

Abstract
Many systems in chemistry, biology, finance, and social sciences present emerging features that are not easy to guess from the elementary interactions of their microscopic individual components. In the past, the macroscopic behavior of such systems was modeled by assuming that the collective dynamics of microscopic components can be effectively described collectively by equations acting on spatially continuous density distributions. It turns out that, to the contrary, taking into account the actual individual/discrete character of the microscopic components of these systems is crucial for explaining their macroscopic behavior. In fact, we find that in conditions in which the continuum approach would predict the extinction of all of the population (respectively the vanishing of the invested capital or the concentration of a chemical substance, etc.), the microscopic granularity insures the emergence of macroscopic localized subpopulations with collective adaptive properties that allow their survival and development. In particular it is found that in two dimensions “life” (the localized proliferating phase) always prevails.
In addition to physics, an increasing range of sciences (chemistry, biology, ecology, finance, urban and social planning) have moved in the last century to quantitative mathematical methods.
Along with the obvious benefits, it turns out that the traditional differential equations approach has brought some fallacy into the study of such sciences.
We present here a very simple generic model that contains proliferating (and dying) individuals, and we show that in reality it behaves very differently than its representation in terms of continuum density distributions. In conditions in which the continuum equations predict the population extinction, the individuals self-organize in spatio-temporally localized adaptive patches, which ensure their survival and development.
This phenomenon admits multiple interpretations in various fields:
If the individuals are interpreted as interacting molecules, the resulting chemical system emerges spatial patches of high density that evolve adaptively in a way similar with the first self-sustaining systems that might have anticipated living cells.
If the individuals are the carriers of specific genotypes represented in the genetic space, the patches can be identified with species, which rather than becoming extinct, evolve between various genomes (locations in the genetic space) by abandoning regions of low viability in favor of more viable regions. This adaptive speciation behavior emerges despite the total randomness we assume for the individuals motions in the genetic space (mutations).
Interpreted as financial traders, the individuals develop a “herding” behavior despite the fact that we do not introduce communication or interaction between them. The model leads to the flourishing of markets, which the continuum analysis would doom to extinction.
All these phenomena have in common the emergence of large, macroscopic structures from apparently uniform background (1) caused by the amplification of small, microscopic fluctuations that originate in the individualized character of the elementary components of the system. This mechanism insures in particular that on large enough two-dimensional surfaces, even if the average growth rate is negative (because of very large death rate), adaptive structures always emerge and flourish.
Imagine an area inhabited by a population of eternal agents A, which are spread out uniformly with average density nA and move around randomly, with diffusion coefficient DA. Imagine now a race of mortals, B, which also are spread over this area, with initial uniform density nB(0). The B agents die at a constant rate, μ, (B→μ) and proliferate (divide) when they meet the “catalyst,” A, with rate λ (B + A→λB + B + A). The Bs are diffusive, hopping at the rate DB. What will happen?
The naive lore based on macroscopic continuity assumptions will predict that A reaches a spatially homogeneous distribution, nA(x) = nA, while the B time variation ∂nB/∂t is represented by the linear differential partial differential equation:
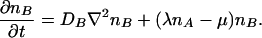 |
1 |
The first term represents the uniformization effect of B diffusion while the μB term represents the fact that a certain fraction of Bs die per unit time. The crucial term λnAnB represents the proliferation of Bs in the presence of the “life giving” As. Note that the equation is linear in nB and that for initial spatially uniform nA and nB distributions it has the time exponential solution
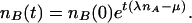 |
2 |
In particular, Eq. 2 predicts that if the macroscopic proliferation rate λnA is lower than the death rate μ, the B population will uniformly decrease to extinction.
Using microscopic representation techniques (2), one finds that populations of discrete proliferating agents are much more resilient than one would first guess based on macroscopic or continuum (partial differential equations) treatment (Fig. 1).
Figure 1.

(Lower) Shown is the exponential growth of the average B population as a function of time, in the actual simulation (solid blue line) compared with the exponential decrease predicted by the continuum approximation (dashed red line). <A> is the average number of A reactants per site. Other symbols are defined in the text. (Upper) The snapshots show the spatial configuration of A and B reactants. B reactants are seen to be localized in islands (notice that what is plotted is the logarithm of B concentrations, thus localization is stronger than first would appear).
The study of diffusion limited reactions (3) already has shown in the past deviations from the continuum theory because of the quantized nature of the reactants. In the present case the effect is even more dramatic. It constitutes the difference between life and death. The continuum approach predicts extinction whereas the direct simulation uncovers the emergence of a thriving, adaptive, developing system (Fig. 1). These simulations were carried out in both synchronous and asynchronous algorithm with no apparent effect on the results (4).
To understand the “source of life” in this system one has to concentrate on the microscopic conditions around the individual A agents, rather then looking at the local average growth rate λnA − μ.
Fig. 2 represents the evolution of the B cloud following a single A agent as it jumps around randomly. The B concentration is shown to trace the A as it performs a random walk. Clearly, the colony does not decay to extinction; instead, it seems to trace the A, “trying” to keep its center of mass at its location. As shown in Fig. 2 Inset, each jump of the A is followed by a momentary decrease in the height of the B concentration; however, because of the multiplicative process there is an overall increase.
Figure 2.

The profile of a B island as a function of time as it follows the random motion of an A agent. The cross-section of the island is taken through the current location of the A agent. (Inset) The time evolution of the height of the B concentration at the point at which A currently is located (solid blue line). The B colony is seen to grow, although the average growth rate over the entire space is negative (nA is extremely low because there is only one in the whole simulation space, thus λnA − μ ≈ −μ). The dashed red line shows the exponential growth with coefficient ɛ0 − 2dDAκ, where κ = Loge(λ/DB) is the slope of the island (this slope is exhibited in the main graph and can be derived from a simple approximate calculation). ɛ0 is λ − μ − 2dDB derived similarly.
Let us consider first the simplest situation of a single A agent jumping randomly (with a diffusion coefficient DA) between the locations in an infinite d-dimensional space. In between A jumps the B density at the A location grows exponentially (5, 6) as nB(t) ≈ nB(0)e(λ−μ−2dDB)t. Where λ, μ, and 2dDB stand for the proliferation, death, and the loss caused by diffusion, respectively. The estimation is made by neglecting the flow of Bs from a neighboring site to the A site. This approximation is justified when the B concentration in the neighboring sites is much lower than on the A site. In the same limit, the ratio between the height of the B density at the A location and the height of the B density on a neighboring site is easily estimated: λ/DB. Consequently, each A jump corresponds to a sudden downward jump by a factor of λ/DB in the height of the B hill. As there are on average 2dDA such jumps per unit time, the net effect of proliferation, diffusion, and death gives the B concentration at the A site as a function of time:
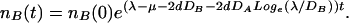 |
3 |
The approximation is in good agreement with the simulation shown in Fig. 2. The slope of the island, on a log scale, is indeed seen to be Loge(λ/DB). The time dependence of the height of the B island in between A jumps is indeed given approximately by an exponent (λ − μ − 2dDB)t. Consequently the dashed red line (Fig. 2 Inset), which represents Eq. 3, follows closely the actual growth seen in the simulation (blue line). The difference between the theory and simulation is mainly caused by cases where two or more A jumps follow each other rapidly, in this case the island's shape does not stabilize before another A jump is made, these rather rare events, modify somewhat the actual result. This analysis turns void if DB = 0, where the spatial dimensions of the island do not grow at all. We do not consider this singular case in this paper.
One may ask what is the situation in the case when single colonies are unstable (i.e., where the exponent in Eq. 3 is negative). One possibility is that in such a situation the continuum approximation is valid and the B concentration decays to zero. Another possibility is that, although single isolated colonies are unstable, global effects such as islands growing, joining, and splitting give us back the survival feature. In particular, because large colonies are more stable than small colonies, one may expect the typical size of an “active” colony to grow with time. This behavior is demonstrated in Fig. 3, which shows the active clusters in a two-dimensional system developing in time. Evidently, the small clusters either decay or merge into larger and larger clusters.
Figure 3.

The spatial distribution of Bs for different times (earlier times to the left), where concentrations greater than 10 are colored red and concentrations below 10 are color coded.
In many natural situations, the proliferation of some entity takes place under the influence of a discrete catalyst. Assuming that our model captures the dynamic of such living systems correctly, the conclusion may be that individual-based life is much more resilient than a hypothetical life density spread continuously across spatial regions.
These conclusions should suffice to induce professionals in biology (7, 8) and in finance and social sciences (9) to consider seriously the regime in which their systems are naively nonviable (decay to extinction) when viewed macroscopically but perfectly viable in reality (and when simulated correctly at the microscopic individual level; ref. 10).
In particular, most of the species in nature could be in this regime: negative naive average macroscopic growth rate but actual survival and proliferation. Similarly, markets that might look unappealing when averaging over the various investing possibilities might prove lucrative enough (at least for the lucky investors that hit profit opportunities A) as to maintain them in the competitive range. In fact this line of thought might provide an explanation to the emergence of life from the random chemistry of its component molecules despite the formal extreme improbability of the event. Equally it might explain the paradoxes in finance between the efficient market hypothesis (absence of systematic profit opportunities in equilibrium markets) and the actual profits that investors extract daily from the market.
We note that more realistic models that can be studied by using the methods described in this paper can be constructed. Such studies could account for the finite carrying capacity of the environment, as well the consumption of resources and competition between different species.
To obtain a more rigorous bound on the parameter range in which life overcomes the gloom prognosis of the macroscopic analysis we used the renormalization group analysis, which indicates that on large enough surfaces life always wins. For higher dimensions, the dominion of life still extends to arbitrary low nA densities, but a minimal finite λ value is required.
In the renormalization group formalism, the collective behavior of the system is identified by integrating out the small length scale, short time fluctuations, leaving us with an effective theory for the large-scale objects. Here, these are the large, stable islands shown in Fig. 3. The new, effective theory is characterized by renormalized coupling constants, i.e., modified numerical values of the effective rates (growth rate, dearth rate, hopping, etc.) on a large length scale. The process of decimating small fluctuations then is iterated again and again, giving us a flow line that reflects the evolution of the effective values of the coupling constants as one integrates larger and larger scales l. The details of this renormalization group analysis, which involves the presentation of the exact master equation of the process as a field integral and the ɛ-expansion around the critical dimension dc = 2 are out of the scope of this report and will be presented elsewhere.
Fig. 4 shows the flow lines of m (m = μ − λnA) and λ caused by the iteration of the decimation process. D ≡ DA + DB is the effective diffusion constant. The flow is given by:
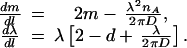 |
4 |
For d ≤ 2 we see that for large length and time scales (that is, after many iterations of the decimation process), λ grows without limit whereas m eventually becomes negative. This finding implies that on the large scale, the system actually behaves as if λnA > μ, and life always wins.
Figure 4.

(Lower) Flow lines for d > 2. Shaded region flows to negative mass (“life”). (Upper) Flow lines for d ≤ 2. The whole parameter space flows to negative mass.
In higher dimensions (d > 2) Fig. 4 Upper indicates a dynamical phase transition where for part of the parameter space the system flows to negative m (life) and for another part the system flows to positive m (death).
It should be noted that the flow portrayed in Fig. 4 is associated with larger and larger length scales. For a finite system, the flows should be truncated and the size of the system may be crucial. Simulations with parameters identical to that of Fig. 3 lead to extinction when carried out on a system size four times smaller.
In conclusion, our results suggest that the dimensionality of the system and its size are crucial features for its capability to emerge and sustain life. This may explain the fact that most ecological systems are two-dimensional. Reinterpreting in the genome space, the present results provide the conceptual link between the atomized structure of the life building blocks and the explosive Darwinian tandem, noise + proliferation.
Footnotes
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.180263697.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.180263697
References
- 1.Anderson P W. Phys Rev. 1958;109:1492–1505. [Google Scholar]
- 2.Solomon S. In: Annual Reviews of Computational Physics II. Stauffer D, editor. Teaneck, NJ: World Scientific; 1995. pp. 243–294. [Google Scholar]
- 3.Mattis D C, Glasser M L. Rev Mod Phys. 1998;70:979–1001. and references therein. [Google Scholar]
- 4.Huberman B, Glance N. Proc Natl Acad Sci USA. 1993;90:7716–7718. doi: 10.1073/pnas.90.16.7716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Nelson D R, Shnerb N M. Phys Rev E Stat Phys Plasmas Fluids Relat Interdiscip Top. 1998;58:1383–1403. [Google Scholar]
- 6.Dahmen, K. A., Nelson, D. R. & Shnerb, M. N. (1998) e-Print Archive, http://xxx.lanl.gov/abs/cond-mat/9807394.
- 7.Allen P, McGlade J. Found Phys. 1987;17:723–738. [Google Scholar]
- 8.Smith J M. Mathematical Ideas in Biology. Cambridge, U.K.: Cambridge Univ. Press; 1968. [Google Scholar]
- 9.Winder N. In: Policy-Relevant Models of the Natural and Anthropogenic Dynamics of Degradation and Desertification and Their Spatio-Temporal Manifestation: Second Report of the Archaeomedes Project (II) for the Directorate General XII of the European Union. van der Leeuw S, editor. 1998. pp. 217–235. [Google Scholar]
- 10.Winder, N. (2000) Hum. Ecol. Rev., in press.