
Figure 1. Mechanisms of cellular gene silencing.
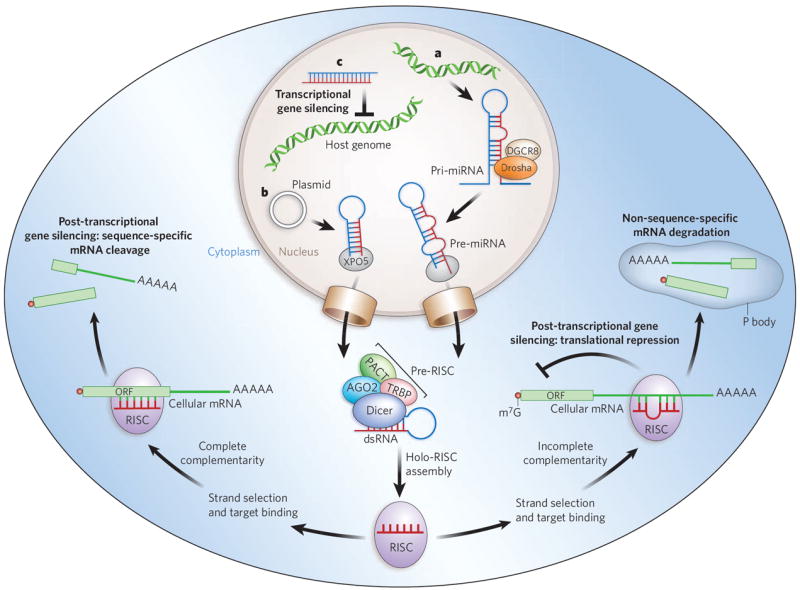
a, Primary microRNAs (pri-miRNAs) are, in plants and animals, processed by Drosha and its partner DGCR8 into precursor miRNAs (pre-miRNAs) and then transported to the cytoplasm by exportin 5 (XPO5). In the cytoplasm, they are bound by a Dicer-containing pre-RISC and processed to yield the guide sequence that is loaded into the holo-RISC, which contains all the components required for gene silencing. AGO2 is the catalytic core of the RISC (present but not shown in the schematically drawn holo-RISC). The guide sequence binds to the corresponding target sequences in the 3′ UTRs of cellular mRNAs. If the miRNA guide sequence is fully complementary to its target site (left pathway), it triggers site-specific cleavage and degradation of the mRNA through the catalytic domain of AGO2. If the base-pairing is incomplete (right pathway) but includes pairing of the seed region (nucleotides 2–8 of the miRNA) with the target, translational inhibition occurs, and this can be accompanied by non-sequence-specific degradation of the mRNA in P bodies. b, Similarly to miRNAs, artificially transcribed shRNAs (in this case from a plasmid) are transported to the cytoplasm by XPO5. The dsRNA in the cytoplasm is recognized and processed by Dicer into ~21–25-nucleotide siRNA fragments that are loaded into the RISC. The siRNAs can target complementary sequences of cellular mRNAs and trigger their degradation through AGO2-mediated cleavage. c, When siRNAs are present in the nucleus and are complementary to promoter regions, they can trigger chromatin remodelling and histone modifications that result in transcriptional gene silencing. In mammalian cells, the details of this mechanism are still under investigation but are known to include Argonaute-family proteins. Accessory proteins indicated in the figure are TRBP (HIV tar-RNA-binding protein; also known as TRBP2P) and PACT (activator of protein kinase PKR; also known as PRKRA). m7G, 7-methylguanosine.