
Figure 6.
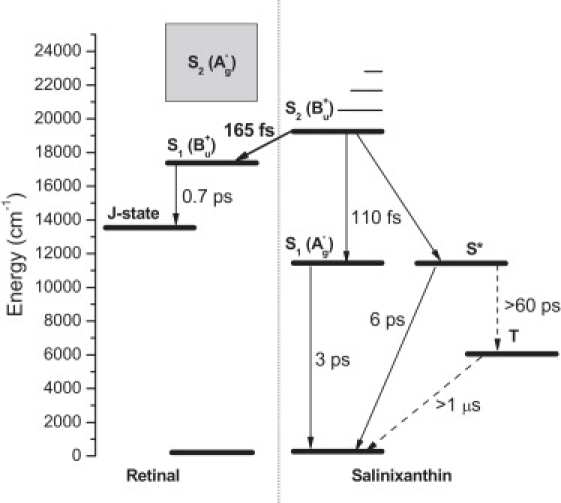
Energy-level diagram depicting relaxation processes that occur after excitation of the S2 state of salinixanthin in xanthorhodopsin. The thick solid arrow represents the energy transfer channel, and the thin solid arrows correspond to the main intramolecular relaxation processes. The dashed lines represent processes involving the salinixanthin triplet state. The processes described in this study are labeled by corresponding time constants. The S∗ to triplet conversion is depicted only schematically because the homofission process first forms a doubly excited triplet that is isoenergetic with the S∗ state, which then relaxes to the lowest triplet state (26,36). The >60 ps time constant obtained from the estimated branching ratio represents the total time for this two-step process. The lifetime of the triplet was not measured in this study and the >1 μs time constant is a rough estimate based on knowledge of carotenoid triplet lifetimes in other systems. The dotted line represents the first step in the xanthorhodopsin photocycle, formation of the J state from the excited retinal chromophore (only the major component is shown, see text for details). Since the energy of the J state is not known, it is only schematically placed above the 11.6 ± 3.4 kcal/mol determined for the K state of bacteriorhodopsin (59). The energy of the forbidden S2 state of retinal in xanthorhodopsin has not been determined. The expected range from studies of bacteriorhodopsin (60) and protonated Schiff bases of retinal (61) is shown in gray. Vibrational bands of the salinixanthin S2 state are depicted to demonstrate that excitation at 490 nm (the 0–1 transition) is somewhat below the lowest expected S2 energy of retinal in xanthorhodopsin. This and the low oscillator strength of the S2 state of retinal would seem to preclude a significant involvement of this state in energy transfer.