

Abstract
To determine if lower levels of hepatitis C virus (HCV)-specific neutralizing antibodies (nAb) are associated with an increased risk of mother-to-child transmission (MTCT) of HCV, anti-HCV nAb titers were assessed in 63 mothers co-infected with HCV and human immunodeficiency virus type 1 (HIV). Among the mothers, 16 transmitted HCV to their infant but no difference was detected between the ability of maternal plasma from transmitters and non-transmitters to neutralize heterologous HCV pseudoparticles (median nAb titer 1:125 vs. 1:100, P=0.23). In the setting of HIV/HCV co-infection, we found no evidence that anti-HCV nAbs are associated with prevention of MTCT of HCV.
Keywords: HCV, HIV, mother-to-infant transmission, perinatal transmission, hepatitis C virus, neutralizing antibody, HCVpp, MTCT
Hepatitis C virus (HCV) is transmitted to a minority of children born to HCV-infected mothers [1-3]. The risk of mother-to-child transmission (MTCT) of HCV is increased if the woman also is infected with the human immunodeficiency virus type 1 (HIV) [3]. HIV increases the abundance of HCV RNA detected in plasma (viral load), and a higher maternal HCV viral load has been associated with increased MTCT of HCV [2]. However, how HIV infection enhances HCV transmission, whether or when HCV is transmitted across the intact placenta, and why the majority of children born to HCV-infected women remain uninfected are unknown.
In the months after acute HCV infection, adaptive immune responses are detectable that are sufficient to clear specific virus variants from blood (and drive evolution) but inadequate to eradicate infection [4,5]. Recent development of HCV pseudoparticles (HCVpp) has allowed characterization of neutralizing antibodies (nAbs) specific for HCV envelope glycoproteins [6,7]. We, and others, have found marked inter-person differences in the titers and kinetics of HCV nAbs [4,6,8-11]. We also found that HIV infection reduces HCV specific antibody titers [12]. In the present investigation, we tested the hypothesis that low maternal HCV nAb titers among HIV and HCV co-infected mothers is associated with MTCT of HCV.
Subjects, materials, and methods
Participants
Study subjects were participants in the Women and Infants Transmission Study (WITS) [2]. From 1989-2005, HIV-infected women were enrolled at institutions in New York City, Chicago, Boston, Houston, and San Juan, Puerto Rico. Women could enroll at any time during pregnancy and were examined at entry and at structured times during pregnancy, at delivery, and thereafter. At each visit, a physical examination was performed and a medical history was obtained; venous blood was collected and components were stored at -70°C. At delivery, or shortly after, and at one, two, four, six, nine, 12, 15, 18, 24, 30, and 36 months of age, infants were examined and venous blood was collected. According to protocol, children were not breast fed. The study was approved by the institutional review board at each participating center. Written informed consent was obtained from each participating woman.
MTCT of HCV was assessed in HCV antibody-positive mother-infant pairs who had at least six months of infant follow-up. Previously, HCV infection was documented in 12 infants with follow-up until September 1994 [13]. Testing of additional mothers and infants enrolled through 2005 revealed another five HCV-infected infants.
To test the hypothesis that lower titer HCV nAbs increased the risk of infant HCV infection, a nested case control study was conducted among mothers of HCV-infected and -uninfected infants. Each HCV case was matched with up to three mothers of HCV-uninfected infants by age (+/- 5 yrs), CD4+ lymphocyte count at delivery (>500, 351-500, 200-350, and <200 cells/mm3), transmission risk (assessed through mode of delivery: scheduled cesarean section, unscheduled cesarean section, vaginal birth; and duration of time from rupture of membranes to delivery: > 4 hours, ≤ 4 hours), and time of cohort enrollment (before March 1, 1994, on or between March 1, 1994 and July 31, 1996, after July 31, 1996). If available, plasma was tested at two timepoints, one at the time of delivery and one either two or 12 months postpartum.
Laboratory studies
Antibodies to HCV were detected by a commercially available ELISA (HCV 2.0 or greater, Ortho Diagnostic Systems) in heparinized plasma. The concentration of HCV RNA was ascertained in plasma by using commercially-available assays (AMPLICOR MONITOR HCV Assay; Roche Diagnostic Systems).
HCV nAb titers were assessed in maternal plasma at the Viral Hepatitis Laboratory at Johns Hopkins. The testing laboratory was blinded to the transmission status of subjects. HCVpp were generated by co-transfection of 293T cells (ATCC) with pNL4-3.Luc.R-E- plasmid containing the env-defective HIV proviral genome and an expression plasmid containing the HCV glycoproteins (strain H77) as previously described [4,7]. Virus-containing medium was collected at 48 and 72 hours, pooled, and stored in aliquots at -80°C. Experiments were done with equivalent amounts of HCVpp as assessed by HIV p24 antigen ELISA (Aalto Bio Reagents). For neutralization experiments, two-fold dilutions of heat inactivated plasma or serum, starting at 1:50, were incubated with HCVpp for 1 hour at 37°C and added to Hep3B hepatoma cells (ATCC) for 5 hours, after which the virus-containing medium was removed. After 72 hours, cells were lysed with 50μl Cell Culture Lysis Reagent (Promega) and luciferase activity detected by addition of 50μl Luciferase Assay Reagent (Promega), measured for 5 seconds in a luminometer (Berthold Technologies). Pseudotype infection resulting in luciferase activity was measured in terms of relative light units (RLUs) in the presence of test plasma (RLUtest) versus infection in the presence of an HCV-negative normal human serum specimen (RLUcontrol) at the same dilution. Percent neutralization was calculated as 100%×[1-(RLUtest/RLUcontrol)], and endpoint neutralization titers reported as the dilution of plasma that resulted in ≥ 50% inhibition of HCVpp infectivity (ID50). If the ID50 values for paired visits from the same mother were within one dilution, neither sample was retested. Discrepant samples were repeated to confirm the ID50 at each visit or to clarify unclear results. Assays were repeated to confirm results for those specimens derived from only a single visit.
Negative control pseudoparticles expressing no envelope protein produced RLU values five orders of magnitude lower than HCVpp. Positive control pseudoparticles expressing the murine leukemia virus glycoprotein (MLVpp) were insignificantly neutralized (consistently less than 35%) by maternal plasma samples at a 1:50 dilution, demonstrating that any antiretroviral drugs present in maternal samples did not inhibit pseudoparticle infectivity.
Statistical analysis
Case and control groups were compared according to matching criteria and other factors. The distribution of maternal HCV nAb titers was assessed and reciprocal values log10 transformed to approximate a normal distribution. Titers that were below the level of detection were assigned an ID50 value of 1:25 for analysis. The rank order of titers was compared in mothers according to infant HCV status to evaluate the statistical significance of differences in distributions. Then, a conditional logistic regression model was constructed of HCV MTCT to assess the contribution of HCV nAb titer along with other factors, such as maternal plasma HCV RNA concentrations. Analyses were performed using SAS PROC PHREG.
Results
We studied plasma from 16 of 17 mothers of HCV-infected infants (one had insufficient sample). In addition, we evaluated plasma from 47 mothers of HCV-uninfected infants. HCV-transmitting mothers were similar to non-transmitting mothers in age, race, use of illicit drugs during pregnancy, CD4 lymphocyte count, mode of delivery, and gestational age at delivery (Supplemental Table 1, available online). A greater proportion of HCV-infected infants than HCV-uninfected infants also were infected with HIV, but this did not reach statistical significance.
Titers of nAb were reproducible in paired visits from maternal samples. Of 63 cases and controls, 59 mothers had paired visits (for four subjects, plasma was available only from a single visit). Of these 59 subjects, 51 (86.44%) had ID50 values that were identical (35 subjects, 59.32%) or differed by one dilution (16 subjects, 27.12%). The ID50s of the remaining eight subjects (13.56%) differed by ≥ 2 dilutions. Since the paired visits showed general agreement, we used the median ID50 value of all neutralization experiments from each mother as a single representative neutralization titer. Including all mothers, ID50 titers ranged from (undetectable- 1:12800), with a median of 1:100. A total of 13 (20.63%) mothers had undetectable neutralization titers at a 1:50 dilution of plasma.
An HCV MTCT model was constructed to examine the role of HCV-specific nAbs. As previously published, the odds of HCV transmission were increased 1.05 fold for each 105 increase in HCV viral load (P=0.03) (Table 1) [2]. HCV-specific nAbs were detected in 15 (93.75%) transmitting and 35 (74.47%) non-transmitting mothers. The median (range) of nAb titers in women who transmitted HCV, 1:125 (undetectable-1:12800), was not significantly different than in women who did not transmit HCV, 1:100 (undetectable-1:3200), (P >0.05) (Table 1, Figure 1). Multivariate analysis detected no interaction or effect modification in the associations between transmission status, maternal HCV nAb titer, and HCV viral load.
Table 1.
Association of maternal and infant factors with mother-to-infant transmission of HCV.
| HCV transmission a | ||||
|---|---|---|---|---|
| Risk factor | Yes (N=16) | No (N=47) | OR (95% CI) | P |
| Log10 maternal HCV nAb titer b | ||||
| Median | 2.10 | 2.00 | 1.8 (0.69-4.67) | 0.23 |
| Maternal HCV RNA/105 at delivery c | ||||
| Median | 19.05 | 9.60 | 1.05 (1.01-1.10) | 0.03 |
| CD4+ lymphocyte count, cells/mm3 | ||||
| Median | 427.50 | 520.00 | 1.02 (0.96-1.08) | 0.52 |
| Ethnicity | ||||
| White | 5 (31.25) | 13 (27.65) | Referent | |
| Black | 4 (25.00) | 15 (31.91) | 0.57 (0.10-3.11) | 0.52 |
| Other | 7 (43.75) | 19 (40.42) | 0.90 (0.21-3.81) | 0.89 |
| Income | ||||
| < $10,000/year | 11 (68.75) | 35 (77.77) | 1.34 (0.35-5.11) | 0.67 |
| ≥ $10,000/year | 5 (31.25) | 10 (22.22) | ||
| Cigarette (tobacco) use | ||||
| Yes | 11 (68.75) | 33 (70.21) | 1.12 (0.26-4.71) | 0.88 |
| No | 5 (31.25) | 14 (29.78) | ||
| Alcohol use | ||||
| Yes | 8 (50.00) | 28 (59.57) | 0.67 (0.20-2.27) | 0.52 |
| No | 8 (50.00) | 19 (40.42) | ||
| Hard drug use d | ||||
| Yes | 10 (62.50) | 32 (68.08) | 0.77 (0.21-2.83) | 0.69 |
| No | 6 (37.50) | 15(31.91) | ||
| Antiretroviral regimen | ||||
| None | 8 (50.00) | 25 (53.19) | Referent | |
| Single-drug therapy e | 5 (31.25) | 14 (29.78) | 1.06 (0.24, 4.60) | 0.94 |
| Combination therapy f | 0 (0.00) | 3 (6.38) | 0.00 g | 0.26 g |
| HAART h | 3 (18.75) | 5 (10.63) | 2.14 (0.28, 16.50) | 0.46 |
| STD history | ||||
| Yes | 7 (43.75) | 29 (61.70) | 0.53 (0.17-1.69) | 0.29 |
| No | 9 (56.25) | 18 (39.29) | ||
| Chorioamnionitis | ||||
| Yes | 1 (6.66) | 1 (2.22) | 2.45 (0.15-39.71) | 0.53 |
| No | 14 (93.33) | 44 (97.77) | ||
| Gestational age | ||||
| ≥ 37 weeks | 12 (75.00) | 29 (61.70) | 0.46 (0.10-2.02) | 0.30 |
| < 37 weeks | 4 (25.00) | 18 (38.30) | ||
| Use of fetal electrodes | ||||
| Yes | 1 (6.66) | 1 (2.22) | 2.45 (0.15-39.71) | 0.53 |
| No | 14 (93.33) | 44 (97.77) | ||
| Infant HIV infection status | ||||
| Infected | 5 (31.25) | 8 (17.02) | 2.59 (0.58-11.47) | 0.21 |
| Uninfected | 10 (62.50) | 39 (82.97) | ||
NOTE. Maternal factors were ascertained at final antenatal visit and refer to characteristics or behaviors during course of pregnancy. HCV= hepatitis C virus, OR= odds ratio, CI= confidence interval, nAb= neutralizing antibody, HAART= highly active antiretroviral therapy, STD= sexually transmitted disease, HIV= human immunodeficiency virus.
Data are no. (%) of subjects, unless otherwise indicated. Since information was missing for certain women, sum of measurements does not equal the total for all factors.
Statistics calculated for every log10 increase.
Statistics calculated for every 105 increase.
Cocaine, heroin or other opiates, methadone, other injecting drug use.
Single-drug therapy was defined as treatment with a single drug.
Combination therapy was defined as either two drugs, one of which could be a protease inhibitor or a nonnucleoside reverse-transcriptase inhibitor (NNRTI), or as ≥ two drugs that did not include a protease inhibitor or NNRTI, or as the combination of zidovudine, lamivudine, and abacavir.
None of the transmitting mothers received combo antiretrovirals, so P-value calculated from standard likelihood estimation routines.
HAART was defined as a treatment regimen including ≥ three drugs, one of which was a protease inhibitor or NNRTI.
Figure 1.
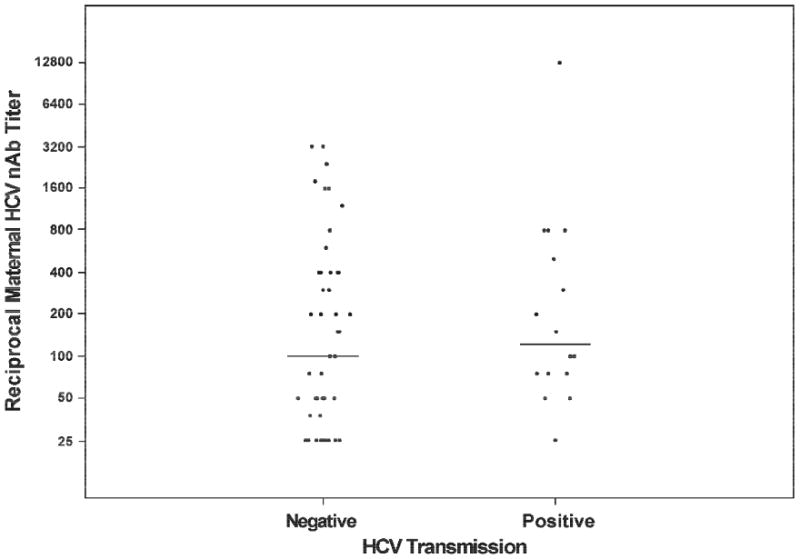
Distribution of HCV neutralizing antibody titer by HCV transmission status. Maternal plasma from two* visits (one at delivery and one at either two or 12 months postpartum) were assessed for their ability to neutralize heterologous HCV pseudoparticles expressing the envelope glycoproteins from strain H77. Points represent log10 transformed reciprocal ID50 titers for each mother, generated by combining assay results from both visits and taking the median value. Values are plotted on a log scale with the nominal values shown on the y-axis. Middle horizontal lines in each group represent median values. Mothers that transmitted HCV to their infant (n=16) did not significantly differ from non-transmitting mothers (n=47), P=0.23. *four mothers were tested at only a single visit due to sample availability.
Discussion
The results of this investigation do not confirm our hypothesis that HCV nAb titers are associated with MTCT of HCV. Since this is the first study we are aware of to investigate the role of nAbs on transmission of HCV infection, there is a limited degree to which we can discuss how our data compare with others. More work is clearly needed to understand why HCV-infected mothers transmit infection to some, but not most, infants.
There are several reasons why HCV nAb titers might not alter the frequency of HCV MTCT. Broadly reactive nAbs are readily detected in chronically-infected persons, yet the virus persists [8,11,14]. Autologous neutralization studies using HCVpp and serum from the prototype H77 infection showed ongoing viral escape from the nAb response during chronic infection [15]. We have recently corroborated these findings in multiple acutely infected subjects, suggesting that serum nAbs recognize previous, but not concurrent viral sequences due to continuing evolution of viral envelope sequences (unpublished data). In line with these results, it follows that nAbs that are ineffective at eliminating circulating virus in the mothers may be equally ineffective at preventing infection of the infant. Likewise, nAbs may efficiently block transmission of some, but not all variants, allowing infection.
The high variability of HCV viral sequences presented a technical limitation to our study. The assay measures the extent to which the mother's plasma neutralizes infection of cells by HCVpp expressing a prototype genotype 1a HCV envelope sequence (H77). Divergence between the H77 sequence and the mother's previous and current circulating viruses, particularly differences in genotype or subtype, may have resulted in an underestimation of nAb titers. In the setting of chronic HCV mono-infection, during which broadly reactive, high titer nAbs have been readily detectable, the use of heterologous HCVpp should provide an adequate representation of nAb levels. However, we have shown that co-infection with HIV lowers anti-HCV antibody titers [12], potentially limiting our detection of nAb in this study. Of note, the median titer for all mothers was 1:100, with 20.63% showing undetectable nAbs. These are much lower than titers we and others have obtained from subjects with chronic HCV mono-infection. Therefore, it is possible that the effect of HIV infection on antibody production diminished our ability to test the hypothesis in this context.
We and others have found that HCV MTCT risk is increased in mothers with higher HCV viral load and in those with concomitant HIV infection. In some, but not most, studies the mode of transmission also appears to be a factor. A strength of the present investigation of nAbs was our ability to control for or examine potential confounding factors. Since HIV infection and age may affect HCV transmission risk and nAb titer, it is important that we adjusted for these factors by matching [2,12]. We did not match based on HCV viral load in order to investigate whether nAb titer contributed to viral load and its established relationship with transmission risk. A significant correlation was detected between increased nAb titers and increasing HCV RNA levels (Spearman's rank correlation coefficient= 0.46, P=0.0002). Since higher HCV viral loads are associated with increased HCV transmission risk, this finding provides a possible explanation for higher nAb titers in HCV-transmitting mothers than HCV non-transmitting mothers (ID50 1:125 versus 1:100, respectively).
This investigation was restricted to HIV-infected women, who were found to have remarkably low titers of nAb overall. Transmission of HCV from HIV-uninfected women is rare, limiting opportunities to investigate the hypothesis in that setting. Thus, we cannot exclude the possibility that HCV nAbs are sufficient, but not necessary, to prevent MTCT of HCV, and it remains possible that the low titer nAbs we observed have a small effect on HCV transmission that would have been detected in a larger study. Nonetheless, these data clearly indicate that maternal nAb titers measured using a heterologous strain do not contribute substantially to prevention of MTCT of HCV among HIV/HCV co-infected mothers. Additional work is needed to understand the biologic basis for HCV transmission.
Supplementary Material
Supplemental Table 1. Characteristics of HCV/HIV co-infected mothers according to whether HCV was transmitted to her infant.
Acknowledgments
Grant support, including additional members of the Women and Infant Transmission Study (WITS) group: NIH (DK-068555 to D.M.N.). Principal investigators, study coordinators, program officers, and funding for the Women and Infants Transmission Study include the following: Clemente Diaz, Edna Pacheco-Acosta (University of Puerto Rico, San Juan, Puerto Rico; U01 AI 34858); Ruth Tuomala, Ellen Cooper, Donna Mesthene (Boston/Worcester Site, Boston, MA; 9U01 DA 15054); Phil LaRussa, Alice Higgins (Columbia Presbyterian Hospital, New York, NY; U01 DA 15053); Sheldon Landesman, Hermann Mendez, Edward Handelsman, Ava Dennie (State University of New York, Brooklyn, NY; HD-3-6117); Kenneth Rich and Delmyra Turpin (University of Illinois at Chicago, Chicago, IL; U01 AI 34841); William Shearer, Norma Cooper (Baylor College of Medicine, Houston, TX; U01 HD 41983); Joana Rosario (National Institute of Allergy and Infectious Diseases, Bethesda, MD); Kevin Ryan (Eunice Kennedy Shriver National Institute of Child Health and Human Development, Bethesda, MD); Vincent Smeriglio, Katherine Davenny (National Institute on Drug Abuse, Bethesda, MD); and Bruce Thompson (Clinical Trials and Surveys Corp, Baltimore, MD; N01 AI 085339). Scientific Leadership Core: Kenneth Rich, (PI), Delmyra Turpin (study coordinator) (U01 AI 50274-01). Additional support has been provided by local Clinical Research Centers as follows: Children's Hospital Boston, MA (NIH GCRC RR02174), Baylor College of Medicine, Houston, TX (NIH GCRC RR00188) and Columbia University, New York, NY (NIH GCRC RR00645).
Footnotes
Authors do not have commercial or other associations that might pose a conflict of interest.
This work has not been previously presented.
Reference List
- 1.Mast EE, Hwang LY, Seto DS, et al. Risk factors for perinatal transmission of hepatitis C virus (HCV) and the natural history of HCV infection acquired in infancy. J Infect Dis. 2005;192:1880–9. doi: 10.1086/497701. [DOI] [PubMed] [Google Scholar]
- 2.Thomas DL, Villano SA, Riester KA, et al. Perinatal transmission of hepatitis C virus from human immunodeficiency virus type 1-infected mothers. J Infect Dis. 1998;177:1480–8. doi: 10.1086/515315. [DOI] [PubMed] [Google Scholar]
- 3.Zanetti AR, Tanzi E, Paccagnini S, et al. Mother-to-infant transmission of hepatitis C virus. Lancet. 1995;345:289–91. doi: 10.1016/s0140-6736(95)90277-5. [DOI] [PubMed] [Google Scholar]
- 4.Netski DM, Mosbruger T, Depla E, et al. Humoral immune response in acute hepatitis C virus infection. Clin Infect Dis. 2005;41:667–75. doi: 10.1086/432478. [DOI] [PubMed] [Google Scholar]
- 5.Cox AL, Mosbruger T, Mao Q, et al. Cellular immune selection with hepatitis C virus persistence in humans. J Exp Med. 2005;201:1741–52. doi: 10.1084/jem.20050121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bartosch B, Dubuisson J, Cosset FL. Infectious hepatitis C virus pseudo-particles containing functional E1- E2 envelope protein complexes. J Exp Med. 2003;197:633–42. doi: 10.1084/jem.20021756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hsu M, Zhang J, Flint M, et al. Hepatitis C virus glycoproteins mediate pH-dependent cell entry of pseudotyped retroviral particles. Proc Natl Acad Sci U S A. 2003;100:7271–6. doi: 10.1073/pnas.0832180100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Logvinoff C, Major ME, Oldach D, et al. Neutralizing antibody response during acute and chronic hepatitis C virus infection. Proc Natl Acad Sci U S A. 2004;101:10149–54. doi: 10.1073/pnas.0403519101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lavillette D, Morice Y, Germanidis G, et al. Human serum facilitates hepatitis C virus infection, and neutralizing responses inversely correlate with viral replication kinetics at the acute phase of hepatitis C virus infection. J Virol. 2005;79:6023–34. doi: 10.1128/JVI.79.10.6023-6034.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Pestka JM, Zeisel MB, Blaser E, et al. Rapid induction of virus-neutralizing antibodies and viral clearance in a single-source outbreak of hepatitis C. Proc Natl Acad Sci U S A. 2007;104:6025–30. doi: 10.1073/pnas.0607026104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Meunier JC, Engle RE, Faulk K, et al. Evidence for cross-genotype neutralization of hepatitis C virus pseudo-particles and enhancement of infectivity by apolipoprotein C1. Proc Natl Acad Sci U S A. 2005;102:4560–5. doi: 10.1073/pnas.0501275102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Netski DM, Mosbruger T, Astemborski J, Mehta SH, Thomas DL, Cox AL. CD4+ T cell-dependent reduction in hepatitis C virus-specific humoral immune responses after HIV infection. J Infect Dis. 2007;195:857–63. doi: 10.1086/511826. [DOI] [PubMed] [Google Scholar]
- 13.Hershow RC, Riester KA, Lew J, et al. Increased vertical transmission of human immunodeficiency virus from hepatitis C virus-coinfected mothers. J Infect Dis. 1997;176:414–20. doi: 10.1086/514058. [DOI] [PubMed] [Google Scholar]
- 14.Bartosch B, Bukh J, Meunier JC, et al. In vitro assay for neutralizing antibody to hepatitis C virus: evidence for broadly conserved neutralization epitopes. Proc Natl Acad Sci U S A. 2003;100:14199–204. doi: 10.1073/pnas.2335981100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.von Hahn T, Yoon JC, Alter H, et al. Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection in vivo. Gastroenterology. 2007;132:667–78. doi: 10.1053/j.gastro.2006.12.008. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Table 1. Characteristics of HCV/HIV co-infected mothers according to whether HCV was transmitted to her infant.