
Fig. 4.
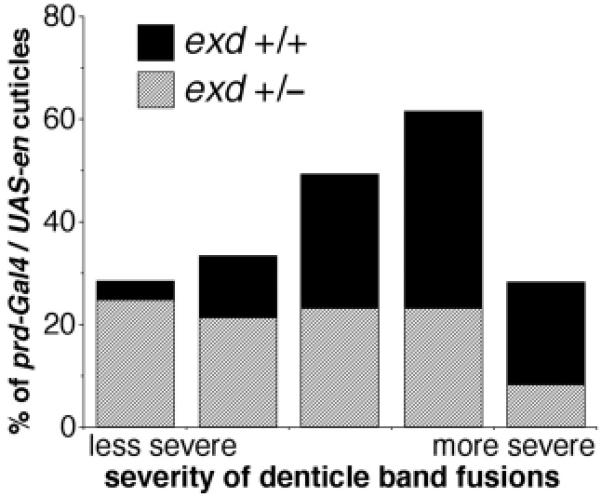
Flies with exd mutations show dose-dependent interactions with ectopically patterned En in embryos. Embryos collected as in Figs 2 and 3 were allowed to develop to the end of embryogenesis, and cuticles were prepared and analyzed. Defects in abdominal denticle bands (mostly pairwise fusions of varying severity) were categorized and tabulated, and the results are shown in a stacked bar graph. As indicated by the percentage of defective cuticles in both the exd+/+ population and the population derived from exd mutant germline clones (data not shown), those embryos that did not receive a copy of the prd-Gal4 driver showed no defects other than those expected from the complete absence of exd function (embryonic cuticles from exd germline clones are completely rescued by one wild-type gene from the father). We did not attempt to analyze exd-null cuticles (which were clearly distinguishable from the exd heterozygotes) for effects of En ectopic expression induced by the driver. Rather, the denticle band fusions caused by ectopic En expression in the population that received a wild-type exd allele from their father were analyzed, and the graph shows the percentage of defects in each category among this population (which are exd+/-, as indicated in the key). Thus, the percentages add to 100% in each case because they include only those cuticles that showed defects caused by ectopic En expression that were of the indicated exd genotype (in each case, these represented the expected overall percentage of cuticles). Notice that when the maternal contribution of exd is removed and the zygotic contribution is simultaneously reduced by half (exd+/-), the overall severity of abdominal cuticle defects in the population caused by ectopic En expression is significantly reduced, indicating a substantial requirement for Exd in En function in the developing abdomen.