

Abstract
In contrast to early theories of socialization which emphasized the role of parents in shaping their children's personalities, recent empirical evidence suggests an evocative relationship between adolescent personality traits and the quality of the parent-adolescent relationship. Research using behavior genetic methods suggest that the association between personality and parenting is genetically mediated, such that the genetic effects on adolescent personality traits overlap with the genetic effects on parenting behavior. In the current study, we examined whether the etiology of this relationship might change depending on the adolescent's personality. Biometrical moderation models were utilized to test for gene-environment interaction and correlation between personality traits and measures of conflict, regard, and involvement with parents in a sample of 2,452 adolescents (M age=17.79). We found significant moderation of both positive and negative qualities of the parent-adolescent relationship, such that the genetic and environmental variance in relationship quality varied as functions of the adolescent's levels of personality. These findings support the importance of adolescent personality in the development of the quality of the parent-adolescent relationship.
Keywords: personality, parenting, behavior genetics, moderation
For many years, it was assumed that family environment, including parenting quality, played a causal role in personality development. This theory of socialization maintained that parents played the major, if not defining, role in child development (Bell, 1968). Subsequent to this, the dynamic interactionistic paradigm recognized that children were not simply the products of parental behavior (Caspi & Shiner, 2006; Magnusson, 1990; Patterson, 1982; Sameroff, 1983), but that individual differences in childhood personality also lead to variations in the quality of the parent-child relationship. Some took this position to its extreme, suggesting that parents have little if any impact on adolescent development (Harris, 1995, 1998). A reasonable synthetic perspective is that child personality and parental behavior are related through “bidirectional interactive processes” (Collins, Maccoby, Steinberg, Heatherington, & Bornstein, 2000, p.222). In the current study, we examine whether the etiology of the parent-adolescent relationship changes depending on the adolescent's personality by determining the moderating impact of adolescent personality on the genetic and environmental influences on the parent-adolescent relationship.
The Role of Adolescent Personality in the Parent-Adolescent Relationship
Evidence from the literature on personality development supports the notion of temperament (Rothbart & Bates, 2006), consisting of core traits (Asendorpf & Van Aken, 2003) or basic dispositions (McCrae & Costa, 1999) that are present from birth and have links to adult personality (Caspi et al., 2003). This is not to say, however, that personality is “set” from birth. There are several mechanisms by which individual characteristics transact with the environment, including interpersonal relationships (Shiner & Caspi, 2003). On of the most important relationships for personality development is the parent relationship. Here, evidence supports a bidirectional influence between temperament/personality and the parent-child relationship. Negative, inappropriate, or unskilled parenting variables appear to play a particularly important role in the development of externalizing behaviors, while warm and supportive parenting behaviors seem to act as protective factors (Bates, Petit, Dodge, & Ridge, 1998; Belsky, Hsieh, & Crnic, 1998; Rubin, Burgess, Dwyer, & Hastins, 2003; Stoolmiller, 2001). Conversely, children with temperaments high in negative emotionality elicit less supportive parenting behaviors, particularly in low-SES households (Paulussen-Hoogeboom, Stams,Hermanns, & Peetsma, 2007).
Shiner and Caspi (2003) have delineated six environmental processes which work to shape the development of personality through adolescence into adulthood: learning processes, environmental elicitation, environmental construal, social and temporal comparisons, environmental selection, and environmental manipulation. Theoretically, all of these processes could play a part in the emerging relationship between adolescent personality traits and relationships with their parents. There is little empirical evidence for the operation of some (e.g., social comparisons, environmental selection and manipulation), but greater support for the ways in which personality traits may shape the elicitation and construal of behavior from others (Shiner & Caspi, 2003). Particularly as children mature into adolescence, individual differences in personality will evoke different responses from parents and selection of different types and frequencies of interactions with parents. It is most likely this process of person-environment transaction that contributes to the increasing stability and consistency of personality during adolescence (Roberts & DelVecchio, 2000).
Although there are a greater number of studies connecting personality to other forms of interpersonal relationships (e.g., peers and romantic partners), accumulating empirical evidence is supportive of an evocative relationship between adolescent personality and parent behaviors. Branje, van Lieshout, and van Aken (2004) found a positive, cross-sectional link between adolescents’ agreeableness and perceived support from parents. Levels of conscientiousness and openness in the older of two siblings was related to perceived support from both parents, while extraversion and neuroticism in the younger siblings was related to perceived support from dad only. In a later study, Branje, van Lieshout, and van Aken (2005) examined the relationship between agreeableness and perceived support across all possible combinations of family relationships for parents and adolescents. They found no general link across family members between self-reported agreeableness and perceived support; however, they did find significant agreeableness-support correlations within most of the possible family dyads (i.e., mother-father, father-oldest child) except for the mother–younger adolescent relationship. Consistent findings for a link between personality and perceived parental support in older siblings suggest that this association becomes stronger over the course of adolescence.
Longitudinal studies provide a better method than cross-sectional studies for teasing apart causal influences in the personality-parenting association. Asendorpf and Wilpers (1998) examined the longitudinal connections between personality traits and interpersonal relationships in a German college student sample. Overall, they found strong stability of personality over a period of 18 months, compared with much greater instability of social relationships. Personality traits influenced social relationships, but not vice versa; specific to family, conscientiousness was related to frequency of contact with parents. Further, change in relationship status was unrelated to changes in personality. The authors concluded that by early adulthood, the quality of relationships with peers, family, and romantic partners is a function of personality. To examine whether personality in adolescence might be more malleable to influences from relationships, Asendorpf and van Aken (2003) studied the association between personality and parenting behavior between the ages of 12 and 17. Adolescents who were higher in levels of conscientiousness reported increasing levels of support from their fathers over this age range. There were no significant relationships between perceived support from parents at age 12 and personality traits at age 17. The authors concluded that “core” traits, like conscientiousness, are relatively stable and enduring traits from birth to adulthood, while “surface” traits are more prone to influence from the environment. Their findings also supported cross-sectional research that hinted at the greater influence of personality on parent relationship as age of the adolescent increases.
Sources of Influences on the Personality-Parenting Relationship: Findings from Behavior Genetics
Using behavior genetic methods, researchers have attempted to explicate the nature of the relationship between personality and parental relationship quality. For instance, some have hypothesized that the association between personality and family environment is genetically mediated. At this point, it is well known that measures of the family environment, including parenting, are modestly heritable (Bouchard & McGue, 1990; Elkins, McGue, & Iacono, 1997; Hur & Bouchard, 1995; McGue, Elkins, Walden, & Iacono, 2005; Plomin & Bergeman, 1991; Plomin, McClearn, Pedersen, Nesselroade, & Bergeman, 1988; Reiss, Neiderhiser, Heatherington, & Plomin, 2000; Rowe, 1981, 1983). It was proposed that these measures are heritable because they are influenced by personality traits, which are, on average, 50% heritable (Bouchared & Loehlin, 2001). That is, a person's individual characteristics affect how they interact with others (or, alternately, how they think others interact with them), so the genetic influences on “environmental” measures actually reflect the personality traits of the person.
The two mechanisms through which genetically influenced characteristics could affect measures of the environment have been labeled evocative and active gene-environment correlations (Scarr & McCartney, 1983). Evocative correlation occurs when differences in people's genetically-influenced characteristics evoke specific responses from the people around them. Adolescents who are highly emotionally labile, for example, will likely elicit very different reactions from their parents than adolescents with calm, even-tempered dispositions. Active correlation occurs when genetic influences on personality affect the process by which individuals select their environments (e.g., amount of time spent with parents) or the process by which they make attributions regarding aspects of their relationships with others. Adolescents prone toward excitement and sensation seeking may be more likely to choose to spend time with like-minded peers rather than with parents who try to reinforce more constrained behavior.
Multivariate genetic models with structural equation modeling detect gene-environment correlation by estimating the degree to which genetic influences on one variable are related to the genetic influences on a second variable. As the degree of genetic overlap between personality and parenting increases, it becomes more probable that genetic influences on personality are responsible for genetic influences on parenting. To date, research has found genetic relationships between personality traits and life events (Billig, Hershberger, Iacono, & McGue, 1996; Saudino, Pedersen, Lichtenstein, McClearn, & Plomin, 1997) and risk of divorce (Jocklin, McGue, & Lykken, 1996). Chipuer, Plomin, Pedersen, McClearn, and Nesselroade (1993) collected measures of current family environment, neuroticism, and extraversion in a sample of older adult twins. They found that the association between the personality traits and the Relationship Scale from the Moos Family Environment Scale was primarily genetically mediated; however, a significant portion of the genetic variation in family environment was unaccounted for by personality. Krueger, Markon, and Bouchard (2003) investigated whether the genetic influences on child-rearing environment, including parenting, could account for genetic influences on adult personality. Utilizing data from the Minnesota Study of Twins Reared Apart (MISTRA), Krueger et al. found that the connections between the personality traits of Negative Emotionality and Constraint and the amount of cohesion in the recalled family environment were genetically mediated. They concluded that the same genotype which led to adult personality traits also influenced recall of childhood rearing environment. Taken together, the results from these myriad studies would suggest that a person's genetically influenced personality traits will often influence the nature of their relationships within the family.
Current Study
The limitation in the behavior genetic work on personality and parenting relationship is that these bivariate quantitative genetic models average over any group differences within the population. This is similar to any main effects model in statistics; for instance, a regression equation predicting adolescent smoking behavior from access to cigarettes will average across the sample. However, much like an examination of moderation in that example may reveal a smaller effect when there is greater parental monitoring, it is also possible that the estimation of genetic and environmental influences on one phenotype may vary as a function of differences in another. This is a form of gene-environment (G×E) interaction, the genetic susceptibility to environmental risk, or, alternatively, differential genetic expression in different environments. When G×E occurs, the genetic influences on a phenotype are more or less important depending on the level of a second trait. Examples of this work and their findings include less genetic influence on depression in unmarried women (Heath, Eaves, & Martin, 1998), smaller genetic influence on disinhibition in more religions families (Boomsma, de Geus, van Baal, & Koopmans, 1999), and greater genetic influence on IQ in high-SES families (Turkheimer, Haley, Waldron, D'Onofrio, & Gottesman, 2003).
Utilizing models that allow for and quantify moderation of this kind to address the current research topic may increase our understanding of how the parent-adolescent relationship develops in the context of the adolescent's emerging personality. Research suggests that there is a bidirectional relationship between the development of personality and the emerging parent-child relationship. In prior work using the same sample of adolescent twins, we found that perceptions of the parenting relationship moderated the genetic and environmental influences on adolescent personality traits (Krueger, South, Johnson, & Iacono, 2007). We build on that study by examining the other direction of influence—how qualities of the parent-adolescent relationship, as reported by both the parent and adolescent, result from the adolescent's personality traits. Using new modeling for biometrical moderation, we examine whether the individual characteristics of the adolescent, that is, personality traits, can change or influence the etiology of the personality-parenting relationship.
Method
Participants
The current study utilized participants from the Minnesota Twin Family Study (MTFS), an ongoing population-based, longitudinal study of adolescent twins and their families. Birth records and public databases were used to locate more than 90% of twin births in the state of Minnesota from 1971 through 1985. Families were excluded from the study if either twin had a cognitive or physical handicap that would preclude them from completing our daylong, in-person assessment, or if the family lived more than one day's drive from our Minneapolis laboratory. Of the eligible families, 83% agreed to participate. Although there were no significant differences between participating and nonparticipating families in regard to socioeconomic status and self-reported mental health problems, parents in participating families had slightly, albeit significantly, more education (0.25 years) than parents in nonparticipating families (Iacono, Carlson, Taylor, Elkins, & McGue, 1999). Reflecting the population of Minnesota at the time of the twins’ birth, approximately 98% of the sample was Caucasian. Children gave informed assent, while parents gave informed consent for themselves and their children. Research protocol was approved by the University of Minnesota Institutional Review Board. Further information regarding all aspects of MTFS recruitment is detailed elsewhere (Iacono et al., 1999).
The MTFS utilizes two cohorts of twins in an accelerated longitudinal design. Participants enter the study at age 11 or 17 years (corresponding to younger and older cohorts, respectively) and return for follow-up assessments approximately every three years thereafter. The original 11-year-old cohort consisted of 756 same-sex, reared-together monozygotic (MZ) and dizygotic (DZ) twin pairs: 376 male (254 MZ; 122 DZ) and 380 female pairs (233 MZ; 147 DZ). The 17-year-old cohort consisted of 626 same-sex twin pairs: 289 male (188 MZ; 101 DZ) and 337 female (223 MZ; 114 DZ) pairs. For the purposes of the current study, we utilized data from both cohorts at the overlapping assessment point of age 17 years: the older cohort at intake and the younger cohort at their second follow-up visit. This included all 1,252 individuals from the older cohort at the intake assessment, and 1,320 twins from the younger cohort who completed the second follow-up assessment (87% of the younger cohort).
Participants were excluded from this total sample of 2,572 if they were missing data on all of the personality variables and all of the parenting variables (N=67) and if co-twin data was entirely missing (N=53). This brought the final sample size to 2,452, including 585 male twin pairs (386 MZ; 199 DZ) and 641 female twin pairs (412 MZ; 229 DZ). The greater percentage of MZ twins relative to DZ twins in this sample is due to an overrepresentation of MZ twins in the population from which the sample was drawn, as well as a somewhat higher participation rate of families with MZ twins (Hur, McGue, & Iacono, 1995). At the time participants completed the measures used in the current study they ranged in age from 16.55 to 20.12 years, with a mean of 17.79 (SD=.65).
Zygosity
In the MTFS, three estimates are used to determine twin zygosity. MTFS staff evaluates the twins’ physical similarity, including visage, hair color, and face and ear shape. Next, parents complete a standard zygosity questionnaire. Finally, ponderal and cephalic indices and fingerprint ridge count were measured. A previous validation study (N=50) demonstrated 100% accuracy of zygosity determination when these three estimates agree. When these three estimates do not agree, a blood sample is requested and a serological analysis is performed.
Assessment of Personality
Personality was measured with the Multidimensional Personality Questionnaire (MPQ; Tellegen & Waller, in press), a 198-item self-report personality measure designed to assess a broad range of personality characteristics across normal populations. All participants were mailed the MPQ prior to the assessment and asked to bring the completed inventory with them to their in-person visit. If the MPQ was not completed upon their arrival for their laboratory assessment or by the end of the day-long visit, participants were asked to take it home and return the completed measure to the study by mail. Internal consistency reliabilities for the MPQ range from .76 to .89, and 30-day test-retest reliabilities range from .82 to .92. The MPQ assesses eleven primary personality domains, 10 of which load on three higher-order factors (the 11th scale, Absorption, does not load on any factor). The three higher-order factors are Positive Emotionality (PEM; a broad measure of positive well-being and tendency to view life as a pleasurable experience), Negative Emotionality (NEM; a propensity to experience psychological distress) and Constraint (CN; a tendency to endorse traditional values and act in a cautious manner). Positive Emotionality subsumes the lower order scales of Well Being, Achievement, Social Potency, and Social Closeness. Negative Emotionality is comprised of Aggression, Alienation, and Stress Reaction. Finally, Constraint is a composite of Traditionalism, Control, and Harm Avoidance. Only the higher-order scales were included in the present analyses. MPQ data were available for 2,169 individuals (men=1,053, women=1,116) from the total sample of 2,452.
Parent-Adolescent Relationship Quality
The Parental Environment Questionnaire (PEQ) was administered to both the adolescent twins and their mothers to tap perceptions of parent-child relationships. The PEQ was mailed to all participants prior to the assessment. If the PEQ was not completed upon their arrival for their laboratory assessment or by the end of the day-long visit, participants were asked to complete it at home and return it by mail. The PEQ asks mothers and twins to complete 50 items assessing aspects of their relationship on a 4-point scale (1 = definitely true, 2 = probably true, 3 = probably false, and 4 = definitely false). Twins rated their relationships with their mothers and fathers, while mothers rated their own relationship with each individual twin. Items in the versions of the PEQ completed by mothers and twins were essentially the same, with minor changes in wording appropriate for the particular rater.
The PEQ was developed by the MTFS because the standard measures of family environment available when the study began failed to assess dyadic relationships within the family, instead focusing on the overall family climate. The PEQ scales were organized around the two broad domains of conflict (vs. nurturance/warmth) and control, and correlate significantly in the expected direction with an alternative measure of the family environment (Elkins et al., 1997). Further details regarding the development, theoretical rationale, and psychometric properties of the PEQ are given in Elkins et al. (1997). Five factor-analytically derived scores are produced by responses to the PEQ: Conflict, Parent Involvement, Regard for Parent, Regard for Child, and Structure. The Structure scale is the only scale to assess the control aspect; in addition, it has low internal reliability (Elkins et al., 1997), and thus we did not consider it in this study. For the present investigation, we utilized scores for the Conflict Scale (12 items: e.g., my parent often loses her/his temper with me; alpha = .82), Involvement Scale (12 items: e.g., I talk about my concerns and my experiences with my parent, alpha=.74), Regard for Parent Scale (8 items: e.g., I want to be like my parent in a number of ways; alpha = .75), and the Parent Regard for Twin Scale (5 items: e.g., my parent is proud of me; alpha = .69; McGue et al., 2005). Scores were prorated for scales missing ratings for 10% or fewer of their items (i.e., the average of the other items was used as the missing item's score); if more than 10% of the scale's constituent items were missing the scale was considered missing.
To create composite indices of Regard, Conflict, and Involvement we averaged ratings from mothers and twins (Burt, Krueger, McGue, & Iacono, 2003; Burt, McGue, Iacono, & Krueger, 2006; Burt, McGue, Krueger, & Iacono, 2005). Because twin ratings about mom and dad were highly correlated (> .80; McGue et al., 2005), we first averaged these ratings. Significant correlations were also found for 1) twin reports of Regard for Parent and Parent Regard for twin (r = .65, p<.0001), and 2) mother reports of Regard for Parent and Parent Regard for twin (r =.41, p<.0001) so scores for these two scales were combined to form summary scores for Regard. Separate composite indices were then created for Regard, Conflict, and Involvement. We averaged the twin composite and the mother report of twin to more fully capture the relational aspect of the parent-adolescent relationship (the correlations between mother report of twin and twin report of mother were .41 for Conflict, .29 for Regard, and .36 for Involvement). This is supported by previous work in the MTFS samples which has found that each informant uniquely contributes to the prediction of external validity criteria, including behavior problems and grades (Burt et al., 2005). PEQ data were available for 2,364 participants for Regard (men=1,119, women=1,245), 2,365 individuals for Conflict (men=1,120, women=1,245), and 2,364 individuals for Involvement (men=1,119, women=1,245).
Biometric Analyses
Biometric modeling was utilized to evaluate the genetic and environmental moderation of parent-adolescent relationship quality by adolescent personality. This type of modeling makes use of twin methodology and structural equation modeling to estimate how much of the variance in a trait (phenotype) is due to additive genetic effects, or the effect of individual genes summed over loci (A), shared environmental effects, or the extent to which growing up in the same family makes people similar (C), and nonshared environmental effects, or the extent to which people are unique, despite growing up in the same family (E). The standard univariate “ACE” model assumes that the A, C, and E components are fixed over the entire population from which the sample is drawn. In other words, there is no provision for the association between the genetic and environmental influences on personality and any other trait.
To test our assertion that the genetic and environmental influences on the parent-adolescent relationship differ as a function of adolescent personality, we needed to utilize a model that allowed the variance components of parent relationship quality to vary as a function of adolescent personality. This type of analysis has been referred to as a test of “gene-environment interaction,” or the notion that different environments can lead to different genetic expression of a phenotype. However, this term does not completely describe the nature of the effects we examined. In the current study, neither the moderator variable, personality, nor the dependent variable, parenting behavior, are wholly “environmental” or wholly “genetic.” The advantage of the model that we utilize is that it is possible to decompose the moderator variable into its genetic and environmental variance components, and test for gene-environment interaction in the presence of gene-environment correlation (Purcell, 2002). A genetic correlation is the amount of overlap in the genetic influences on two phenotypes and ranges from −1 to +1; similar types of correlations (i.e., overlap) can occur for shared and nonshared environmental influences. Therefore, we will use the more accurate term “biometrical moderation” to refer to the analyses conducted in these studies; this term better captures the goal of this study—to determine whether the magnitude of genetic and environmental influences on parenting depends on the adolescent's level of personality (gene-environment interaction), and the extent to which influences acting on parenting also exerted influences on personality (gene-environment correlation).
Biometric models were fit to the raw data using Mx software system (Neale, Boker, Xie, & Maes, 2003). To ease interpretation, the parenting variables were recoded as necessary so that they would be positively correlated with the personality variables (which remained scored in the same direction for all models, with higher scores corresponding to greater PEM, NEM or CN). To correct for potential biases in model fitting, the personality and parenting relationship scales were adjusted for effects of age and gender (McGue & Bouchard, 1984). Each scale was regressed on age, age2, age × gender, and age2 × gender, and the standardized residuals from these regressions were used in subsequent analyses. Because not all participants had both MPQ and PEQ data for all scales, we used full-information maximum-likelihood with the raw data, a procedure that was also necessary for the moderated biometric models we were using. This procedure relies on the assumption that data are missing at random (Little & Rubin, 2002), an assumption that we considered reasonable in this case, as questionnaire return status was not linked in any way to any study measure status. Fit of the moderation models were judged relative to the fit of a bivariate decomposition model in which the six moderation parameters (βXcM and βXuM for A, C, and E) were fixed at zero (so that aC+βXcM became aC + (0*M) = aC).
Two indices were used to evaluate model fit: (1) the likelihood-ratio test (LRT; distributed as χ2, and computed as the difference in the −2 log-likelihood values for the two models); and (2) the Akaike Information Criterion (AIC; Akaike, 1987). The LRT is used as a goodness-of-fit index, representing the degree of fit between model expectations and observed data. Statistically significant values are associated with a relatively poor fit. Improvements in the model's fit, from adding or omitting parameters, can be assessed by a statistically significant change in LRT. Akaike's Information Criterion (AIC; Akaike, 1987) is also conventionally used to compare the fit of alternative models. The AIC considers goodness of fit in the likelihood sense (how well the model reproduces the observed data), but prefers models that capture the data both accurately and parsimoniously over more complex models. Because the aim of model fitting is to explain the data as parsimoniously as possible, the model with the lowest AIC value is generally considered best.
Results
Descriptive Statistics
The phenotypic correlations between personality and parenting variables were calculated using Mplus (Muthén & Muthén, 1998-2006), which uses an MLR estimator to produce confidence intervals that are adjusted for the non-independence of the twin data. The relationships among the transformed variables were in the expected directions (see Table 1). Regard was positively correlated with PEM (r = 0.25, 95% CI = (0.20, 0.31), p<.0001), negatively correlated with NEM (r = −0.25, 95% CI = (−0.29, −0.21), p<.0001), and positively correlated with CN (r = 0.24, 95% CI = (0.19, 0.29), p<.0001). Conflict was negative correlated with PEM (r =−0.11, 95% CI = (−0.16, −0.06), p<.0001), positively correlated with NEM (r = 0.35, 95% CI = (0.30, 0.39), p<.0001), and negatively correlated with CN (r = −0.26, 95% CI = −0.31, −0.21, p<.0001). Involvement was positively correlated with PEM (r = 0.29, 95% CI = (0.24, 0.34), p<.0001), negatively correlated with NEM (r = −0.25, 95% CI = (−0.30, −0.21), p<.0001), and positively correlated with CN (r = 0.26, 95% CI = (0.21, 0.31), p<.0001).
Table 1.
Phenotypic and Twin Correlations for Parenting and Personality Measures
| Twin Correlations | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | MZ |
DZ |
||
| 1 | Regard | - | 0.70 | 0.52 | |||||
| 2 | Conflict | −0.63 | - | 0.72 | 0.46 | ||||
| 3 | Involvement | 0.77 | −0.62 | - | 0.71 | 0.53 | |||
| 4 | Positive Emotionality | 0.25 | −0.11 | 0.29 | - | 0.52 | 0.22 | ||
| 5 | Negative Emotionality | −0.25 | 0.35 | −0.25 | −0.10 | - | 0.49 | 0.17 | |
| 6 | Constraint | 0.24 | −0.26 | 0.26 | 0.17 | −0.08 | - | 0.54 | 0.21 |
Also shown in Table 1 are basic MZ and DZ twin correlations for the personality and parenting measures. Twin correlations were computed using an intra-class correlation coefficient in SPSS. These MZ and DZ intra-class correlations can be compared to obtain a general indication of the extent to which genetic and environmental influences are operating on the phenotype. In all cases, the MZ correlations exceeded the DZ correlations, suggesting that both personality and parenting measures are likely to be influenced by genetic effects. However, MZ correlations for the parenting measures were less than double the DZ correlations, implicating the importance of shared environmental effects.1
Biometric Moderation Analysis
We tested whether adolescent personality traits moderated the genetic and environmental influences on the quality of the parent-adolescent relationship by fitting our variables to the biometrical moderation model in Figure 1. There were nine combinations of parenting and personality variables: Regard moderated by PEM, NEM, and CN; Conflict moderated by PEM, NEM and CN; and Involvement moderated by PEM, NEM, and CN. For each of these nine combinations, we compared the fit of a model with all moderation parameters fixed to 0 (No Moderation) to a model with all moderation parameters estimated (A, C, and E Full Moderation). As shown in Table 2, full ACE biometrical moderation was significant for seven of the nine possible combinations of parenting and personality variables. These results were supported by both LRT and AIC. The strongest effects were for the moderating effects of PEM (χ2=54.89, df=6, p<.0001), NEM (χ2=56.53, df=6, p<.0001) and CN (χ2=59.87, df=6, p<.0001) on Regard. The moderating effects of PEM on Involvement and CN on Conflict could be removed without a significant decrease in fit, suggesting that the genetic and environmental effects on Involvement and Conflict were the same across all levels of PEM and CN, respectively.
Figure 1.

Path diagram of a biometrical moderation model with adolescent personality (PERSONALITY) moderating the genetic and environmental influences on parent-adolescent relationship (PARENTING). The model is a variation of the bivariate (Cholesky) decomposition model, in which the variances and covariances of the observed variables are decomposed into the proportion of variance associated with genetic (a2), shared environmental (c2) and nonshared environmental (e2) components that are shared between the phenotypes and unique to one of. There are two sets of paths contributing genetic and environmental influences: those common to parent-adolescent relationship and the moderator (personality), and those unique to parent relationship. The paths from the moderator (M) variable to the dependent variable are now linear functions of the form a + βM, where a is the parameter for genetic influence on the variable, β is a regression coefficient, and M is the level of the moderator variable.
Table 2.
Fit Statistics From the Models of Variance Components Allowing for Gene-Environment Interaction and Correlation
| Regard | Moderating Variable |
|||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Positive Emotionality |
Negative Emotionality |
Constraint |
||||||||||||||||
| −2lnL |
df |
ΔX2 |
Δdf |
p |
AIC |
−2lnL |
df |
ΔX2 |
Δdf |
p |
AIC |
−2lnL |
df |
ΔX2 |
Δdf |
p |
AIC |
|
| No Moderation | 10566.98 | 4038 | 2490.98 | 10558.10 | 4038 | 2482.10 | 10577.61 | 4038 | 2501.61 | |||||||||
| Only A Moderation | 10547.67 | 4036 | 19.31 | 2 | 0.000 | 2475.67 | 10509.91 | 4036 | 48.19 | 2 | 0.000 | 2437.91 | 10555.89 | 4036 | 21.72 | 2 | 0.000 | 2483.89 |
| Only C Moderation | 10537.09 | 4036 | 29.89 | 2 | 0.000 | 2465.09 | 10516.23 | 4036 | 41.87 | 2 | 0.000 | 2444.23 | 10553.98 | 4036 | 23.63 | 2 | 0.000 | 2481.98 |
| Only E Moderation | 10519.83 | 4036 | 47.15 | 2 | 0.000 | 2447.83 | 10529.69 | 4036 | 28.41 | 2 | 0.000 | 2457.69 | 10531.40 | 4036 | 46.21 | 2 | 0.000 | 2459.40 |
| A and C Moderation (no E) | 10535.49 | 4034 | 31.49 | 4 | 0.000 | 2467.49 | 10507.58 | 4034 | 50.52 | 4 | 0.000 | 2439.58 | 10552.29 | 4034 | 25.32 | 4 | 0.000 | 2484.29 |
| A and E Moderation (no C) | 10517.11 | 4034 | 49.87 | 4 | 0.000 | 2449.11 | 10504.38 | 4034 | 53.71 | 4 | 0.000 | 2436.38 | 10524.95 | 4034 | 52.66 | 4 | 0.000 | 2456.95 |
| C and E Moderation (no A) | 10514.19 | 4034 | 52.79 | 4 | 0.000 | 2446.19 | 10502.93 | 4034 | 55.16 | 4 | 0.000 | 2434.93 | 10520.14 | 4034 | 57.47 | 4 | 0.000 | 2452.14 |
| A, C, and E Full Moderation | 10512.09 | 4032 | 54.89 | 6 | 0.000 | 2448.09 | 10501.57 | 4032 | 56.53 | 6 | 0.000 | 2437.57 | 10517.74 | 4032 | 59.87 | 6 | 0.000 | 2453.74 |
| Conflict | ||||||||||||||||||
| No Moderation | 10597.12 | 4039 | 2519.12 | 10385.29 | 4039 | 2307.29 | 10491.52 | 4039 | 2413.52 | |||||||||
| Only A Moderation | 10596.10 | 4037 | 1.02 | 2 | 0.600 | 2522.10 | - | - | - | - | - | - | - | - | - | - | - | - |
| Only C Moderation | 10588.46 | 4037 | 8.67 | 2 | 0.013 | 2514.46 | - | - | - | - | - | - | - | - | - | - | - | - |
| Only E Moderation | 10595.86 | 4037 | 1.26 | 2 | 0.532 | 2521.86 | - | - | - | - | - | - | - | - | - | - | - | - |
| A and C Moderation (no E) | 10587.26 | 4035 | 9.86 | 4 | 0.043 | 2517.26 | - | - | - | - | - | - | - | - | - | - | - | - |
| A and E Moderation (no C) | 10584.03 | 4035 | 13.09 | 4 | 0.011 | 2514.03 | - | - | - | - | - | - | - | - | - | - | - | - |
| C and E Moderation (no A) | 10583.78 | 4035 | 13.35 | 4 | 0.010 | 2513.78 | - | - | - | - | - | - | - | - | - | - | - | - |
| A, C, and E Full Moderation | 10581.12 | 4033 | 16.00 | 6 | 0.014 | 2515.12 | 10365.75 | 4033 | 19.53 | 6 | 0.003 | 2299.75 | 10480.89 | 4033 | 10.63 | 6 | 0.101 | 2414.89 |
| Involvement | ||||||||||||||||||
| No Moderation | 10497.52 | 4039 | 2419.52 | 10505.15 | 4039 | 2427.15 | 10518.30 | 4039 | 2440.30 | |||||||||
| Only A Moderation | - | - | - | - | - | - | 10471.38 | 4037 | 33.77 | 2 | 0.000 | 2397.38 | - | - | - | - | - | - |
| Only C Moderation | - | - | - | - | - | - | 10476.68 | 4037 | 28.47 | 2 | 0.000 | 2402.68 | - | - | - | - | - | - |
| Only E Moderation | - | - | - | - | - | - | 10486.91 | 4037 | 18.23 | 2 | 0.000 | 2412.91 | - | - | - | - | - | - |
| A and C Moderation (no E) | - | - | - | - | - | - | 10470.66 | 4035 | 34.49 | 4 | 0.000 | 2400.66 | - | - | - | - | - | - |
| A and E Moderation (no C) | - | - | - | - | - | - | 10468.75 | 4035 | 36.40 | 4 | 0.000 | 2398.75 | - | - | - | - | - | - |
| C and E Moderation (no A) | - | - | - | - | - | - | 10471.07 | 4035 | 34.08 | 4 | 0.000 | 2401.07 | - | - | - | - | - | - |
| A, C, and E Full Moderation | 10486.68 | 4033 | 10.84 | 6 | 0.094 | 2420.68 | 10467.82 | 4033 | 37.33 | 6 | 0.000 | 2401.82 | 10502.59 | 4033 | 15.70 | 6 | 0.015 | 2436.59 |
Note. −2lnL = −2 log likelihood; df = degrees of freedom; AIC = Akaike's Information Criterion. Best-fitting moderation models are shown in bold.
Confirmation of findings with transformed data
We sought to confirm that these results were not simply the result of non-normality in the data using two approaches. First, we again compared the No Moderation and ACE Full Moderation models, this time using transformed scores for Conflict (skew = 0.436), Regard (skew = −1.117), and Involvement (skew = −.571). In order to use the same type of transformation on all variables, Conflict was first reverse scored and all variables were subjected to a square transformation. Results reported above for the raw data (age and gender regressed) were generally replicated using transformed scores for Regard, Conflict, and Involvement; however, the effects of NEM on Conflict (χ2=7.77, Δdf=6, p=ns) and CN on Involvement (χ2=8.88, Δdf=6, p=ns) were no longer significant.
In a second, separate attempt to confirm that our findings were not the result of extreme data points, we truncated the raw data for the three personality variables—PEM, NEM, and CN. After age and gender were regressed out of the personality variables, the z-score residuals were trimmed such that anyone above +2 or below −2 was recoded to these boundaries. When these truncated personality variables were used, all seven significant effects found with raw data were replicated. 2
Further analysis of moderation models
We now turn to further interpretation of the moderation models, but limit our discussion to the five models that were best supported with analyses using raw and transformed data: Regard moderated by PEM, NEM, and CN, Conflict moderated by PEM, and Involvement moderated by NEM. Since the moderation of Conflict by NEM and Involvement by CN could not be completely confirmed using transformed data, we remain cautious in our interpretation of these results and do not discuss these models further.
Having established that the full moderation model with all moderation parameters freely estimated provided a better fit to the data than the no moderation models for five of the combinations of parenting and personality variables, we then sought to establish which moderation parameters were driving the effect. That is, we wished to determine whether all of the three types of variance (genetic, shared environmental, unique environmental) were moderated by the personality variables, or whether moderation occurred for some of these variance components and not others. Starting from the no moderation, baseline model, we added moderation for each of the A, C, and E paths and their combinations in turn. In sum, a total of six models (in addition to the no moderation and full moderation models) were run: 1) only A moderation (no C and E); 2) only C moderation (no A and E); 3) only E moderation (no A and C); 4) A and C moderation (no E); 5) A and E moderation (no C); and 6) C and E moderation (no A). The results for this full series of models are discussed in turn below. The results of the full series of models are presented in Table 2, with the best fitting moderation models highlighted in bold.
Table 3 presents the estimated variance components (A, C, and E) for Regard, Conflict, and Involvement from the no-moderation models and the best-fitting moderation models. Estimates from the no-moderation models are equivalent to estimates from a standard bivariate decomposition model, such that they apply to the population in the aggregate, regardless of level of personality. When moderation is significant for any of the ACE components, then the variance estimates vary as functions of every level of the moderator variable (here, personality); for ease of presentation, they are shown in Table 3 at five different levels, scaled in standard deviation units (z-scored): −2, −1, 0, 1, and 2 standard deviations away from the mean of the moderator.3 Two types of ACE estimates are given in Table 3, for both the moderation and no-moderation models: (1) the raw A, C, and E estimates are shown in the first three columns, followed by the total phenotypic variance; and (2) A, C, and E estimates expressed as proportions of the total variance in parenting (e.g., A% = A / A+C+E, also referred to as h2, the heritability estimate). The final three columns present the genetic and environmental correlations (rA, rC, and rE).
Table 3.
Estimates of Unstandardized and Standardized Variance Components and Genetic and Environmental Correlations in Parent-Adolescent Relationship Quality and the Personality Moderating Variables
| Moderating Variable |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Variance components | Total Variance | Proportions of variance | Correlations with moderator | |||||||
| A |
C |
E |
|
A(%) |
C(%) |
E(%) |
rA |
rC |
rE |
|
| Positive Emotionality (PEM) |
||||||||||
| Regard | ||||||||||
| No Moderation Model Regard at Level of PEM | 0.40 | 0.32 | 0.29 | 1.00 | 0.40 | 0.31 | 0.28 | 0.23 | 1.00 | 0.18 |
| −2 | 0.22 | 0.69 | 0.43 | 1.34 | 0.16 | 0.52 | 0.32 | 0.17 | 1.00 | 0.36 |
| −1 | 0.31 | 0.46 | 0.33 | 1.11 | 0.28 | 0.42 | 0.30 | 0.23 | 1.00 | 0.28 |
| 0 | 0.42 | 0.28 | 0.26 | 0.96 | 0.44 | 0.30 | 0.27 | 0.27 | 1.00 | 0.18 |
| 1 | 0.55 | 0.15 | 0.20 | 0.89 | 0.62 | 0.16 | 0.22 | 0.31 | 1.00 | 0.04 |
| 2 | 0.69 | 0.05 | 0.15 | 0.90 | 0.77 | 0.06 | 0.17 | 0.33 | 1.00 | −0.14 |
| Conflict | ||||||||||
| No Moderation Model Conflict at Level of PEM | 0.56 | 0.17 | 0.26 | 0.99 | 0.56 | 0.18 | 0.26 | −0.06 | 1.00 | 0.15 |
| −2 | 0.74 | 0.01 | 0.28 | 1.04 | 0.72 | 0.01 | 0.27 | −0.18 | 1.00 | 0.39 |
| −1 | 0.64 | 0.07 | 0.26 | 0.96 | 0.66 | 0.07 | 0.27 | −0.12 | 1.00 | 0.28 |
| 0 | 0.54 | 0.17 | 0.24 | 0.95 | 0.57 | 0.18 | 0.25 | −0.05 | 1.00 | 0.15 |
| 1 | 0.46 | 0.33 | 0.23 | 1.02 | 0.45 | 0.32 | 0.23 | 0.03 | 1.00 | 0.01 |
| 2 | 0.39 |
0.53 |
0.23 |
1.16 |
0.34 |
0.46 |
0.20 |
0.13 |
1.00 |
−0.12 |
| Negative Emotionality (NEM) |
||||||||||
| Regard | ||||||||||
| No Moderation Model Regard at Level of NEM | 0.40 | 0.31 | 0.29 | 1.00 | 0.40 | 0.31 | 0.28 | 0.28 | 1.00 | 0.18 |
| −2 | 0.36 | 0.09 | 0.20 | 0.65 | 0.55 | 0.13 | 0.32 | 0.35 | 1.00 | 0.36 |
| −1 | 0.36 | 0.20 | 0.24 | 0.79 | 0.45 | 0.25 | 0.30 | 0.35 | 1.00 | 0.27 |
| 0 | 0.36 | 0.36 | 0.28 | 0.99 | 0.36 | 0.36 | 0.28 | 0.35 | 1.00 | 0.18 |
| 1 | 0.36 | 0.56 | 0.32 | 1.24 | 0.29 | 0.45 | 0.26 | 0.35 | 1.00 | 0.10 |
| 2 | 0.36 | 0.81 | 0.38 | 1.54 | 0.23 | 0.52 | 0.25 | 0.35 | 1.00 | 0.03 |
| Involvement | ||||||||||
| No Moderation Model Involvement at Level of | 0.44 | 0.29 | 0.27 | 1.00 | 0.45 | 0.29 | 0.27 | 0.31 | 1.00 | 0.20 |
| NEM | ||||||||||
| −2 | 0.24 | 0.28 | 0.26 | 0.78 | 0.31 | 0.36 | 0.33 | 0.63 | 1.00 | 0.20 |
| −1 | 0.32 | 0.28 | 0.26 | 0.86 | 0.38 | 0.33 | 0.30 | 0.46 | 1.00 | 0.20 |
| 0 | 0.44 | 0.28 | 0.26 | 0.98 | 0.45 | 0.29 | 0.26 | 0.32 | 1.00 | 0.20 |
| 1 | 0.60 | 0.28 | 0.26 | 1.13 | 0.53 | 0.25 | 0.23 | 0.22 | 1.00 | 0.20 |
| 2 | 0.79 |
0.28 |
0.26 |
1.32 |
0.59 |
0.21 |
0.19 |
0.14 |
1.00 |
0.20 |
| Constraint (CN) |
||||||||||
| Regard | ||||||||||
| No Moderation Model Regard at Level of CN | 0.40 | 0.32 | 0.29 | 1.01 | 0.40 | 0.32 | 0.28 | 0.40 | 1.00 | 0.06 |
| −2 | 0.44 | 0.54 | 0.46 | 1.44 | 0.30 | 0.37 | 0.32 | 0.32 | 1.00 | −0.02 |
| −1 | 0.44 | 0.40 | 0.36 | 1.20 | 0.37 | 0.33 | 0.30 | 0.32 | 1.00 | 0.02 |
| 0 | 0.44 | 0.28 | 0.27 | 0.99 | 0.44 | 0.28 | 0.27 | 0.32 | 1.00 | 0.08 |
| 1 | 0.44 | 0.18 | 0.20 | 0.82 | 0.54 | 0.22 | 0.24 | 0.32 | 1.00 | 0.15 |
| 2 | 0.44 | 0.10 | 0.14 | 0.68 | 0.65 | 0.15 | 0.20 | 0.32 | 1.00 | 0.26 |
Moderation of Regard by Positive Emotionality
The models based on PEM and Regard are displayed graphically in Figure 2. The first panel illustrates the unstandardized variance components for Regard from the no-moderation model decomposition of PEM and Regard. As shown, the A (genetic) component is .40, while the E (non-shared environmental) and C (shared environmental) components are about .30, corresponding to the variance estimates given in Table 3. As this model does not take into account moderation of parenting by personality, the variance components are static across the range of PEM. As shown in Table 2, the moderation model clearly fit the data better than the no moderation model. Further parsing of the full ACE moderation model revealed that two submodels of the moderation model, in which there was moderation on E only (χ2=47.15, Δdf=2, p<.0001, AIC=2447.83) and moderation on C and E only (χ2=52.79, Δdf=4, p<.0001, AIC=2446.19), technically fit better than the full ACE moderation model (χ2=54.89, Δdf=6, p<.0001, AIC=2448.09). However, examination of the plot of the full ACE model revealed a clearly visible effect of A and C, thus convincing us that the full moderation model best represented the true nature of the moderation of Regard by PEM. Figure 2 shows the results of estimates from the full ACE moderation model, in which all three ACE variance components for Regard vary as functions of Positive Emotionality (shown as z-scores from −2 to +2). As PEM increases, the genetic variance of Regard increases, while both the shared and non-shared environmental effects decrease.
Figure 2.

Panel 1: No-Moderation Model of Regard and Positive Emotionality. Panel 2: Moderation Model of Regard as a function of Positive Emotionality. A=genetic variance, C=shared environmental variance, E=nonshared environmental variance.
With no moderation, the proportion of variance in individual differences in Regard (from a bivariate decomposition model with PEM) is 40% genetic, 32% shared environmental, and 29% non-shared environmental. This is very similar to the results from the moderation model at a mean level (z-score of 0) of PEM (h2=.42, c2=.28, e2=.26). As Table 3 shows, though, the genetic component of variance increases from low to high levels of PEM; because the total phenotypic variance in Regard decreases from low to high levels of PEM, the heritability of Regard is greatest when PEM is greatest. At low levels of PEM, Regard is largely due to shared (c2=52%) and non-shared environmental effects (e2=32%), with a smaller contribution from genetic effects (h2=16%). At high levels of PEM, a majority of the variance in Regard is due to additive genetic effects (h2=77%), while non-shared environmental effects have decreased (e2=17%) and there are only very small shared environmental effects (c2=6%).
Examination of the genetic correlations between Regard and PEM reveal a somewhat stronger association between the genetic influences on PEM and the genetic influences on Regard at high levels of PEM (see rA in Table 3).3 The genetic correlation between PEM and Regard increased from rA =0.17 at low levels of PEM (−2) to rA=0.33 at high levels of PEM (+2). For adolescents who are higher in Positive Emotionality, there are more genetic influences common to both this constellation of personality traits and the amount of warmth in their relationship with both parents. Conversely, the overlap between non-shared environmental influences on PEM and Regard decreases as level of PEM increases (where the proportion of variance attributable to E is lowest). Therefore, the variance in Regard due to non-shared environmental influences is greater at low levels of PEM, and it is here than the non-shared environmental correlation between PEM and regard is greatest (rE = 0.36 at PEM of −2).
Moderation of Conflict by Positive Emotionality
Figure 3 shows the best-fitting moderation model of Positive Emotionality on Conflict. Again, while the full moderation model was not technically the best fitting moderation model according to AIC and p value (see Table 2), it was very close, and the plot of the full moderation model showed an effect of A that justified its choice as the best-fitting model. As PEM increased, the genetic variance component of Conflict decreased, the shared environmental variance increased, and the nonshared environmental variance decreased slightly (see Table 3). Thus, at low levels of PEM, the proportion of variance in Conflict was weighted heavily toward genetic (h2=72%) influences, with the rest split mainly attributable to non-shared environmental (e2=27%) effects. At high levels of PEM, proportion of variance due to genetic effects was much smaller (h2=34%), while shared environmental effects (c2=46%) had increased dramatically and unique environmental (e2=20%) effects had decreased slightly. The non-shared environmental correlation between PEM and Conflict decreased as levels of PEM increased, from rE = 0.39 at PEM of −2 to rE = −0.12 at PEM of +2.
Figure 3.
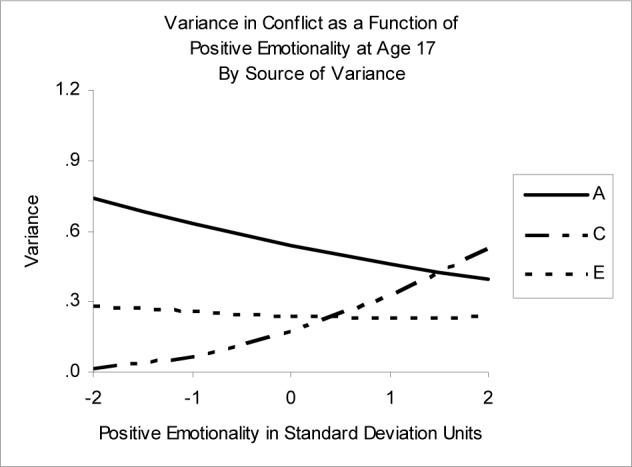
Variance in Conflict as a function of Positive Emotionality. A=genetic variance, C=shared environmental variance, E=nonshared environmental variance.
Moderation of Regard by Negative Emotionality
In the best-fitting moderation model of Regard and NEM, there was significant moderation of the C and E variance components (see Figure 4). Contrary to PEM, for which the influence of shared environment on Regard was largely negligible at higher levels of personality, shared environment was greater than genetic and non-shared environmental effects at high levels of NEM. The C and E variance components increased as functions of NEM; as a result, there was greater total variance in Regard at high levels of NEM. Thus, even though the genetic variance remained stable from low to high levels of NEM, the proportion of variance due to genetic effects declined. At low levels of NEM (−2), most of the variance in Regard was split between genetic (h2=55%) and non-shared environmental effects (e2=32%). However, at high levels of NEM (+2), the proportion of variance was weighted more heavily toward shared environment (h2=23%, c2=52%, e2=25%). The overlap between unique environmental influences on Regard and NEM were greatest at low levels of NEM (rE=.36 at NEM of −2 SD).
Figure 4.
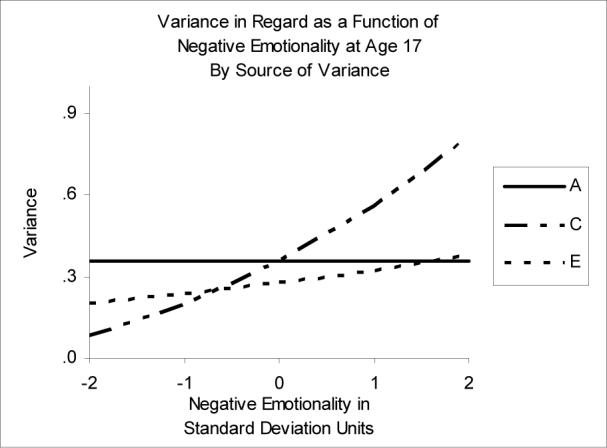
Variance in Regard as a function of Negative Emotionality. A=genetic variance, C=shared environmental variance, E=nonshared environmental variance.
It is interesting to note that, as with the results for PEM and Conflict, when the valence (i.e., positively vs. negatively tinged) of the relationship and the valence of the personality trait are mismatched, the etiology of the relationship variable is more attributable to shared environmental effects at higher levels of the personality trait. That is, C effects increase with increasing levels of personality when either: 1) the parenting variable measures a positive aspect of the relationship (Regard) and the personality variable is a relatively more negative trait (NEM), or 2) when the parent variable measures a negative aspect of the relationship (Conflict) and the personality trait is relatively more positive (PEM).
Moderation of Involvement by Negative Emotionality
In the best-fitting moderation model, only the A variance component of Involvement was significantly moderated by NEM (see Figure 5). As the level of Involvement increased, the genetic variance increased; since the C and E variance components were stable across all levels of NEM, this resulted in greater phenotypic variance and higher heritability at high levels of NEM. At low levels of NEM, the proportion of variance due to genetic and environmental effects were almost equally divided (h2=31%, c2=36%, e2=33%). The proportion of variance due to genetic effects almost doubled from low to high levels of NEM (h2=59% at SD=+2). However, the genetic correlation between Involvement and NEM decreased from low to high levels of NEM. Even though genetic effects had a greater influence on Involvement at high levels of NEM, the types of genetic effects operating on Involvement and NEM were different.
Figure 5.
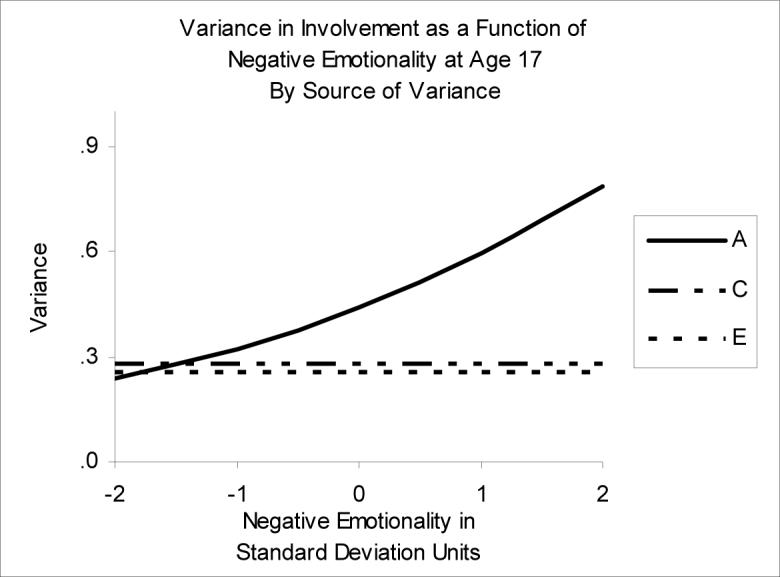
Variance in Involvement as a function of Negative Emotionality. A=genetic variance, C=shared environmental variance, E=nonshared environmental variance.
Moderation of Regard by Constraint
The results of moderation analysis for Regard and Constraint were similar to the moderation model of Regard and Positive Emotionality, although in the best-fitting model for Regard and CN moderation was significant for C and E only. The influences on Regard shifted from an almost equal distribution of the proportion of variance among all three components at low levels of CN to largely genetic influences at higher levels of CN (see Figure 6 and the values in Table 3, which show the ACE decomposition of Regard as a function of CN). Estimates derived from the moderation model show a decrease in both sources of environmental variance, from (c2=37%, e2=32%) at low CN to (c2=15%, e2=20%) at high CN.
Figure 6.
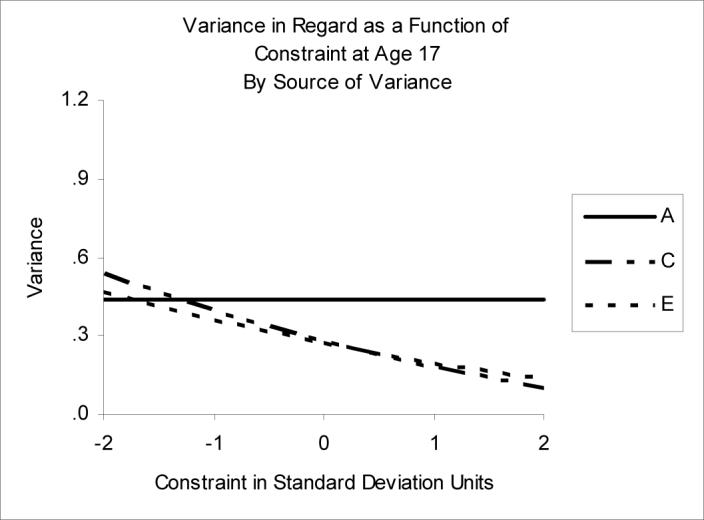
Variance in Regard as a function of Constraint. A=genetic variance, C=shared environmental variance, E=nonshared environmental variance.
Notably different from the moderation of Regard by PEM, however, was the finding of greater non-shared environmental influences between Regard and CN at higher levels of CN. The non-shared environmental correlation between CN and Regard increased with increasing levels of CN. For adolescents higher in Constraint, the presence of these non-shared environmental influences common to CN and Regard suggest a within family selection process linking the two.
Extension of Analyses to Both Sources of Report on Parent-Adolescent Relationship
Because we utilized a composite report of parent-adolescent relationship, we also conducted analyses separately for the mother's and adolescent's perspective of the relationship, to determine if the pattern of results differed depending on the reporter. In general, the results for the composite report are closer to the results found using adolescent report rather than mother report. Using the raw scores for adolescent report, six of the seven moderation models that were significant using the composite report were also significant (PEM and Regard, Conflict; NEM and Regard, Conflict, Involvement; CN and Regard), while for mother report of the relationship, five of the same models which were significant for composite report were also an improvement over the no-moderation models (PEM with Conflict; NEM with Regard, Conflict, Involvement; CN with Regard). However, when transformed scores were used, moderation models fit better for four of the same five models using adolescent-only report as when the composite measure was used (PEM with Regard, Conflict; NEM with Regard; CN with Regard), while moderation models using mother report were significant for only two of the same models as when the composite measure was used (NEM and Regard, Involvement). Given the moderately strong, but not perfect, nature of the correlation between mother and adolescent report of the relationship, we feel that the composite report most likely is the best way to capture the parent-adolescent relationship. Differences in moderation of the etiology of the parent-adolescent relationship depending on the reporter are difficult to interpret, especially since there is no theory to guide why results would be different (or not). Complete results for moderation analyses of the parent-adolescent relationship (as reported by the mother and the adolescent) can be obtained from the first author
Discussion
In the developmental context of the family, it appears that personality and interpersonal relationships are involved in a bidirectional process of reinforcement. The goal of the current study was to examine whether personality traits can moderate genetic and environmental influences on positive and negative qualities of the parent-adolescent relationship. Specifically, we utilized a sample of more than 2,400 individual twins at a mean age of 17 years to examine whether adolescent personality traits of Negative Emotionality, Positive Emotionality, and Constraint moderated the genetic and environmental contributions to three aspects of the parent-adolescent relationship—namely, Regard, Conflict, and Involvement. We found: (1) significant moderation of all three features of the parent relationship by these personality traits; and (2) changes in the genetic and environmental correlations between personality and parenting at different levels of personality. The present study's large sample size was a notable strength. Of course, it is possible that with an even larger sample size, we may have found significant moderation in the four models where moderation was not detected (i.e., PEM and Involvement, NEM and Conflict, CN and Conflict, CN and Involvement).
The goal of the current study was to extend previous work examining genetic and environmental influences on the relationship between personality and parent relationship quality. Our results support previous research which attributes genetic and environmental influences on perceptions of the family environment to personality traits (Krueger et al., 2003). Further, our findings suggest that whether a genotype will lead to positive or negative aspects of the parent relationship depends on the personality of the adolescent. Beginning with the groundbreaking work of David Rowe (1981, 1983), behavior genetic methods have been used to show that putatively “environment” measures have a sizeable genetic component. Prior work from the Minnesota Twin Family Study that examined the parenting measures used in the current study found small to moderate heritability estimates (Elkins et al., 1997) which tended to increase with age of the adolescent (McGue et al., 2005). In accord with these results, we found that aspects of the parent relationship were moderately heritable. In the no-moderation models, which again are comparable to standard bivariate decomposition models, 40% of the variance in Regard, 56% of the variance in Conflict, and 45% of the variance in Involvement was due to genetic effects, with the rest of the variance attributed generally equally to shared and non-shared environment. We also found moderate genetic correlations between aspects of the parent-adolescent relationship and adolescent personality traits, which is consistent with prior research findings that the heritability of family and parenting measures is due, at least in part, to the characteristics (i.e., personality traits) of the family members (Chipuer et al., 1993; Krueger et al., 2003).
The limitation in these prior studies (and in the results of our own un-moderated models) is that estimates of genetic and environmental influences from univariate models and estimates of genetic correlations from bivariate models of personality and family environment are static and fixed across the entire population. Newer models that test for biometrical moderation, or gene-environment interaction, examine whether genetic and environmental influences may differ in different environments. Conceptually, the idea is similar to examining differences in the main effect of a variable across different groups. For example, does the effect of a reading intervention (main effect) differ between men and women (groups)? Now extending this idea to the realm of biometrical modeling, we replace the main effect with genetic and environmental influences (here, on parenting behaviors) and different groups with different levels of a moderator variable (here, personality traits). Thus, we have the ability to calculate a specific heritability estimate for an individual dependent on where that person falls along the dimension of a second, moderating variable (Purcell, 2002).
A major contribution of the current study was the finding that, for aspects of warmth, conflict, and involvement in the parent-adolescent relationship, the relative contribution of genetic and environmental influences varied as a function of the adolescent's amount of negative emotionality, positive emotionality, or constraint. Specifically, we found that PEM, NEM, and CN significantly moderated the genetic and environmental influences on Parental Regard, PEM significantly moderated the etiology of Conflict, and NEM moderated the etiology of Involvement. Along with variations in the proportion of variance due to genes and environmental influences, we found different levels of genetic and environmental correlations between personality and parenting at varying levels of personality. These moderation models present a fuller picture of the relative influence of genes vs. environment on the etiology of the parent-adolescent relationship.
Considering only the static parameter estimates from a no moderation model, the etiology of Regard was weighted toward genetic influences, but with sizable shared and non-shared environmental influences. Results from moderation models tell a more nuanced story. The total phenotypic variance in Regard was generally lowest, and the heritability highest, at the most positive level of the personality trait; that is, at high levels of positive emotionality and constraint and low levels of negative emotionality. Adolescents with the greatest levels of positively valenced personality traits had a level of Regard in their relationship with their parents that better reflects their genotype. This is an example of genes operating differently in different environments—being high in positive emotionality or constraint (or low in negative emotionality) allows for the expression of a genetic predisposition to positive aspects of the parent-adolescent relationship.
Our findings provide further empirical support for the role of adolescent personality in the quality of the parent-adolescent relationship. The etiology of the parent-adolescent relationship appears to differ depending on the adolescent's level of certain personality traits. Shiner and Caspi (2003) outline six possible mechanisms that shape personality development. The unique finding from the current study is that it is not simply one of these processes at work for any given combination of personality trait and aspect of parenting—the type of mechanism depends on the level of the personality trait. For example, the substantial heritability and moderate genetic overlap found between positive emotionality and regard at the extreme high end of personality suggests that at least some of the same genes that influence personality are also influencing the parent-adolescent relationship. This could simply be an example of a more even-tempered adolescent eliciting more positive parenting behaviors, or, at least in part, construing their parents actions in a more positive light. Alternatively, the low heritability and substantial shared environmental influence on regard found at low levels of PEM may indicate a process whereby an adolescent with low levels of positive emotions will compare themselves, possibly unfavorably, to their siblings, and thus his or her relationship with her parents will be based on common factors shared by the parents and all siblings within the household.
One of the most consistent findings from the current analysis was the importance of shared environment, particularly at extremely high or low levels of personality. Even in the no-moderation models, shared environmental effects had a sizeable influence on the variance in Regard, Conflict, and Involvement. These effects were enhanced or diminished depending on the level of the personality trait. For instance, at lower levels of positive (PEM, CN) and higher levels of negative (NEM) personality traits, the amount of Regard in the parent-adolescent relationships was more attributable to factors shared within the family than genetics or unique environment. When an adolescent has a less even-tempered personality, the quality of the parent-adolescent relationship is due largely to factors that are shared in common with other family members. Thus, a more “difficult” teenager may have a relationship with his or her parent which is based on a standard set of rules and interactions shared by all children in the house. Shared environmental influences have been notoriously difficult to detect across a range of phenotypes (Rowe, 1994), leading some to conclude that family has little influence on personality development (Harris, 1995, 1998). The finding of significant shared environmental effects on individual differences in parenting behaviors strengthens the argument for using these new moderation models (Purcell, 2002)—shared environmental effects may be particularly important for certain people within the population, but are lost when estimates are averaged across the whole sample.
It is well-established at this point that most psychological phenomena are heritable (Turkheimer, 2000). The field of behavior genetics must now move beyond static estimates of heritability and genetic and environmental correlations, toward elucidation of how genetic influences and environment transact. The current paper provides evidence that differences in personality can affect the etiology of parent relationship quality. An obvious extension of this work is to examine the dynamic influences of personality traits on relationship quality over time. The stability of personality generally increases from childhood to adulthood (Roberts & DelVecchio, 2000), but personality is already more stable than relationship quality in adolescence (Branje et al., 2004) and young adulthood (Asendorpf & Wilpers, 1998). Further, both personality (McGue, Bacon, & Lykken, 1993) and aspects of family environment (McGue et al., 2005) become more heritable over time. Longitudinal biometric models could help to determine whether the increasing stability in personality is related to expression of genetic variance, and articulate how genetic and environmental transactions unfold over time.
Acknowledgments
This work was supported in part by USPHS grants AA00175, AA09367, and DA05147.
Footnotes
An anonymous reviewer suggested that we augment our results by providing MZ and DZ twin correlations for low versus high levels of personality. If the moderator variable were the same for both twins (e.g., parental marital status or socioeconomic status), then it would be quite easy to calculate twin correlations at different levels of the moderator. However, in this study, the twin pairs are often discordant for level of personality traits (i.e., Twin 1's level of PEM differs from Twin 2's level of PEM). So to calculate a twin correlation at “low versus high levels of personality” is not possible. Indeed, the fact that the biometric moderation models use all individual data, and not covariances between twins, is a strength of the study and why these models are able to calculate ACE variance estimates at different levels of the moderator (Purcell, 2002).
Results for models using the square transformed and truncated data are available from the first author.
Variance component estimates for parenting are presented at five levels of personality, but they could easily be extended to any personality score found in the population.
References
- Akaike H. Factor analysis and AIC. Psychometrika. 1987;52:317–332. [Google Scholar]
- Asendorpf JB, van Aken MAG. Personality-relationship transaction in adolescence: Core versus surface personality characteristics. Journal of Personality. 2003;71:629–666. doi: 10.1111/1467-6494.7104005. [DOI] [PubMed] [Google Scholar]
- Asendorpf JB, Wilpers S. Personality effects on social relationships. Journal of Personality and Social Psychology. 1998;74:1531–1544. [Google Scholar]
- Bates JE, Petit GS, Dodge KA, Ridge B. Interaction of temperamental resistance to control and restrictive parenting in the development of externalizing behavior. Developmental Psychology. 1998;34:982–995. doi: 10.1037//0012-1649.34.5.982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bell RQ. A reinterpretation of the direction of effects in studies of socialization. Psychological Review. 1968;75:81–85. doi: 10.1037/h0025583. [DOI] [PubMed] [Google Scholar]
- Belsky J, Hsieh K, Crnic K. Mothering, fathering, and infant negativity as antecedents of boys’ externalizing problems and inhibition at age 3: Differential susceptibility to rearing influence? Development and Psychopathology. 1998;10:301–319. doi: 10.1017/s095457949800162x. [DOI] [PubMed] [Google Scholar]
- Billig JP, Hershberger SL, Iacono WG, McGue M. Life events and personality in late adolescence: Genetic and environmental relations. Behavior Genetics. 1996;26:543–554. doi: 10.1007/BF02361227. [DOI] [PubMed] [Google Scholar]
- Boomsma D, de Geus E, van Baal G, Koopmans J. A religious upbringing reduces the influence of genetic factors on disinhibition: evidence for interaction between genotype and environment on personality. Twin Research. 1999;2:115–125. doi: 10.1375/136905299320565988. [DOI] [PubMed] [Google Scholar]
- Bouchard TJ, Jr., Loehlin JC. Genes, evolution, and personality. Behavior Genetics. 2001;31:243–273. doi: 10.1023/a:1012294324713. [DOI] [PubMed] [Google Scholar]
- Bouchard TJ, Jr., McGue M. Genetic and rearing environmental influences on adult personality: An analysis of adopted twins reared apart. Journal of Personality. 1990;58:263–292. doi: 10.1111/j.1467-6494.1990.tb00916.x. [DOI] [PubMed] [Google Scholar]
- Burt SA, Krueger R, McGue M, Iacono W. Parent– child conflict and the comorbidity among childhood eternalizing disorders. Archives of General Psychiatry. 2003;60:505–513. doi: 10.1001/archpsyc.60.5.505. [DOI] [PubMed] [Google Scholar]
- Burt SA, McGue M, Iacono WG, Krueger RF. Differential parent-child relationships and adolescent externalizing symptoms: Cross-lagged analyses within a monozygotic twin differences design. Developmental Psychology. 2006;42:1289–1298. doi: 10.1037/0012-1649.42.6.1289. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burt SA, McGue M, Krueger RF, Iacono WG. How are parent– child conflict and childhood externalizing symptoms related over time? Results from a genetically-informative cross-lagged study. Development and Psychopathology. 2005;17:145–165. doi: 10.1017/S095457940505008X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Branje JT, van Lieshout CFM, van Aken MAG. Relations between big five personality characteristics and perceived support in adolescents’ families. Journal of Personality and Social Psychology. 2004;86:615–628. doi: 10.1037/0022-3514.86.4.615. [DOI] [PubMed] [Google Scholar]
- Branje JT, van Lieshout CFM, van Aken MAG. Relations between agreeableness and perceived support in family relationships: Why nice people are not always supportive. International Journal of Behavioral Development. 2005;29:120–128. [Google Scholar]
- Caspi A, Harrington H, Milne B, Amell JW, Theodore RF, Moffitt TE. Children's behavioral styles at age 3 are linked to their adult personality traits at age 26. Journal of Personality. 2003;71:495–513. doi: 10.1111/1467-6494.7104001. [DOI] [PubMed] [Google Scholar]
- Caspi A, Shiner RG. Personality development across the life course. In: Eisenberg N, Damon W, Lerner RM, editors. Handbook of child psychology: Vol. 3, Social, emotional, and personality development. 6th ed. John Wiley & Sons Inc.; Hoboken, NJ: 2006. pp. 300–365. [Google Scholar]
- Chipuer HM, Plomin R, Pedersen NL, McClearn GE, Nesselroade JR. Genetic influence on family environment: The role of personality. Developmental Psychology. 1993;29:110–118. [Google Scholar]
- Collins WA, Maccoby EE, Steinberg L, Hetherington EM, Bornstein MH. Contemporary research on parenting: The case for nature and nurture. American Psychologist. 2000;55:218–232. [PubMed] [Google Scholar]
- Elkins IJ, McGue M, Iacono WG. Genetic and environmental influences on parent-son relationships: Evidence for increasing genetic influence during adolescence. Developmental Psychology. 1997;33:351–363. doi: 10.1037//0012-1649.33.2.351. [DOI] [PubMed] [Google Scholar]
- Harris JR. Where is the child's environment? A group socialization theory of development. Psychological Review. 1995;102:458–489. [Google Scholar]
- Harris JR. The nurture assumption: Why children turn out the way they do. Free Press; New York: 1998. [Google Scholar]
- Heath AC, Eaves LJ, Martin NG. Interaction of marital status and genetic risk for symptoms of depression. Twin Research. 1998;1:119–122. doi: 10.1375/136905298320566249. [DOI] [PubMed] [Google Scholar]
- Hur Y-M, Bouchard TJ., Jr. Genetic influences on perceptions of childhood family environment: A reared apart twin study. Child Development. 1995;66:330–345. [PubMed] [Google Scholar]
- Hur YM, McGue M, Iacono WG. Unequal rate of monozygotic and like-sex dizygotic twin birth: Evidence from the Minnesota Twin Family Study. Behavior Genetics. 1995;25:337–40. doi: 10.1007/BF02197282. [DOI] [PubMed] [Google Scholar]
- Iacono WG, Carlson SR, Taylor J, Elkins IJ, McGue M. Behavioral disinhibition and the development of substance use disorders: Findings from the Minnesota Twin Family Study. Development and Psychopathology. 1999;11:869–900. doi: 10.1017/s0954579499002369. [DOI] [PubMed] [Google Scholar]
- Jocklin V, McGue M, Lykken DT. Personality and divorce: A genetic analysis. Journal of Personality and Social Psychology. 1996;71:288–299. doi: 10.1037//0022-3514.71.2.288. [DOI] [PubMed] [Google Scholar]
- Krueger RF, Markon KE, Bouchard TJ. The extended genotype: The heritability of personality accounts for the heritability of recalled family environments in twins reared apart. Journal of Personality. 2003;71:809–833. doi: 10.1111/1467-6494.7105005. [DOI] [PubMed] [Google Scholar]
- Little RJ, Rubin DB. Statistical analysis with missing data. Second edition John Wiley & Sons; New York: 2002. [Google Scholar]
- Magnusson D. Personality development from an interactional perspective. In: Pervin LA, editor. Handbook of personality: Theory and measurement. Guilford Press; New York: 1990. pp. 193–222. [Google Scholar]
- McCrae RR, Costa PT. A five-factor theory of personality. In: Pervin LA, John OP, editors. Handbook of Personality: Theory and Research. Guilford; New York: 1999. [Google Scholar]
- McGue M, Bacon S, Lykken DT. Personality stability and change in early adulthood: A behavioral genetic analysis. Developmental Psychology. 1993;29:96–109. [Google Scholar]
- McGue M, Bouchard TJ. Adjustment of twin data for the effects of age and sex. Behavior Genetics. 1984;14:325–343. doi: 10.1007/BF01080045. [DOI] [PubMed] [Google Scholar]
- McGue M, Elkins I, Walden B, Iacono WG. Perceptions of the parent-adolescent relationship: A longitudinal investigation. Developmental Psychology. 2005;41:971–984. doi: 10.1037/0012-1649.41.6.971. [DOI] [PubMed] [Google Scholar]
- Muthén LK, Muthén BO. Mplus User's Guide. Fourth Edition Muthén & Muthén; Los Angeles, CA: 19982006. [Google Scholar]
- Neale MC, Boker SM, Xie G, Maes HH. Mx: Statistical Modeling. 6th Edition Department of Psychiatry; VCU Box 900126, Richmond, VA 23298: 2003. [Google Scholar]
- Patterson GR. Coercive family process. Castalia; Eugene, OR: 1982. [Google Scholar]
- Paulussen-Hoogeboom MC, Stams GJJM, Hermanns JMA, Peetsma TTD. Child negative emotionality and parenting from infancy to preschool: A meta-analytic review. Developmental Psychology. 2007;43:438–453. doi: 10.1037/0012-1649.43.2.438. [DOI] [PubMed] [Google Scholar]
- Plomin R, Bergeman CS. The nature of nurture: Genetic influence on “ environmental” measures. Behavioral and Brain Sciences. 1991;14:373–427. [Google Scholar]
- Plomin R, McClearn GE, Pedersen NL, Nesselroade JR, Bergeman CS. Genetic influence on childhood family environment perceived retrospectively from the last half of the life span. Developmental Psychology. 1988;24:738–745. [Google Scholar]
- Purcell S. Variance components models for gene-environment interaction in twin analysis. Twin Research. 2002;5:554–571. doi: 10.1375/136905202762342026. [DOI] [PubMed] [Google Scholar]
- Reiss D, Neiderhiser JM, Heatherington M, Plomin R. The relationship code: Decipering genetic and social patterns in adolescent development. Harvard University Press; Cambridge, MA: 2000. [Google Scholar]
- Roberts BW, DelVecchio WF. The rank-order consistency of personality traits from childhood to old age: A quantitative review of longitudinal studies. Psychological Bulletin. 2000;126:3–25. doi: 10.1037/0033-2909.126.1.3. [DOI] [PubMed] [Google Scholar]
- Rothbart MK, Bates JE. Temperment. In: Eisenberg N, Damon W, Lerner RM, editors. Handbook of child psychology: Vol. 3, Social, emotional, and personality development. 6th ed. John Wiley & Sons Inc.; Hoboken, NJ: 2006. pp. 99–166. [Google Scholar]
- Rowe DC. Environmental and genetic influences on dimensions of perceived parenting: A twin study. Developmental Psychology. 1981;17:203–208. [Google Scholar]
- Rowe DC. A biometrical analysis of perceptions of family environment: A study of twin and singleton sibling kinships. Child Development. 1983;54:416–423. [PubMed] [Google Scholar]
- Rowe DC. The limits of family influence: Genes, experience, and behavior. Guilford Press; New York, NY: 1994. [Google Scholar]
- Rubin KH, Burgess KB, Dwyer KM, Hastings PD. Prediciting preschoolers’ externalizing behaviors from toddler temperament, conflict, and maternal negativity. Developmental Psychology. 2003;39:164–176. [PubMed] [Google Scholar]
- Sameroff AJ. Developmental systems: Contexts and evolution. In: Kessen W, editor. Handbook of child psychology, 4th ed. Vol. 1. History, theory, and methods. Wiley; New York: 1983. pp. 237–294. [Google Scholar]
- Saudino KJ, Pedersen NL, Lichtenstein P, McClearn GE, Plomin R. Can personality explain genetic influences on life events? Journal of Personality and Social Psychology. 1997;72:196–206. doi: 10.1037//0022-3514.72.1.196. [DOI] [PubMed] [Google Scholar]
- Scarr S, McCartney K. How people make their own environments: A theory of genotype-environment effects. Child Development. 1983;54:424–435. doi: 10.1111/j.1467-8624.1983.tb03884.x. [DOI] [PubMed] [Google Scholar]
- Shiner R, Caspi A. Personality differences in childhood and adolescence: measurement, development, and consequences. Journal of Child Psychology and Psychiatry. 2003;44:2–32. doi: 10.1111/1469-7610.00101. [DOI] [PubMed] [Google Scholar]
- Stoolmiller M. Synergistic interaction of child manageability problems and parent-discipline tactics in predicting future growth in externalizing behavior for boys. Developmental Psychology. 2001;37:814–825. doi: 10.1037//0012-1649.37.6.814. [DOI] [PubMed] [Google Scholar]
- Tellegen A, Waller NG. Exploring personality through test construction: Development of the Multidimensional Personality Questionnaire (MPQ) University of Minnesota Press; Minneapolis: in press. [Google Scholar]
- Turkheimer E. Three laws of behavior genetics and what they mean. Current Directions in Psychological Science. 2000;9:160–164. [Google Scholar]
- Turkheimer E, Haley A, Waldron M, D'Onofrio B, Gottesman II. Socioeconomic status modifies heritability of IQ in young children. Psychological Science. 2003;14:623–628. doi: 10.1046/j.0956-7976.2003.psci_1475.x. [DOI] [PubMed] [Google Scholar]