

Abstract
Brood parasitism as an alternative female breeding tactic is particularly common in ducks, where hosts often receive eggs laid by parasitic females of the same species and raise their offspring. Herein, we test several aspects of a kin selection explanation for this phenomenon in goldeneye ducks (Bucephala clangula) by using techniques of egg albumen sampling and statistical bandsharing analysis based on resampling. We find that host and primary parasite are indeed often related, with mean r = 0.13, about as high as between first cousins. Relatedness to the host is higher in nests where a parasite lays several eggs than in those where she lays only one. Returning young females parasitize their birth nestmates (social mothers or sisters, which are usually also their genetic mothers and sisters) more often than expected by chance. Such adult relatives are also observed together in the field more often than expected and for longer periods than other females. Relatedness and kin discrimination, which can be achieved by recognition of birth nestmates, therefore play a role in these tactics and probably influence their success.
Alternative breeding tactics, whereby members of a species differ in the ways they compete over reproduction, have been discovered in a variety of animals (1). Most examples concern males, but one widespread such tactic in females is conspecific brood parasitism, which occurs in some insects, fishes, amphibia, and birds. It is particularly common in ducks (2). The parasite lays eggs in the nest of a host female of the same species, which raises the parasitic as well as her own offspring, probably at a cost, because she receives no help from the parasite. The parasite can thereby avoid any extra energy cost and predation risk from incubation and rearing of chicks (2). In goldeneyes and some other waterfowl, host females sometimes prevent a parasite from entering the nest (2).
Female ducks have high return rate to the birth site (natal philopatry; ref. 3). This return rate makes it likely that some local females are closely related (mother–daughter or sisters). Propagation of genes through relatives (4) therefore might favor conspecific brood parasitism among females in ducks (5, 6) and in certain other birds where females nest close to their birth site (7, 8). Relatedness between host and parasite may reduce the fitness cost of being parasitized for the host (5) or even increase its inclusive fitness (6), but the role of female philopatry and relatedness in brood parasitism is still a matter of debate (2, 5–13).
Herein, we test two assumptions of the kin selection hypothesis (5, 6): (i) host and parasite tend to be related in ducks, and (ii) this relationship comes about through female natal philopatry and discrimination of close relatives. We use previously unused techniques (14) of nondestructive albumen sampling and protein fingerprinting of eggs, which greatly help clarify parasitism. Among 902 eggs from 143 female goldeneye ducks (Bucephala clangula) in 71 nests of a wild population, bandsharing (15) is higher in the 42 pairs of host and primary parasite than in random pairs of other females. Additional results show that not only philopatry, but also kin discrimination through recognition of birth nestmates, is important in this alternative female breeding tactic.
Ducks are unusual in forming pairs away from the breeding grounds. The male follows the female to her breeding site, which may be far from his birth place but near hers (3, 16), making the local females likely to be related. To test the idea that hosts and parasites are more closely related than other females, we herein develop protein bandsharing analysis of electrophoretic albumen bands, produced with sensitive isoelectric focusing in immobilized pH gradients (14, 17). Albumen strictly represents the maternal genotype, being secreted by specific cells in the oviduct (18, 19). Sampling the egg albumen is therefore like sampling the female directly. Albumen contains more than a dozen major proteins (18, 20), and genetic polymorphism has been found in many of them; thus, there is considerable genetic variation among females in albumen proteins. Therefore, albumen bands, like DNA fingerprint bands (e.g., refs. 15 and 21), will reflect differences in mean relatedness between categories of individuals.
Materials and Methods
We studied goldeneyes from 1982 to 1997 at Mjörn, a 56-km2 lake 30 km northeast of Göteborg, southwest Sweden, monitoring up to 125 goldeneye nests. Some nests were in natural cavities, but the majority were in dispersed nest boxes in forest trees (on average 120 m from the shore). In a given year, most parasites laid eggs only parasitically, but several parasites in addition produced and incubated a clutch of their own (unpublished work). The present analysis concerns two periods, 1986 and 1988–1991, when albumen was sampled from each of 668 and 234 eggs (59 and 22 nests, respectively), 0–2 days after laying. The samples were stored at 2°C for 1–5 days, and then kept frozen at −70°C until the electrophoresis.
After drilling a small hole in the egg, we take a minute albumen sample (<0.3 ml from a total egg volume of ≈60 ml) with a syringe, seal the hole with cyanoacrylate super glue, and return the egg to the nest; hatchability is preserved (14). The sample is analyzed electrophoretically, producing a rich pattern of albumen protein bands with much variation among females. We use isoelectric focusing in immobilized pH gradients on four different polyacrylamide gels (14). In isoelectric focusing, proteins are separated in an electric field applied over a stable pH gradient in the gel. In contrast with the result of most other electrophoretic methods, each protein therefore comes to rest as a narrow band at its isoelectric point, where the net charge of the protein relative to the gel is zero (17). This aspect greatly facilitates comparison of bands between as well as within gels.
All eggs from a clutch were run on the same gel, and both authors independently categorized the bands of each egg without knowing its identity (14). Representatives of all of the different electromorphs were rerun together on the same gels, with repeats on several gels to allow direct comparison and estimation of bandsharing across all gels. An example of a gel is shown in Fig. 1.
Figure 1.

Electrophoretic gel with albumen band patterns for the 12 eggs of a goldeneye clutch parasitized by three females. There are five host eggs (lanes marked H) and four, two, and one parasite eggs (lanes marked 1, 2, and 3, respectively). Bands that were scored for bandsharing analysis are marked with a short dash in the margin. Several different bands occur in all 902 eggs and can therefore be used as location references; they are marked with a longer dash.
Our previous analyses of eggs from 21 individually marked females, identified by video cameras beneath the nest when alighting for egg laying, showed that they had unique albumen band patterns, fully repeatable between years (14). This variation between females, combined with data on laying dates and sites, makes it possible to distinguish the eggs of different females. Among the 21 video-recorded females, 3–14 unique band patterns (differing from each other in one or more bands) emerged per gel type. Combination of the band patterns from all four gel types used made all 21 females distinguishable, because no two females were identical in all bands (14). Other analyses suggest that the resolution of individual identity is similar to that obtained with a DNA multilocus fingerprinting probe; for mother–daughter or sisters, the likelihood that they will have identical albumen banding patterns was estimated at 0.005–0.01 (14).
Video recordings and nest checks at parasitized nests in 1988–1991 showed that the 12 individually marked hosts, which later incubated the clutch, initiated egg laying and produced the highest number of eggs in the nest in all cases but one (ref. 14 and M.Å., unpublished work). For 1986, we therefore assume that the female laying the highest number of eggs was the host. A Monte Carlo simulation based on the numbers of eggs laid by the video-recorded females shows that the risk of mistaking the primary parasite (with more eggs than any other parasite in the nest) for the host is low, about 5.1%. We may have confused host and primary parasite in 1 or 2 of the 29 parasitized nests from 1986 analyzed below; however, this confusion should not influence our estimate of relatedness, because the two females making up the host–parasite pair are still the same. For other than the primary parasite, the risk of confusion with the host is negligibly small, because the other parasites laid so few eggs (unpublished work).
There is no evidence from this or other studies (2) that laying female ducks eliminate eggs not their own.
The present isoelectric focusing method results in a similar number of individually repeatable bands as does multilocus DNA fingerprinting. Albumen is of strictly maternal origin, with band variation that reflects genotypic differences (see above). Therefore, bandsharing analysis, as with DNA fingerprint studies (e.g., 8, 15, 21–24), can be applied to albumen bands for estimating differences in mean relatedness between categories of individuals. For each egg, presence/absence of each band was recorded in a matrix. In total, 98 different bands were found. There was no indication that two females laying in the same nest had identical band patterns (such as two eggs with the same band pattern being laid on the same day in the nest). Removal of 39 bands, each identical in occurrence to another band, resulted in 59 nonredundant bands. The mean number of bands per egg was 11.5 (SD = 1.43). Relatedness is seriously underestimated if common bands are included, and bands with a population frequency >50% should be omitted (15), which reduced the number of bands used to 50.
The bandsharing analysis permits a test of whether host and parasite are closer relatives than other females. We use the bandsharing similarity index Sxy = 2Nxy/(Nx + Ny), where Nxy is the number of bands shared by individuals x and y, and Nx and Ny are the total numbers of bands present in individuals x and y, respectively (e.g., refs. 21–23).
In addition to estimating host–parasite relatedness from eggs, we also tested for possible nestmate recognition and kin discrimination by analyzing the occurrence of marked individuals together on the lake. The hypothesis is that birth nestmates, i.e., the ducklings in the brood and the host female that raises them, may recognize and associate with each other later in life.
Although some of the ducklings are not siblings if the nest has been parasitized, they and the host female will on average be much more closely related than random females. Individual recognition of birth nestmates therefore permits (imperfect) kin discrimination. With the present degree of parasitism, on average about one-third of the chicks are not the genetic offspring of the host, and the mean relatedness between host and chicks is not 0.5 but about 0.4 (see below).
In total, 139 adult females were marked with a unique combination of a numbered steel band and up to three color bands, and 1,172 chicks were marked in the nest with a numbered aluminum tag in each wing. While visiting the study area for trapping females or monitoring broods, we recorded all observations of marked females on the lake. We focused on two frequently visited subareas, each of a size roughly similar to the home range of females (which have an average maximum diameter of 0.52 km; M.Å., unpublished work). We noted whether two marked females were associated: staying or moving together while feeding, preening, or resting, usually less than 5 m from each other (most of the time much closer) and being clearly separated from others during the 15–60 min of observation required to determine the identity of a female. Some pairs of marked females were seen repeatedly over periods of up to several weeks. The time between first and last observation of a pair is used as an estimate of the permanence of their association.
Depending on where they were observed most often, marked females were assigned to one of the two areas (see Table 2). Several of these females were known to be birth nestmates. In the list of N marked females thus obtained for an area, we counted the number of pairs of nestmates, Nr, that could be formed. The total number of different pairs that can be formed from N individuals is NP = N(N − 1)/2. We then counted the number, nm, of different pairs of marked females that had actually been observed together at least once and noted how many of these pairs consisted of nestmates, nr. Under the null hypothesis of no kin discrimination, each possible pair of marked nestmates (in total Nr pairs) has the same chance of occurrence as each possible pair of marked nonnestmates (Np − Nr pairs) in the area.
Table 2.
Individually marked females observed on the water, alone or in pairs, with known birth nestmates or with other marked females
| Area and year | N (number of marked females observed) | Np [number of possible pairs N(N − 1)/2] | Nr (maximum number of nestmate pairs) | nm (number of pairs of marked females observed) | nr (number of nestmate pairs observed) |
|---|---|---|---|---|---|
| U 1986 | 5 | 10 | 3 | 1 | 1 |
| U 1987 | 13 | 78 | 4 | 6 | 1 |
| J 1989 | 15 | 105 | 2 | 4 | 1 |
| J 1990 | 11 | 55 | 4 | 1 | 0 |
| U 1990 | 10 | 45 | 1 | 3 | 1 |
Nr is the number of pairs of birth nestmates that can be formed among the observed marked individuals for the relevant area and year. There may be additional birth nestmate pairs not known to us among the females. Because known nestmates are overrepresented among the observed pairs, unknown nestmates are also expected to be overrepresented (compared to unrelated birds) among the observed pairs. The fact that unknown nestmates will not be recorded as such therefore tends to make the test conservative).
A Monte Carlo test (25, 26) of the null hypothesis was done as follows. Drawing nm pairs at random from the altogether Np possible pairs, we counted the number of pairs drawn that consisted of nestmates. This count was done for each combination of area and year for which there were at least two nestmates among the marked birds observed. The total number of nestmate pairs drawn was counted, and the result was compared with the actual number of pairs of nestmates (Σnr) observed in the combined sample for all areas and years (see Table 2). The process was repeated 105 times. The proportion of cases in which at least as many pairs of nestmates were drawn as the number actually observed was calculated, estimating the probability of obtaining the observed or a more extreme outcome under the null hypothesis of no nestmate recognition.
Results and Discussion
Host–Parasite Relatedness.
If the ratio of gain to loss in inclusive fitness is greater than that for producing an additional own offspring, hosts may gain inclusive fitness by preferentially accepting close relatives as parasites, eliminating costs of parental care for the relative (5, 6). Whatever the costs of being parasitized, which are as yet not sufficiently known, the costs for the host should be more acceptable with increasing relatedness to the parasite. A prediction is therefore that the primary parasite (laying most parasitic eggs) should be related to the host. Lack of relatedness is evidence against the kin selection hypothesis.
The two study periods are first treated separately, then combined. Breeding in 1986 progressed to incubation in 29 parasitized nests (26 of which were parasitized by more than one female), with a mean bandsharing value of 0.288 between host and primary parasite. As the control group, we used the 42 other females sampled in the same area, for which the number of different pairs that can be formed is 42C2 = 42 × 41/2 = 861. We calculated bandsharing for each of these possible pairs and used their mean as the estimate of bandsharing in random pairs of females not involved in host–parasite relationship with each other (15). This mean is 0.163, i.e., 0.125 less than that for host–parasite pairs. The bandsharing distribution is shown in Fig. 2.
Figure 2.

Distribution of bandsharing values in the 29 pairs of host and primary parasite in 1986 and in the 861 different random pairs that can be drawn among the 42 other females not involved in host–parasite relationship with each other. Bandsharing values are significantly higher for the host–parasite pairs than for the randomly formed pairs (see Host–Parasite Relatedness).
The second sample, from 1988 to 1991, is smaller but shows a similar pattern. There were 13 pairs of host and primary parasite, with a mean bandsharing of 0.257, and 24 control females, with a mean bandsharing of 0.180 for the 24C2 = 276 different pairs that they can form.
We tested bandsharing (discussed in refs. 22 and 23) with a bootstrap test (25, 26) tailored to the present situation. It estimates the probability that a difference at least as large as that observed between the host–parasite and the control group will arise by chance, under the null hypothesis that both groups represent the same statistical population. The combined test for the two periods results in P = 0.0026, refuting the null hypothesis (see Appendix). Host and primary parasite clearly tend to be more closely related than the females in the control group.
How Closely Related Are Hosts and Primary Parasites?
Using a regression approach to relatedness, Reeve et al. (15) derived an estimate of relatedness from DNA fingerprints as
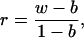 |
1 |
where w is the mean bandsharing proportion within groups (such as families), and b is the mean bandsharing proportion between groups. This relatedness estimator does not require knowledge of allelic state at the loci involved, nor does it require a standard curve relating bandsharing similarity to known relatedness levels (15). It can therefore be used with the present albumen data, which reflect maternal genotype (see above) and relatedness between females. Our results contain one pair of marked females known to be genetic mother and daughter (with true relatedness 0.5), because the nest was not parasitized. Their estimated relatedness (Eq. 1) is 0.594. Adding the two other similar marked pairs where >90% of the eggs in the nest were laid by the host (making it likely that the females in the observed pairs were genetic mother–daughter), the three cases together give an estimated mean r = 0.49. This value supports the validity of the present approach for estimating mean relatedness.
In this study, w is the mean bandsharing in host–primary parasite pairs (0.288 in 1986), and b is the mean bandsharing in pairs of randomly drawn control females that are not host–parasite pairs (0.163). For the 1986 data, Eq. 1 then gives an estimated relatedness of 0.125/0.837 ≈0.149. For the 1988–1991 data, the estimate is r = 0.093. The weighted mean for both periods is r = 0.132, slightly higher than for first cousins (r = 0.125). This result does not imply that host–parasite pairs are usually cousins. More likely, parasites range from being close relatives (mother–daughter or sisters; see below) to being unrelated to the host. Primary parasites produced about two-thirds of the parasitic eggs (unpublished work). If other than primary parasites are also included, the weighted mean relatedness between host and parasite as expected becomes lower; it is then estimated at 0.100.
The estimator in Eq. 1 tends to underestimate true relatedness (15). For this and other reasons, such as the possibility of random misscoring of bands, our estimates are likely to be lower than the true relatedness.
Kin Discrimination and Favoritism.
What is the reason for higher relatedness in host–parasite pairs? One possibility is simply that high return rate of females to the birth site makes close relatives likely to use the same nest site, although they do not recognize each other. Another possibility is that birth nestmates, which are usually close relatives (see below), in addition recognize and treat each other favorably (6, 27, 28). For example, birth nestmates may associate with each other, and hosts may be more tolerant of their birth nestmates than of other parasites. Preferential treatment of adult relatives has rarely been looked for in ducks, but it has been found in female canvasback ducks (Aythya valisineria: ref. 3). Goldeneye broods usually remain together with the host female for at least 5–6 weeks after hatching (29); thus, the chicks and the host may be able to recognize and associate with each other in later years.
The possibility that individual recognition and kin discrimination play a role can be tested in at least two ways.
(i) A first prediction is that individual recognition between birth nestmates increases a parasite's ease of access to the nest. A female that lays parasitically in several nests therefore should usually be more closely related to the host for which she lays the most eggs than to the host for which she lays the fewest eggs. This idea can be tested for 20 parasites, each of which laid in several nests. As predicted, host–parasite bandsharing was higher (0.272) in nests where the parasite laid the highest number of eggs (on average 3.0) than where she laid only one egg (0.139; P = 0.018, one-tailed exact permutation test). The mean distance between the two nests was 0.94 km. (Because this test is done within the group of host–parasite pairs, it is statistically independent from the previous test of relatedness, which was done between host–parasite pairs and other females.)
(ii) A second prediction is that recognition and relatedness between host and parasite make their interactions peaceful and reduce the risk that the clutch is abandoned or subject to predation. The risk should be higher if there are conspicuous conflicts with unrelated parasites that try to get access to the nest (there is evidence of conflicts in goldeneyes; see ref. 30). Although not significant at the α = 0.05 level, there was a trend in the predicted direction: bandsharing between host and primary parasite in both periods tended to be higher among parasitized nests that hatched successfully (1986: 0.347, n = 16; 1988–1991: 0.304, n = 8) than among nests that were abandoned or preyed on during the incubation period (1986: 0.216, n = 13; 1988–1991: 0.181, n = 5). One-tailed resampled randomization tests for the two periods gave P = 0.088 and P = 0.163. Combining these results with Fisher's method (31) gives P = 0.075.
If the previous analysis is expanded to include all parasites laying in the nests, their weighted mean bandsharing with the host is, for successful clutches: 0.292 (1986) and 0.297 (1988–1991), and for other clutches: 0.202 (1986) and 0.184 (1988–1991).
If, as seems likely, host females are not able to distinguish their own genetic offspring from other chicks in the brood, the average relatedness between host and chicks in the brood may be of interest in considerations of inclusive fitness (32, 33). For all of the 35 hatched clutches studied herein, the weighted mean relatedness between the host and all chicks in the nest is estimated (from our protein bandsharing data, with Eq. 1) at 0.399 ≈ 0.4, as compared with 0.5 for strictly genetic mother–offspring broods. In these calculations, we assumed that the (unknown) fathers of the chicks were unrelated to the host female.
Philopatry Effects Only?
Can the previous results on host–parasite relatedness be explained simply as consequences of philopatric nest choice not involving individual recognition? Detailed analysis of observed shifts of nest site for marked females suggests that this explanation is unlikely. Even successfully breeding females often shift nest site by over 1 km between years, and returning young females usually breed much farther from the birth nest (on average 0.82 km; n = 16) than required for philopatry to be a sufficient explanation (also see ref. 10). Although our observations of parasitism by marked females are few (n = 6), it has been recorded up to 1.6 km from the birth nest.
For a returning young female, her only substantial chance of parasitizing a close relative through philopatry alone (without individual recognition) is to lay parasitically in the birth nest (Table 1). The earliest females start breeding when 2 years old (mean = 3.1 years; M.Å., unpublished work). For a philopatric 2- to 5-year-old female, estimates from our nesting records show that the probability that one of her birth nestmates is the host in her birth nest is on average only 0.19, if the nest is inhabited (Table 1). (Corresponding estimates of mothers' chances of parasitizing daughters purely through philopatry show that these probabilities are an order of magnitude lower, i.e., negligible.)
Table 1.
Probability that a female who parasitizes an active nest will by chance have a birth nestmate as host, depending on the age of the female and the distance of the parasitized nest from her birth nest
| Nest distance | Age of female in years (no. of females)
|
|||||
|---|---|---|---|---|---|---|
| 2 (24) | 3 (18) | 4 (10) | 5 (6) | 6 (3) | Mean | |
| 0 | 0.083 | 0.111 | 0.400 | 0.167 | 0 | 0.19 |
| 1–2 | 0.105 | 0.083 | 0.125 | 0 | 0 | 0.078 |
| 3–6 | 0.042 | 0.028 | 0 | 0 | 0 | 0.0175 |
| 7–20 | 0.0030 | 0.016 | 0.0079 | 0 | 0 | 0.0067 |
Nest distance is counted in number of active nests from the female's birth nest (0). The results are based on all females returning (and present at the age specified) of 1,172 chicks wing-tagged as nestlings. For instance, at age 3, 2 of 18 females present (P = 0.111) would have parasitized a birth nestmate (social mother or sister) by laying eggs in the birth nest (0). Means are for ages 2–5.
Table 1 therefore shows that females that parasitize their birth nests are expected to parasitize a birth nestmate in about 20% of the cases. Do returning daughters actually show a strong preference for the birth nest? Two kinds of evidence suggest that they do not do so. First, returning females wing-tagged as chicks do not usually breed in the birth nest or its nearest neighbors (also see ref. 10). Among eight such females that began nesting in a year when the birth nest was empty, only one nested there. The seven others nested on average 1.1 km (20 nest boxes) away.
Other evidence is provided by our video recordings of active nest boxes (M.Å., unpublished work), which in three cases made it possible to determine whether returning young females parasitized their birth nests. None of them did so. One of these females instead parasitized her mother, who was breeding in the second inhabited nest from the daughter's birth nest, 400 m away. The second female parasitized her sister, in the 20th inhabited nest from the birth nest, 1.6 km away. The third female also parasitized a nest 400 m from her birth nest, but the host could not be determined, because that nest failed before incubation began; it is therefore not used in the following calculation.
Assuming as a null hypothesis that there is no individual recognition, what is the likelihood that the two parasitizing females each will parasitize a birth nestmate purely by chance? In both cases, the parasite was 3 years old. In Table 1, the estimated probability of a birth nestmate being the host in nest no. 2 from the birth nest in year 3 is 0.083. The corresponding probability for nest no. 20 is 0.016. The product of these two independent probabilities is 0.0013. Owing to limited sample sizes, these yearly estimates are uncertain. If, therefore, the means for years 2–5 (Table 1) are used instead, the result is 0.078 × 0.0067 = 0.00052. Calculated either way, the results strongly suggest that the null hypothesis should be rejected, instead corroborating the alternative that individual recognition is involved.
What else than individual recognition and consequential kin discrimination might explain a tendency for higher host-parasite relatedness in successful rather than in failing nests (test ii above)? One possibility is that some females nest successfully during several years in the same nest, producing many offspring that may return to the same nest and parasitize it. Even without recognition of birth nestmates, such a mechanism might produce the result in test ii. Two aspects make this interpretation unlikely, however. First, recruiting females lay eggs only rarely in their birth nest (see above). Second, host females in the successful nests in test ii did not continue to use the same nest longer than hosts in unsuccessful nests. For 1986 females, successful females nested in the same box on average 1.31 years, and unsuccessful females 1.23 years (P = 0.69, Wilcoxon test). For 1988–1991 females, the corresponding figures are 2.50 vs. 2.80 years (P = 0.82). Therefore, there is no evidence that the observed difference arose through this alternative mechanism.
For these reasons, the natal and breeding philopatry of females is not precise and consistent enough to explain all our results on host–parasite relatedness; kin discrimination is also required. This conclusion differs from that for moorhens, where host–parasite relatedness is explained fully by philopatry (8). An important difference is that philopatry was much more precise in the moorhen population, where 82% of the locally recruited breeders nested within two territories of their parents. Relatives were therefore likely to be parasitized, simply because brood parasitic moorhens almost exclusively selected neighbors as hosts (8). In the present goldeneye population on the other hand, only 13% (2 of 16) of the recruiting wing-tagged females nested in the birth nest or one of its two nearest neighbor nests (M.Å., unpublished work). Philopatry in goldeneyes therefore is much less precise than in the moorhens, which showed no sign of kin discrimination (8). Its likely occurrence in goldeneyes (and some other ducks; ref. 3) may be related to parasitism being much more common and important in this species (like in many other ducks; ref. 2). In the moorhen, only 4–9% of the eggs were parasitic (8), compared with 36% in the goldeneye (14).
Parasitism by unrelated females cannot be prevented entirely, because laying goldeneye females often spend much time far from the nest (mean distance nest − feeding territory ≈0.5 km). Video recordings showed that hosts during the laying period usually visit the nest only every other day, which may allow parasites easy access in days of host absence. When present, however, the host in 14 of 22 video-recorded cases rose to the entrance hole and prevented the approaching female from entering (M.Å., unpublished work). In contrast, only one time in nine did a parasite do so. As expected if there is discrimination, hosts therefore reject some but not all visits by other females (also see ref. 2). The females not rejected entered the nest while the host was present, and in four of these cases, the parasite laid an egg without any apparent aggression occurring (M.Å., unpublished work). Relatedness is, however, not known in most cases, and a larger sample is needed for meaningful testing of host behavior in relation to parasite relatedness.
Associations on the Lake Between Birth Nestmates.
We do not know how parasites find the nests of related hosts. Individual recognition and association on the lake with females from the birth nest, combined with joint nest visits, is one possibility. Compared with the likelihood of randomly drawing pairs of females that are birth nestmates from the pool of all marked females present, nestmate pairs are overrepresented in our field observations of marked pairs (Table 2, nr). Using these data in a Monte Carlo test of the null hypothesis of no nestmate or kin discrimination (see Materials and Methods), we find that the probability of observing as many as Σnr = 4 or more different pairs of nestmates is low, P < 0.0038. The null hypothesis is therefore refuted.
The sample areas are small, about 0.7 km in maximum diameter, of a size similar to the home ranges over which these females were observed (see below). It is therefore unlikely that limited individual movements in combination with philopatry alone can explain why pairs of nestmates were overrepresented in the observations. Most individuals moved over the entire sample area, in many cases also outside of it. In particular, there was no evidence that females occurring in pairs with nestmates had smaller home ranges than other females. Based on the mean distance between places where the female was seen, the home ranges (±SEM) for the marked females of Table 2 that were observed at least three times are, for females seen in pairs with nestmates, 0.62 ± 0.12 km (n = 4); for females seen in pairs with other marked females, 0.48 ± 0.10 km (n = 7); and for females not seen in pairs with other marked females, 0.45 ± 0.15 km (n = 5). The results suggest that birth nestmates occur more often together than expected by chance, because they recognize and tend to favor company with each other. As birth nestmates are usually genetically related (mother–daughter or sisters, see above), association between them leads to association between relatives.
The duration of associations in pairs of marked females strengthens this conclusion. We observed three pairs of marked birth nestmates and five other pairs more than once. The kin discrimination hypothesis predicts that nestmate pairs will have longer associations than other pairs; the null hypothesis predicts no difference. The time in days between first and last observation was, for the three nestmate pairs, 23 (mother–daughter), 20, and 9 (sisters), and for the five other pairs 8, 4, 2, 2, and 2, a significant difference (P = 0.018, one-tailed exact randomization test). If we also represent the pairs observed only once with an association value of 1 day, the difference between the two groups becomes even more pronounced (P = 0.012). As predicted by the kin discrimination hypothesis, these results therefore suggest that birth nestmates, which are usually genetically related, tend to form longer associations than other females. Family members in several other diving ducks may also be found together long after the young are fledged, even on the wintering grounds (34) or on the breeding grounds in subsequent years (35).
Theoretical models of relatedness in conspecific brood parasitism have produced contrasting results. One model predicts host–parasite relatedness (6); the other predicts that parasitism is less likely as the relatedness between host and parasite increases (12). However, some assumptions in this latter model, e.g., that the host clutch is reduced by the exact number of parasitic eggs added, seem unrealistic in relation to ducks, and a modified version predicts that relatedness and kin discrimination can facilitate the evolution of conspecific brood parasitism (M.A., unpublished work).
Two recent empirical studies besides the present one have used molecular methods to estimate relatedness in brood parasites with female natal philopatry (8, 13). All three studies, two of birds and one of a lacebug insect, find host–parasite relatedness; thus, it may have some generality. (In species with well developed cooperative breeding, the picture may be different; see refs. 7 and 12.) There is also other recent evidence for the importance of relatedness in social situations where it has not often been looked for, in lekking as well as territorial male grouse and in lekking peacocks (36–38). Relatedness may therefore be even more important in social evolution than expected.
Conclusions
Genetic relatedness and kin discrimination, which may be achieved indirectly by a combination of natal philopatry and recognition of birth nestmates (or possibly by more sophisticated kin recognition), play a role in conspecific brood parasitism and social behavior in goldeneyes, as previously suggested on theoretical grounds (5, 6). Our preliminary results also suggest that relatedness affects the success of the female alternative reproductive tactics involved, and therefore probably also their frequency. The present findings lead to important questions for research at the interface of reproductive behavior and kin selection, concerning the means and precision by which females discriminate and favor relatives, e.g., at the nest (27, 28); the costs, benefits, and inclusive fitness of the tactics involved (1, 4, 39); and the factors that determine whether a female will be host, parasite, or both (2, 6). The possibility that relatedness plays a role in the evolution of alternative reproductive tactics deserves more attention. Several recent results suggest that relatedness and kin discrimination have even wider importance in animal sociality than anticipated by most researchers except, of course, W. D. Hamilton (4, 27).
Acknowledgments
We thank J. Bohlin and U. Unger for help in the field; D. Queller and K. Reeve for helpful discussion; S. Holm for statistical advice; S. Andersson, D. Hasselquist, A. Johnsen, M. Olsson, J. Reynolds, B. Sheldon, T. Slagsvold, H. Smith, the referees, and the editor for constructive suggestions; the Swedish Natural Sciences Research Council for grant support; and E. Bonabeau and the Santa Fe Institute for inviting M.A. to the 1998 workshop on the evolution of social behavior.
Appendix
For 1986, our approximate bootstrap test (25, 26) of bandsharing involved sampling with replacement 29 values from the 29 observed host–parasite bandsharing values, calculating the mean of the sample, determining whether it is smaller than the mean of the 861 random control pairs (0.163), and scoring the result if it is smaller. The process was repeated 106 times, and the total score was divided by this number to estimate the probability of obtaining an equally or more extreme result if the null hypothesis were true. The (one-tailed) outcome for 1986 was P = 0.00267. The corresponding test for the second period, 1988–1991, was P = 0.1059. These two results from different tests of the same hypothesis can be combined as suggested by Fisher (31), which leads to a total probability of P = 0.0026 (χ2 test; df = 4; −2Σ ln P = 16.34), refuting the null hypothesis.
We also tested the null hypothesis by using a highly conservative randomization test, which resulted in P = 0.045. Our results therefore reject the null hypothesis and corroborate the alternative that host and primary parasite are on average more closely related than females not involved in a host–parasite relationship with each other.
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
See commentary on page 12942.
Article published online before print: Proc. Natl. Acad. Sci. USA, 10.1073/pnas.220137897.
Article and publication date are at www.pnas.org/cgi/doi/10.1073/pnas.220137897
References
- 1.Gross M R. Trends Ecol Evol. 1996;11:92–98. doi: 10.1016/0169-5347(96)81050-0. [DOI] [PubMed] [Google Scholar]
- 2.Sayler R D. In: Ecology and Management of Breeding Waterfowl. Batt B D J, Afton A D, Anderson M G, Ankney C D, Johnson D H, Kadlec J A, Krapu G L, editors. Minneapolis: Univ. of Minnesota Press; 1992. pp. 290–322. [Google Scholar]
- 3.Anderson M G, Rhymer J M, Rohwer F C. In: Ecology and Management of Breeding Waterfowl. Batt B D J, Afton A D, Anderson M G, Ankney C D, Johnson D H, Kadlec J A, Krapu G L, editors. Minneapolis: Univ. of Minnesota Press; 1992. pp. 365–395. [Google Scholar]
- 4.Hamilton W D. J Theor Biol. 1964;7:1–52. doi: 10.1016/0022-5193(64)90038-4. [DOI] [PubMed] [Google Scholar]
- 5.Andersson M, Eriksson M O G. Am Nat. 1982;120:1–16. [Google Scholar]
- 6.Andersson M. In: Producers and Scroungers. Barnard C J, editor. London: Croom-Helm; 1984. pp. 195–228. [Google Scholar]
- 7.Emlen S T, Wrege P H. Ethology. 1986;71:2–29. [Google Scholar]
- 8.McRae S T, Burke T. Behav Ecol Sociobiol. 1996;38:115–129. [Google Scholar]
- 9.Eadie J M, Kehoe F P, Nudds T D. Can J Zool. 1988;66:1709–1721. [Google Scholar]
- 10.Pöysä H, Runko P, Ruusila V. J Avian Biol. 1997;28:63–67. [Google Scholar]
- 11.Weatherhead P J. J Avian Biol. 1998;29:321–322. [Google Scholar]
- 12.Zink A G. Am Nat. 2000;155:395–405. doi: 10.1086/303325. [DOI] [PubMed] [Google Scholar]
- 13.Loeb M L G, Diener L M, Pfennig D W. Anim Behav. 2000;59:379–383. doi: 10.1006/anbe.1999.1328. [DOI] [PubMed] [Google Scholar]
- 14.Andersson, M. & Åhlund, M. (2001) Ecology, in press.
- 15.Reeve H K, Westneat D F, Queller D C. Mol Ecol. 1992;1:223–232. [Google Scholar]
- 16.Greenwood P J, Harvey P H. Annu Rev Ecol Syst. 1982;13:1–21. [Google Scholar]
- 17.Righetti P G. Immobilized pH Gradients: Theory and Methodology. Amsterdam: Elsevier; 1990. [Google Scholar]
- 18.White H B I. In: Egg Incubation: Its Effects on Embryonic Development in Birds and Reptiles. Ferguson M W J, Deeming D C, editors. Cambridge, U.K.: Cambridge Univ. Press; 1991. pp. 1–15. [Google Scholar]
- 19.Palmer B D, Guillette L J., Jr . In: Egg Incubation: Its Effects on Embryonic Development in Birds and Reptiles. Ferguson M W J, Deeming D C, editors. Cambridge, U.K.: Cambridge Univ. Press; 1991. pp. 29–46. [Google Scholar]
- 20.Burley R W, Vadhera D V. The Avian Egg: Chemistry and Biology. New York: Wiley; 1989. [Google Scholar]
- 21.Wetton J H, Carter R E, Parkin D T, Walters D. Nature (London) 1987;327:147–149. doi: 10.1038/327147a0. [DOI] [PubMed] [Google Scholar]
- 22.Lynch M. Mol Biol Evol. 1990;7:478–484. doi: 10.1093/oxfordjournals.molbev.a040620. [DOI] [PubMed] [Google Scholar]
- 23.Danforth B B N, Freeman-Gallant C R. Mol Ecol. 1996;5:221–228. doi: 10.1111/j.1365-294x.1996.tb00309.x. [DOI] [PubMed] [Google Scholar]
- 24.Birkhead T R, Burke T, Zann R, Hunter F M, Krupa A P. Behav Ecol Sociobiol. 1990;27:315–324. [Google Scholar]
- 25.Davison A C, Hinkley D V. Bootstrap Methods and Their Application. Cambridge, U.K.: Cambridge Univ. Press; 1997. [Google Scholar]
- 26.Manly F J. Randomization, Bootstrap and Monte Carlo Methods in Biology. 2nd Ed. London: Chapman & Hall; 1997. [Google Scholar]
- 27.Hamilton W D. In: Kin Recognition in Animals. Fletcher D J C, Michener C D, editors. Chicester, U.K.: Wiley; 1987. pp. 417–438. [Google Scholar]
- 28.Sherman P W, Reeve H K, Pfennig D W. In: Behavioural Ecology. 4th Ed. Krebs J R, Davies N B, editors. Oxford: Blackwell; 1997. pp. 69–96. [Google Scholar]
- 29.Eadie J M, Mallory M L, Lumsden H G. In: The Birds of North America. Poole A, Gill F, editors. Philadelphia: Acad. Nat. Sci.; 1995. , No. 170, pp. 1–32. [Google Scholar]
- 30.Grenquist P. Proc Int Ornithol Congr. 1963;13:685–689. [Google Scholar]
- 31.Sokal R R, Rohlf F J. Biometry. 3rd Ed. New York: Freeman; 1995. [Google Scholar]
- 32.West Eberhard M J. Q Rev Biol. 1975;50:1–33. [Google Scholar]
- 33.Brown J L. Helping and Communal Breeding in Birds. Princeton, NJ: Princeton Univ. Press; 1987. [Google Scholar]
- 34.Cooke F, Robertson G J, Smith C M. Condor. 2000;102:137–144. [Google Scholar]
- 35.Anderson M G. Ornithol Monogr. 1985;37:57–67. [Google Scholar]
- 36.Höglund J, Alatalo R V, Lundberg A, Rintamäki P T, Lindell J. Proc R Soc London Ser B. 1999;266:813–816. [Google Scholar]
- 37.Pirtney S B, MacColl A D C, Lambin X, Moss R, Dallas J F. Biol J Linn Soc. 1999;68:317–331. [Google Scholar]
- 38.Petrie M, Krupa A, Burke T. Nature (London) 1999;401:155–157. [Google Scholar]
- 39.Lyon B E. Nature (London) 1998;392:380–383. [Google Scholar]