutic effects of anti-4-1BB plus cisplatin were completely dependent upon both primary and secondary anti-tumor activities of CD8+ T cells. This suggests that cisplatin rather preconditions the host, possibly dampening the immunosuppressive network, and providing the necessary lymphoid space and cytokine milieu for augmented 4-1BB-mediated expansion and differentiation of CD8+ T cells.
It is noteworthy that B cells were the most sensitive to cisplatin toxicity and anti-4-1BB was very effective in repopulating B cells (Fig. 2). Anti-4-1BB may cross-link 4-1BB on the B cell surface (19) or alternatively repopulated CD4+ T cells may provide growth factors for B cells. Cisplatin enhanced 4-1BB expression on antigen-primed CD8+ T cells and anti-4-1BB markedly enhanced the repopulation of lymphocytes (Fig. 2). Because 4-1BB is induced in an antigen-specific manner, the majority of repopulated T cells are likely against tumor antigens. Although we could not directly quantify the tumor-specific CD8+ T cells in current studies because of the unavailability of CT26-specific tetramers, we found that, in another tumor model (B16 melanoma), the major portion of the CD8+ T cells repopulated by anti-4-1BB were tumor-antigen-specific (unpublished observation).
It is a novel finding that cisplatin induced 4-1BB on the kidney epithelium (Fig. 3). Provision of the anti-apoptotic signals to renal tubular epithelium by anti-4-1BB seemed to contribute to the improvement in cisplatin-induced nephrotoxicity. Both enhancement of Bfl-1 and suppression of Bax (Fig. 4) appear important factors involved in anti-4-1BB-mediated amelioration of nephrotoxicity.
Current studies suggest that one single type of chemotherapy or immunotherapy may not be able to deal with all facets of tumor progression, and tumor- and immune system derived regulatory barriers. Therefore, a rational combination of synergistic chemotherapy and immunotherapy could provide a successful clinical approach to human cancers.
Supplementary Material
Acknowledgments
This work was supported in part by grants from the National Cancer Center, Korea(NCC-0810720-1); Korean Research Foundation (KRF-2005-201-E00008, KRF-2005-084-E00001 to BSK); Arthritis Foundation (Innovative Research Award to BSK); and NIH Grant R01EY013325 (BSK).
Footnotes
Completing interests statement
The authors declare that they have no competing financial interests.
References
- 1.Nowak AK, Lake RA, Marzo AL, et al. Induction of tumor cell apoptosis in vivo increases tumor antigen cross-presentation, cross-priming rather than cross-tolerizing host tumor-specific CD8 T cells. J Immunol. 2003;170(10):4905–13. doi: 10.4049/jimmunol.170.10.4905. [DOI] [PubMed] [Google Scholar]
- 2.Nowak AK, Robinson BW, Lake RA. Synergy between chemotherapy and immunotherapy in the treatment of established murine solid tumors. Cancer Res. 2003;63(15):4490–6. [PubMed] [Google Scholar]
- 3.Bergmann-Leitner ES, Abrams SI. Treatment of human colon carcinoma cell lines with anti-neoplastic agents enhances their lytic sensitivity to antigen-specific CD8+ cytotoxic T lymphocytes. Cancer Immunol Immunother. 2001;50(9):445–55. doi: 10.1007/s002620100229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dudley ME, Wunderlich JR, Robbins PF, et al. Cancer regression and autoimmunity in patients after clonal repopulation with antitumor lymphocytes. Science. 2002;298(5594):850–4. doi: 10.1126/science.1076514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Ghiringhelli F, Larmonier N, Schmitt E, et al. CD4+CD25+ regulatory T cells suppress tumor immunity but are sensitive to cyclophosphamide which allows immunotherapy of established tumors to be curative. Eur J Immunol. 2004;34(2):336–44. doi: 10.1002/eji.200324181. [DOI] [PubMed] [Google Scholar]
- 6.Lake RA, Robinson BW. Immunotherapy and chemotherapy--a practical partnership. Nat Rev Cancer. 2005;5(5):397–405. doi: 10.1038/nrc1613. [DOI] [PubMed] [Google Scholar]
- 7.Kim JA, Averbook BJ, Chambers K, et al. Divergent effects of 4-1BB antibodies on antitumor immunity and on tumor-reactive T-cell generation. Cancer Res. 2001;61(5):2031–7. [PubMed] [Google Scholar]
- 8.Wilcox RA, Flies DB, Zhu G, et al. Provision of antigen and CD137 signaling breaks immunological ignorance, promoting regression of poorly immunogenic tumors. J Clin Invest. 2002;109(5):651–9. doi: 10.1172/JCI14184. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Choi BK, Kim YH, Kang WJ, et al. Mechanisms involved in synergistic anticancer immunity of anti-4-1BB and anti-CD4 therapy. Cancer Res. 2007;67(18):8891–9. doi: 10.1158/0008-5472.CAN-07-1056. [DOI] [PubMed] [Google Scholar]
- 10.Kim KM, Kim HW, Kim JO, Baek KM, Kim JG, Kang CY. Induction of 4-1BB (CD137) expression by DNA damaging agents in human T lymphocytes. Immunology. 2002;107(4):472–9. doi: 10.1046/j.1365-2567.2002.01538.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lebwohl D, Canetta R. Clinical development of platinum complexes in cancer therapy: an historical perspective and an update. Eur J Cancer. 1998;34(10):1522–34. doi: 10.1016/s0959-8049(98)00224-x. [DOI] [PubMed] [Google Scholar]
- 12.Ohno S, Strebel FR, Stephens LC, et al. Haematological toxicity of carboplatin and cisplatin combined with whole body hyperthermia in rats. Br J Cancer. 1993;68(3):469–74. doi: 10.1038/bjc.1993.372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Schrier RW. Cancer therapy and renal injury. J Clin Invest. 2002;110(6):743–5. doi: 10.1172/JCI16568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Ries F, Klastersky J. Nephrotoxicity induced by cancer chemotherapy with special emphasis on cisplatin toxicity. Am J Kidney Dis. 1986;8(5):368–79. doi: 10.1016/s0272-6386(86)80112-3. [DOI] [PubMed] [Google Scholar]
- 15.Wei Q, Dong G, Franklin J, Dong Z. The pathological role of Bax in cisplatin nephrotoxicity. Kidney Int. 2007;72(1):53–62. doi: 10.1038/sj.ki.5002256. [DOI] [PubMed] [Google Scholar]
- 16.Lim HY, Kim KK, Zhou FC, Yoon JW, Hill JM, Kwon BS. 4-1BB-like molecule is expressed in islet-infiltrating mononuclear cells and in the gray matter of the brain. Cell Biol Int. 2002;26(3):271–8. doi: 10.1006/cbir.2001.0847. [DOI] [PubMed] [Google Scholar]
- 17.Lee HW, Park SJ, Choi BK, Kim HH, Nam KO, Kwon BS. 4-1BB promotes the survival of CD8+ T lymphocytes by increasing expression of Bcl-xL and Bfl-1. J Immunol. 2002;169(9):4882–8. doi: 10.4049/jimmunol.169.9.4882. [DOI] [PubMed] [Google Scholar]
- 18.Lee HW, Nam KO, Seo SK, Kim YH, Kang H, Kwon BS. 4-1BB cross-linking enhances the survival and cell cycle progression of CD4 T lymphocytes. Cell Immunol. 2003;223(2):143–50. doi: 10.1016/s0008-8749(03)00169-2. [DOI] [PubMed] [Google Scholar]
- 19.Schwarz H, Valbracht J, Tuckwell J, von Kempis J, Lotz M. ILA, the human 4-1BB homologue, is inducible in lymphoid and other cell lineages. Blood. 1955;85(4):1043–52. [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.