
Figure 5.
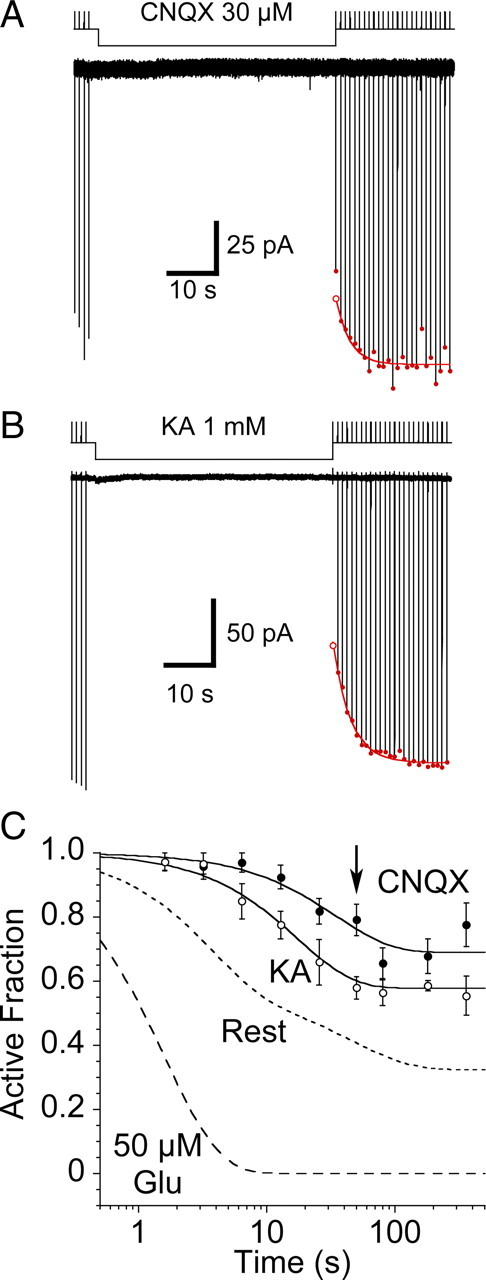
Disulfide trapping of the S729C mutant in the presence of CNQX or kainate. A, Coapplication of 30 μm CNQX slowed the onset of, and reduced the extent of trapping by 10 μm CuPhen; the first response following the end of the antagonist application was reduced in amplitude because CNQX remained bound during the rise of the current evoked by the test application of 10 mm glutamate. B, Trapping in the presence of a saturating concentration of the weak partial agonist kainate also slowed the onset, and reduced the extent of trapping by 10 μm CuPhen, but to a lesser extent than CNQX; the first response following the end of the partial agonist application was reduced in amplitude because kainate remained bound during the rise of the current evoked by the test application of 10 mm glutamate. C, Data summarizing trapping in the presence of 1 mm kainate (n = 6) and 30 μm CNQX (n = 4–10 patches per data point), compared with trapping at rest and in the presence of 50 μm glutamate (dashed lines); the arrow indicates the trapping interval for the representative examples shown in A and B. The rate of trapping of the S729C mutant in the presence of kainate (0.06 ± 0.01 s–1) is slower than for receptors at rest, even though kainate causes weak desensitization (supplemental Fig. 2, available at www.jneurosci.org as supplemental material); trapping by CNQX is slower still (0.03 ± 0.01 s–1); note also the inverse correlation between rate and extent of trapping.