

Abstract
Although several studies have previously focused on the conjunctival epithelial response to surface dryness, little is known about the effect of a dry environment on corneal epithelium, which is the most clinically significant tissue affected in dry eye. The aim of this study was to quantitatively evaluate the effect of desiccating stress on the number of proliferating corneal epithelial cells and corneal epithelial thickness in mice placed in a controlled-environment chamber (CEC) that induces dry eye.
Corneal epithelial cell proliferation and thickness were studied in 8–12 week old female BALB/c mice placed in the CEC (temperature: 22.3 ± 0.7°C; relative humidity: 22.5 ± 4.5%; airflow: 15 L/min) for 7 days and compared to a control group of mice with no dry eye. Actively proliferating cells were identified by immunofluorescence using a FITC-conjugated antibody against the Ki-67 protein, a cell proliferation marker expressed during active phases of the cell cycle. To detect the spatial distribution of proliferative cells, Ki-67+ cells were counted in three areas of the epithelium: center, periphery, and limbus. Corneal epithelial thickness was evaluated in the central cornea after staining with hematoxylin-eosin. Results from each experimental group were compared using the Mann-Whitney test.
The number of Ki-67+ cells observed in the corneal epithelium of mice exposed to the CEC was significantly higher in each area (center: 32.1 ± 1.1; periphery: 94.2 ± 5.3; limbus: 4.0 ± 1.5) than in the control group (center: 13.2 ± 1.0, p = 0.02; periphery: 42.9 ± 2.3, p = 0.02; limbus: 0.0, p = 0.01). In mice subjected to desiccating stress, a significant number of Ki-67+ positive cells were detected in the basal and suprabasal cell layers (central area 46%; periphery 30.8%: limbus 0%), whereas in the control group the cells were exclusively distributed through the basal cell layer. Ki-67+ cells were not found in the corneal stroma or endothelium in any group. The corneal epithelium was found to be significantly thicker in dry eye mice (54.94 ± 6.09 μm) as compared to the controls (43.9 ± 6.23 μm: p < 0.0001) by a mean of 25%.
These results demonstrate that desiccating stress increases corneal epithelial turnover and thickness, similar to what is observed in other chronic inflammatory states of other epithelialized surfaces. The CEC can facilitate the study of the regulation of epithelial cell function and turnover at the molecular and cellular levels under desiccating stress conditions.
Keywords: dry eye, animal model, corneal epithelium, cell proliferation
1. Introduction
The outermost surface of the cornea consists of a squamous, non-keratinizing epithelium, which serves as a protective barrier against fluid loss and pathogen penetration, and provides a transparent medium for vision. A continuous replenishment of epithelial cells is fundamental to guarantee these functions. Under normal circumstances, replenishment of the corneal epithelium originates in the limbus, an anatomical niche where epithelial stem cells are located. Stem cells are relatively undifferentiated, slow-cycling and have a large capacity to divide, producing a progeny of basal transient amplifying (TA) epithelial cells that migrate toward the center of the cornea (Pellegrini et al., 1999). TA cells have shorter cell cycle times that allow for faster cell divisions but have a more limited proliferative capacity (Pellegrini et al., 1999; Wilson et al., 1999). Peripheral (young) TA cells are capable of multiple divisions while central mature TA cells only have one remaining division (Lehrer et al., 1998; Pellegrini et al., 1999). According to Thoft’s so-called “X, Y, and Z” hypothesis, subsequent cell divisions of TA cells give rise to post-mitotic cells (PMC), which occupy the suprabasal layer, and progressive differentiation of these PMC results in terminally differentiated cells in the superficial layers that finally exfoliate (Thoft and Friend, 1983). The epithelium completely turns over in approximately 7 days under normal conditions (Hanna et al., 1961; Pflugfelder et al., 1999). Upon stimulation the corneal epithelium can adopt three strategies to expand its cell population: recruitment of stem cells to produce more TA cells; increasing the number of times a TA cell can replicate; and/or increasing the efficiency of TA cell replication by shortening the cell cycle time (Lehrer et al., 1998; Pellegrini et al., 1999). Several factors (e.g., tear film, neuronal innervation, reflex blinking) help to sustain an environment that is favourable for corneal epithelial turnover. Although previous reports have shown that alteration of these factors in dry eye results in abnormal proliferation and differentiation of the conjunctival epithelium (Dursun et al., 2002; Jones et al., 1998; Kunert et al., 2002), little is known about their effect on corneal epithelial proliferation and thickness.
Recently, a mouse model of evaporative dry eye has been developed using a controlled-environment chamber (CEC) that allows for control and monitoring of temperature, airflow, and humidity, and regulation of these parameters in a manner that leads predictably to a dry eye state in mice, characterized by corneal fluorescein staining, loss of conjunctival goblet cells and diminished lacrimation (Barabino et al., 2005). The initial characterization of this model in normal mice has shown that the CEC can induce ocular surface signs of dry eye, with features that mimic human keratoconjunctivitis sicca (KCS) (Barabino et al., 2005).
The aim of this study was to quantitatively evaluate the effect of experimentally induced dry eye using the CEC on the number of proliferating corneal epithelial cells in different portions of the corneal epithelium, and to examine corneal epithelial thickness in the dry eye state.
2. Materials and methods
2.1 Animals
Cell proliferation and thickness of the corneal epithelium were studied in 8–12 week old female BALB/c mice (Taconic Farms, Germantown, NY) placed in the CEC for 7 days, as recently described (Barabino et al., 2005). Animals in the CEC were compared to a control group. Cell proliferation was studied in 8 animals (4 per group). Corneal morphology and central corneal thickness were evaluated in 12 mice placed in the CEC and 10 normal mice. The protocol was approved by the Schepens Eye Research Institute Animal Care and Use Committee, and all animals were treated according to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research.
2.2 Controlled Environment Chamber (CEC)
The CEC has been previously described (Barabino et al., 2005). Briefly, the overall mean relative humidity (RH) in the CEC was 22.5 ± 4.5%, and the air flow maintained constant at 15 l/min for 7 days. The recorded temperature was in the range of 22.3 ± 0.7°C. Animals demonstrated normal behavior similar to that of littermates in the standard cages.
2.3 Immunofluorescence
After sacrificing the animals, eyes were excised and embedded in OCT compound (Tissue-Tek; Miles Laboratories; Elkhart, Ind) and frozen at −80 °C. Sagittal sections of 8 μm were cut with a cryostat and placed on glass slides. Sections were then processed for immunohistochemical and morphological evaluation. Actively proliferating cells were identified by immunofluorescence using a FITC-conjugated mouse monoclonal antibody against the Ki-67 protein (Novocastra Laboraties, Newcastle, UK), a cell proliferation marker expressed during active phases of the cell cycle (Brown and Gatter, 2002). Slides were fixed in Acetone, rinsed in PBS, and blocked with 1% bovine serum albumin (BSA). The antibody was diluted 1:100 and placed on the sections. The incubation time for the antibody was one hour at room temperature. Cover slips were mounted and DAPI was used to mark the nuclei of all cells. A negative control experiment was run using an isotype-matched antibody; positive controls were lymphonodal tissue. To quantify alterations in Ki-67-expressing cells, sections were viewed and photographed with a Nikon Eclipse E800 microscope equipped with a digital SPOT camera. Labelled cells were counted through the entire epithelial thickness in five zones: a central area, two peripheral areas and two limbal areas. The central area was defined as the area within 0.75 mm of the corneal center; the periphery was defined as being 0.75 to 1.5 mm radial distance from the center. The limbus was defined as the intervening zone between the cornea and conjunctiva, slightly more than 1.5 mm radially from the corneal center. The number of Ki-67+ cells was counted in 4 sections per cornea.
2.4 Corneal Morphology and Epithelial Thickness
Corneal sections of 3 μm were stained with hematoxylin-eosin. One masked observer evaluated the corneal morphology and the corneal epithelium thickness of the central area under a microscope (Nikon Eclipse E800, Melville, NY) with a 20x objective. Central corneal epithelial thickness was evaluated in 22 corneas (12 dry eye murine corneas; 10 normal murine corneas), 5 sections for each cornea.
2.5 Statistical Analysis
Changes in the number of Ki-67-positive cells and measurements of central corneal epithelial thickness were summarized using descriptive statistics (ie, sample size, mean, standard deviation, minimum, maximum, and median). The nonparametric Mann-Whitney test was performed to evaluate the differences between the study and control groups; Wilcoxon ranked test was used to compare each region and to evaluate the difference between basal and suprabasal compartments. The statistical analysis was performed with commercial software (SPSS for Windows, ver.9.0; SPSS Sciences, Chicago, IL). All tests were two-tailed, and p < 0.05 was considered to be statistically significant.
3. Results
3.1 Corneal Epithelial Proliferation
Cycling Ki-67+ cells were present in the corneal epithelium of both normal and dry eye corneas. The proliferation rate in the corneal epithelium of mice exposed to the CEC was higher (center: 32.1 ± 1.1; periphery: 94.2 ± 5.3; limbus: 4.0 ± 1.5) than in the control group (center: 13.2 ± 1.0, p=0.02; periphery: 42.9 ± 2.3, p=0.02; limbus: 0.0, p=0.01). Ki-67+ cells were found in the limbus of dry eye corneas, but not in the normal limbal epithelium. The adjacent peripheral epithelium showed the highest degree of labelling in both groups. Figure 1 and 2 are representative images of these data.
Figure 1.

Immunofluorescence staining of central area of normal (top) and dry (bottom) eyes. Actively proliferating cells were identified by immunofluorescence using a FITC-conjugated antibody against the Ki-67 protein (green); coverslips were mounted with DAPI to mark the nuclei of all cells (red); photographs were taken and merged to recognize the cycling cells (orange). Ki-67+ cells were not found in the corneal stroma or endothelium in any group. All photographs were taken at the same magnification.
Figure 2.

Proliferating cells in central, peripheral and limbal corneal epithelium in normal (left) and dry (right) eye mice. The number of Ki-67+ cells was significantly higher in each individual area (center; periphery; limbus), as compared to the control group. The peripheral epithelium had the highest degree of labelling in both groups. Ki-67+ cells were found in the limbus of dry eye corneas, but not in the normal epithelium. Data show median ± interquartile; Mann-Whitney test.
Ki-67+ cells were subdivided into two compartments: basal and suprabasal. In the control group without dry eye, Ki-67+ cells were exclusively distributed through the basal cell layer (Figure 3). In mice subjected to desiccating stress, a significant number (34.2%) of Ki-67+ cells were detected in the suprabasal cell layers. Of those cells, the number of suprabasal cells in the periphery was 30.8%, whereas 46% was found in the central cornea. No suprabasal Ki67+ cell was detected in the limbus of dry eye mice. Ki-67+ cells were not found in the corneal stroma or endothelium in any group (Figures 1 and 3).
Figure 3.
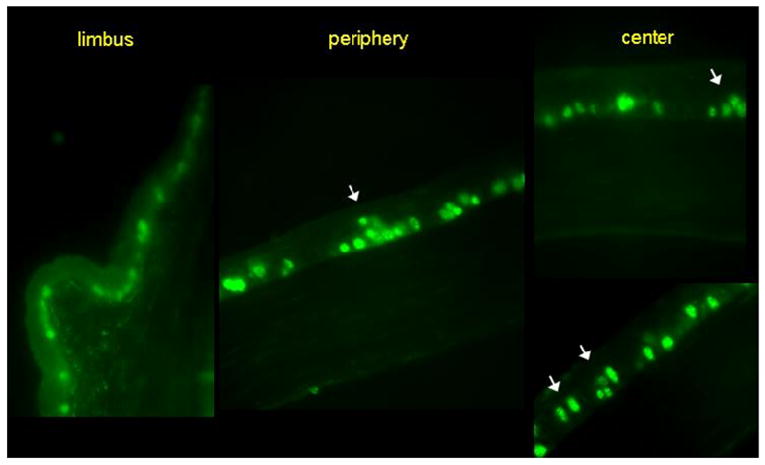
Ki-67+ cells distribution among the corneal epithelium in mice subjected to desiccating stress (green). A significant number of Ki-67+ cells (34.2%) were detected in the suprabasal cell layers of the central and peripheral corneal epithelium (white arrows). No suprabasal cell was detected in the limbus of dry eye mice.
3.2 Corneal Epithelial Morphology and Thickness
As shown by hematoxylin-eosin staining, the corneal epithelium in dry eye mice had a hyperplastic appearance with flattened and keratinized cells in the apical portion, and was comprised of six or more epithelial cell layers (acanthosis). In contrast to the apical cells, basal and suprabasal cells had a cuboidal shape and appeared to be smaller as compared to the control non-dry eye group (Figure 4, Panel A).
Figure 4.

Corneal epithelial thickness: comparison between normal and dry eye mice. A. Central corneal sections of 3 μm were stained with hematoxylin-eosin (H&E). Normal control cornea had a stratified epithelium of 4 to 6 cell layers and an avascular stroma with cells aligned in a lamellar manner. After 7 days of exposure to the CEC epithelial stratification was marked in the central regions. Besides marked stratification, the most striking finding was the increased cellularity and the general hyperplastic appearance of cells, evidenced by the cuboidal shape of postmitotic suprabasal cells (photographs were taken at the same magnification). B. Measurement of the central part of the corneal epithelium revealed that the epithelium was significantly thicker in eyes of dry eye mice as compared to the controls. Data show median ± interquartile; Mann-Whitney test.
Measurements of the central part of the corneal epithelium revealed that the epithelium was 25% thicker in dry eye mice (54.94 ± 6.09 μm) as compared to controls (43.9 ± 6.23 μm, p < 0.0001) (Figure 4, Panel B).
4. Discussion
The present study demonstrates that exposure to a desiccating stress in a mouse model of experimental dry eye significantly increases the cell proliferation rate and the central thickness of the corneal epithelium. The presence of Ki-67+ cells in the basal and suprabasal areas of the corneal epithelium in this model suggests that drying conditions stimulate cells to cycle and proliferate through the entire epithelium as a consequence of apical cell surface damage. Consequently the epithelium acquires a hyperplastic appearance, evidenced by the cuboidal shape of postmitotic suprabasal cells, and the increase in the number of cell layers.
Several studies have previously focused on the conjunctival epithelial response to surface dryness. Dursun et al. (2002) showed an increase in epithelial proliferation in the conjunctiva of mice in which dry eye was induced by desiccating stress in combination with pharmacological inhibition of lacrimal secretion using scopolamine. Jones et al. (1998) demonstrated that the conjunctival epithelium of Sjögren’s Syndrome patients is characterized by abnormal proliferation and differentiation; similar findings were reported by Kunert et al. (2002) in patients with non-Sjögren’s dry eye. These authors also found the conjunctival epithelium to be thicker, and described an altered histological appearance. The cornea, however, is the most clinically significant tissue affected in dry eye states as its normal functioning is critically related to central vision; conversely, its morbidity accounts for the most severe consequences of dry eye syndrome. As such, to our knowledge, this is the first study that evaluates the proliferative response to desiccating stress and the thickening of the corneal epithelium, and suggests that abnormally rapid tissue turnover and hyperplasia is a common response to the epithelial “stress response” throughout the entire ocular surface. Previous studies in humans have focused on total corneal, not just epithelial, thickness, and have reported a reduction in central corneal thickness in Sjögren’s and non Sjögren’s dry eye; in these studies the central thickness of the corneal epithelium did not change (Erdélyi et al., 2007; Tuominen et al., 2003) or was not evaluated (Liu and Pflugfelder, 1999; Sanchis-Gimeno et al., 2005; Villani et al., 2007).
The precise reasons behind, or mechanisms that lead to, the increased epithelial turnover and thickness in our model of experimental dry eye are not fully understood. Whether due to hyperosmolar stress, or the microabrasive effect of blinking on an ocular surface epithelium devoid of its normal protective barriers (Li et al., 2004; 2006), the higher renewal rate as evidenced by our study may be regarded as a compensatory measure against dry eye-induced damage (or induced apoptosis), and may have significant implications for the health and maintenance of the ocular surface. It has been proposed that chronic environmental stress may stimulate ocular surface sensory nerve endings, serving the whole ocular surface, inducing neurogenic inflammation (Beuerman and Stern, 2005). Whether neurogenic or not, there is ample and increasing evidence demonstrating that dry eye may be a cause of ocular surface inflammation, that in turn can amplify the disease pathology (Brignole et al.. 2000; 2001). Several studies have demonstrated increased levels of inflammatory cytokines, such as IL-1α and IL-1β, TNF-α, IL-6 and IL-8 in dry eye (Gamache et al., 1997; Jones et al., 1998;Pflugfelder et al., 1998; Tishler et al., 1998; Turner et al., 2000). Data from both animal models and human clinical samples have suggested the critical involvement of several pro-inflammatory cytokines in epithelial hyper-proliferation, and keratinisation (Hirai et al., 2006; Tseng et al., 1984). Several of these cytokines such as IL-6 and IL-8 have been reported to influence the growth of epithelial cells, and to promote hyper-proliferation of the epidermis in psoriasis (Denda et al., 1998; Gillitzer et al., 1994; Wilson and Kim, 1998). Additionally, IL-1, which is amply overexpressed in a variety of dry eye states (Pflugfelder et al., 1999; Solomon et al., 2001) can lead to epithelial cell proliferation and keratinisation and can induce apoptosis of corneal stromal and dermal fibroblasts in vitro (Dana et al., 1998; Solomon et al., 2001; Wilson et al., 1996) making it a likely culprit, at least in part, for some of the changes we have observed in the corneal epithelium.
Abnormal proliferation and differentiation of the conjunctival epithelium (Jones et al., 1998; Pflugfelder et al., 1990; Yeh et al., 2003) has been correlated to cell cycle dysregulation induced by abnormal expression of proteins that can amplify or induce apoptosis, such as TNF superfamily gene products CD40 and CD40 ligand, Fas and Fas ligand, and APO2.7 (Brignole et al., 2000; 2001; Stern et al., 2004). Yeh et al. (2003), in their experimentally induced dry eye model comprised of systemic administration of scopolamine and exposure to a strong air draft, found apoptosis of cells in the ocular surface epithelia including the central and peripheral corneal epithelium, and suggested that apoptosis may play an important role in the pathogenesis of KCS-related epitheliopathy. Excessive apoptosis, or shedding, of the surface epithelium, if sustained and not compensated for any epithelial cycling, may in due course lead to epithelial thinning or defects, commonly seen in severe chronic dry eye states. It is therefore possible, as we hypothesize, that some degree of compensatory hyper-proliferation can balance, especially in the early phase of the disease, the increased levels of apoptosis as demonstrated by Yeh et al (2003), allowing for maintenance of the epithelial barrier. Thus, it is possible that desiccating stress is triggering changes in gene expression that induces increased cell proliferation as a “counter-balance” to activation of pro-apoptotic factors. The increased thickness of the corneal epithelium in our experimentally induced dry eye may be a consequence of this active compensatory hyperproliferation along with a decreasing shedding rate in the acute phase of the disease. A trophic breakdown of the corneal epithelium might emerge in the chronic phase of the disease, as evidenced by corneal epithelium defects and thinning in the human clinical state of aqueous deficient dry eye. However data presented in this study cannot be extrapolated to the human condition of aqueous deficient disease without confirmatory studies in that entity. Further studies are clearly essential to more precisely define the relationship between the pathways that regulate inflammation, apoptosis, and proliferation in course of desiccating stress and in chronic states of dry eye.
In conclusion, our results demonstrate that desiccating stress in the mouse model of dry eye induces epithelial turnover and increased thickness of the corneal epithelium, similar to what has been observed in the conjunctival epithelium in dry eye, and in other chronic inflammatory epitheliopathies outside of the eye. Changes in epithelial barrier function as a result of these changes, as well as the effect of these hyperplastic epithelial changes on regression in surface ablative refractive surgery when dry eye complicates the postoperative course, are areas ripe for further clinical investigation. At the preclinical level, the mouse CEC appears to be a useful tool to study the regulation of epithelial cell function and cycling at the molecular and cellular levels under stress conditions.
Acknowledgments
The authors would like to thank Dr. Debra Schaumberg for assistance in statistical analysis, Dr. David Sullivan for his helpful comments, and Mr. Don Pottle and Dr. Qiang Zhang for excellent technical assistance.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- Barabino S, Shen L, Chen L, Rashid S, Rolando M, Dana MR. The controlled-environment chamber: a new mouse model of dry eye. Invest Ophthalmol Vis Sci. 2005;46:2766–2771. doi: 10.1167/iovs.04-1326. [DOI] [PubMed] [Google Scholar]
- Beuerman RW, Stern ME. Neurogenic inflammation: a first line of defence for the ocular surface. Ocul Surf. 2005;3:203–206. doi: 10.1016/s1542-0124(12)70256-2. [DOI] [PubMed] [Google Scholar]
- Brignole F, Pisella PJ, De Saint Jean M, Goldschild M, Goguel A, Baudouin C. Flow cytometric analysis of inflammatory markers in KCS: 6-months treatment with topical cyclosporine A. Invest Ophthalmol Vis Sci. 2001;42:90–95. [PubMed] [Google Scholar]
- Brignole F, Pisella PJ, Goldschild M, De Saint Jean M, Goguel A, Baudouin C. Flow cytometric analysis of inflammatory markers in conjunctival epithelial cells of patients with dry eyes. Invest Ophthalmol Vis Sci. 2000;41:1356–1363. [PubMed] [Google Scholar]
- Brown DC, Gatter KC. Ki67 protein: the immaculate deception? Histopathology. 2002;40:2–11. doi: 10.1046/j.1365-2559.2002.01343.x. Review. [DOI] [PubMed] [Google Scholar]
- Dana MR, Zhu SN, Yamada J. Topical modulation of interleukin 1 activity in corneal neovascularization. Cornea. 1998;17:403–409. doi: 10.1097/00003226-199807000-00011. [DOI] [PubMed] [Google Scholar]
- Denda M, Sato J, Masuda Y, Tsuchiya T, Koyama J, Kuramoto M, Elias PM, Feingold KR. Exposure to a dry environment enhances epidermal permeability barrier function. J Invest Dermatol. 1998;111:858–863. doi: 10.1046/j.1523-1747.1998.00333.x. [DOI] [PubMed] [Google Scholar]
- Dursun D, Wang M, Monroy D, Li DQ, Lokeshwar BL, Stern ME, Pflugfelder SC. A mouse model of keratoconjunctivitis sicca. Invest Ophthalmol Vis Sci. 2002;43:632–638. [PubMed] [Google Scholar]
- Erdélyi B, Kraak R, Zhivov A, Guthoff R, Németh J. In vivo confocal laser scanning microscopy of the cornea in dry eye. Graefes Arch Clin Exp Ophthalmol. 2007;245:39–44. doi: 10.1007/s00417-006-0375-6. [DOI] [PubMed] [Google Scholar]
- Gamache DA, Dimitrijevich SD, Weimer LK, Lang LS, Spellman JM, Graff G, Yanni JM. Secretion of proinflammatory cytokines by human conjunctival epithelial cells. Ocul Immunol Inflamm. 1997;5:117–128. doi: 10.3109/09273949709085060. [DOI] [PubMed] [Google Scholar]
- Gillitzer R, Berger R, Mielke V, Muller C, Wolff K, Stingl G. Upper keratinocytes of psoriatic skin lesions express high levels of NAP-1/IL-8 mRNA in situ. J Invest Dermatol. 1994;21:301–307. doi: 10.1111/1523-1747.ep12478128. [DOI] [PubMed] [Google Scholar]
- Hanna C, Bicknell DS, O’Brien JE. Cell turnover in the adult human eye. Arch Ophthalmol. 1961;65:695–698. doi: 10.1001/archopht.1961.01840020697016. [DOI] [PubMed] [Google Scholar]
- Hirai N, Kawasaki S, Tanioka H, Connon CJ, Yamasaki K, Yokoi N, Komuro A, Kinoshita S. Pathological keratinisation in the conjunctival epithelium of Sjogren’s syndrome. Exp Eye Res. 2006;82:371–378. doi: 10.1016/j.exer.2005.07.009. [DOI] [PubMed] [Google Scholar]
- Jones DT, Ji A, Monroy D, Pflugfelder SC. Alteration of ocular surface gene expression in Sjögren’s Syndrome. Adv Exp Med Biol. 1998;438:533–536. doi: 10.1007/978-1-4615-5359-5_75. [DOI] [PubMed] [Google Scholar]
- Kunert KS, Tisdale AS, Gipson IK. Goblet cell numbers and epithelial proliferation in the conjunctiva of Patients with dry eye syndrome treated with cyclosporine. Arch Ophthalmol. 2002;120:330–337. doi: 10.1001/archopht.120.3.330. [DOI] [PubMed] [Google Scholar]
- Lehrer MS, Sun TT, Lavker RM. Strategies of epithelial repair: modulation of stem cell and transit amplifying cell proliferation. J Cell Sci. 1998;111:2867–2875. doi: 10.1242/jcs.111.19.2867. [DOI] [PubMed] [Google Scholar]
- Li DQ, Luo L, Chen Z, Kim HS, Song XJ, Pflugfelder SC. JNK and ERK MAP kinases mediate induction of IL-1beta, TNF-alpha and IL-8 following hyperosmolar stress in human limbal epithelial cells. Exp Eye Res. 2006;82:588–596. doi: 10.1016/j.exer.2005.08.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li DQ, Chen Z, Song XJ, Luo L, Pflugfelder SC. Stimulation of matrix metalloproteinase by hyperosmolarity via a JNK pathway in human corneal epithelial cells. Invest Ophthalmol Vis Sci. 2004;45:4302–4311. doi: 10.1167/iovs.04-0299. [DOI] [PubMed] [Google Scholar]
- Liu Z, Pflugfelder SC. Corneal thickness is reduced in dry eye. Cornea. 1999;18:403–407. doi: 10.1097/00003226-199907000-00002. [DOI] [PubMed] [Google Scholar]
- Pellegrini G, Golisano O, Paterna P, Lambiase A, Bonini S, Rama P, De Luca M. Location and Clonal Analysis of Stem Cells and Their Differentiated Progeny in the Human Ocular Surface. J Cell Biol. 1999;145:769–782. doi: 10.1083/jcb.145.4.769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pflugfelder SC, Huang AJ, Feuer W, Chuchovski PT, Pereira IC, Tseng SC. Conjunctival cytologic features of primary Sjögren’s syndrome. Ophthalmology. 1990;97:985–991. doi: 10.1016/s0161-6420(90)32478-8. [DOI] [PubMed] [Google Scholar]
- Pflugfelder SC, Jones D, Ji Z, Alfonso A, Monroy D. Altered cytokine balance in the tear fluid and conjunctiva of patients with Sjögren’s syndrome keratoconjunctivitis sicca. Curr Eye Res. 1999;19:201–211. doi: 10.1076/ceyr.19.3.201.5309. [DOI] [PubMed] [Google Scholar]
- Sanchis-Gimeno JA, Lleó-Pérez A, Alonso L, Rahhal MS, Martínez-Soriano F. Reduced corneal thickness values in postmenopausal women with dry eye. Cornea. 2005;24:39–44. doi: 10.1097/01.ico.0000138836.45070.0f. [DOI] [PubMed] [Google Scholar]
- Solomon A, Dursun D, Liu Z, Xie Y, Macri A, Pflugfelder SC. Pro- and anti-inflammatory forms of interleukin-1 in the tear fluid and conjunctiva of patients with dry-eye disease. Invest Ophthalmol Vis Sci. 2001;42:2283–2292. [PubMed] [Google Scholar]
- Stern ME, Gao J, Siemasko KF, Beuerman RW, Pflugfelder SC. The role of the lacrimal functional unit in the pathophysiology of dry eye. Exp Eye Research. 2004;78:409–416. doi: 10.1016/j.exer.2003.09.003. Review. [DOI] [PubMed] [Google Scholar]
- Tishler M, Yaron I, Geyer O, Shirazi I, Naftaliev E, Yaron M. Elevated tear interleukin-6 levels in patients with Sjögren syndrome. Ophthalmology. 1998;105:2327–2329. doi: 10.1016/S0161-6420(98)91236-2. [DOI] [PubMed] [Google Scholar]
- Thoft RA, Friend J. The X, Y, Z hypothesis of corneal epithelial maintenance. Invest Ophthalmol Vis Sci. 1983;24:1442–1443. [PubMed] [Google Scholar]
- Tseng SC, Hatchell D, Tierney N, Huang AJ, Sun TT. Expression of specific keratin markers by rabbit corneal, conjunctival, and esophageal epithelia during vitamin A deficiency. J Cell Biol. 1984;99:2279–2286. doi: 10.1083/jcb.99.6.2279. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuominen IS, Konttinen YT, Vesaluoma H, Moilanen JA, Helintö M, Tervo TM. Corneal innervation and morphology in primary Sjogren’s syndrome. Invest Ophthalmol Vis Sci. 2003;44:2545–2549. doi: 10.1167/iovs.02-1260. [DOI] [PubMed] [Google Scholar]
- Turner K, Pflugfelder SC, Ji Z, Feuer WJ, Stern M, Reis BL. Interleukin-6 levels in the cojunctival epithelium of patients with dry eye disease treated with cyclosporine ophthalmic emulsion. Cornea. 2000;19:492–496. doi: 10.1097/00003226-200007000-00018. [DOI] [PubMed] [Google Scholar]
- Villani E, Galimberti D, Viola F, Mapelli C, Ratiglia R. The cornea in Sjögren’s syndrome: an in vivo confocal study. Invest Ophthalmol Vis Sci. 2007;48:2017–2022. doi: 10.1167/iovs.06-1129. [DOI] [PubMed] [Google Scholar]
- Wilson SE, He Y, Weng J, Li Q, McDowall AQ, Vital M, Chwang EL. Epithelial injury induces keratocyte apoptosis: hypothesized role for the interleukin-1 system in the modulation of corneal tissue organization and wound healing. Exp Eye Res. 1996;62:325–337. doi: 10.1006/exer.1996.0038. [DOI] [PubMed] [Google Scholar]
- Wilson SE, Kim WJ. Keratocyte apoptosis: implications on corneal wound healing, tissue organization and disease. Invest Ophthalmol Vis Sci. 1998;39:220–226. [PubMed] [Google Scholar]
- Wilson SE, Liang Q, Kim WJ. Lacrimal gland HGF, KGF, and EGF mRNA levels increase after corneal epithelial wounding. Invest Ophthalmol Vis Sci. 1999;40:2185–2190. [PubMed] [Google Scholar]
- Yeh S, Song XJ, Farley W, Li DQ, Stern ME, Pflugfelder SC. Apoptosis of ocular surface cells in experimentally induced dry eye. Invest Ophthalmo Vis Sci. 2003;44:124–129. doi: 10.1167/iovs.02-0581. [DOI] [PubMed] [Google Scholar]