

Abstract
It is not certain whether coral reefs are sources of or sinks for atmospheric CO2. Air–sea exchange of CO2 over reefs has been measured directly and inferred from changes in the seawater carbonate equilibrium. Such measurements have provided conflicting results. We provide community metabolic data that indicate that large changes in CO2 concentration can occur in coral reef waters via biogeochemical processes not directly associated with photosynthesis, respiration, calcification, and CaCO3 dissolution. These processes can significantly distort estimates of reef calcification and net productivity and obscure the contribution of coral reefs to global air–sea exchange of CO2. They may, nonetheless, explain apparent anomalies in the metabolic performance of reefs close to land and reconcile the differing experimental findings that have given rise to the CO2 debate.
There is debate as to whether coral reefs are sources of or sinks for atmospheric CO2 (1–4). They may take up about 2% of the annual anthropogenic production of CO2 if they are sinks (5) or they may release up to 8% if they are sources (6). Gross productivity on coral reefs is among the highest for natural ecosystems (7) and photosynthesis by reef benthos encourages invasion of CO2 from the atmosphere by reducing its concentration in overlying seawater. Respiration and formation of reef rock (calcification) have the reverse effect. Although calcification decreases the overall concentration of inorganic carbon in seawater, associated acidification,
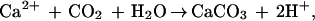 |
increases the amount present as dissolved (gaseous) CO2.
Measurements of organic and inorganic carbon metabolism on different reefs have yielded conflicting data on the direction of the net CO2 flux (1, 8). Metabolism on the Tiahura fringing barrier reef, Moorea, apparently released CO2 to the atmosphere (8), whereas metabolism on Shiraho Reef, Ishigaki Island, Japan, had apparently the reverse effect (1). Most researchers consider that reefs are a source of CO2 (2, 3, 6, 8, 9). Critics of the Shiraho Reef study argue (i) that the reef must have been dominated by noncalcareous algae that increased the ratio of organic to inorganic carbon metabolism, (ii) that the measurements were not representative of the whole reef, and (iii) that erroneous conclusions were drawn from inadequate data (2, 3). Critics also suggest that insufficient measurements were taken to differentiate changes in the concentration of CO2 caused by benthic metabolism from natural variability in the CO2 concentration of seawater flowing onto the reef (2). Overall, these criticisms follow from the general view that coral reefs are sources of CO2. The net air–sea flux of CO2 is thought to be controlled by calcification because the ratio of photosynthesis to respiration on unperturbed reefs over 24 h is considered to be close to unity (10).
In March 1996, we made an expedition to Lizard Island, northern Great Barrier Reef, Australia (Fig. 1), to measure changes in the O2 concentration and pH of seawater flowing across a 300-m section of the reef flat by using a floating instrument package (11–14). Measurements were made in March when tides permitted the instrument package to float freely over the reef flat, a short distance above the benthos.
Figure 1.

Map of Lizard Island, adjacent land formations, and surrounding reefs showing the location of the experimental transect. (Inset) Location of Lizard Island relative to the Australian mainland (minimum distance = 15 nautical miles); a detailed description of the reef system at Lizard island and of the benthic communities present on the transect is provided elsewhere (11).
On arriving at Lizard Island, we encountered environmental conditions that we had not anticipated. Almost twice as much rain had fallen at Lizard Island between January 1, 1996, and March 14, 1996 (1,097 mm), as had fallen, on average, in each of the 10 previous summers (January 1 to 31 March; mean rainfall = 599 mm). Moreover, 42% of this rain fell in the 12 days that preceded our measurements (Fig. 2A). The salinity of seawater around Lizard Island was 3‰ below normal on the day before measurements began (i.e., 32‰), rising to 1‰ below normal by the time measurements were completed (Fig. 2B). Concurrence of unusually low tides and heavy rains had caused reef flat organisms to be directly exposed to freshwater or to greatly reduced salinities. In addition, seawater temperature had been high, close to 30°C, over the previous 3 months (Fig. 2B) and reef flat coral communities had been recently attacked by crown of thorns starfish. In consequence, live coral cover on the reef flat was very low (0–10%) and there was a considerable amount of dead coral overgrown by filamentous algae.
Figure 2.

Environmental conditions before, during, and after measurements were made (shaded regions) of reef flat community metabolism at Lizard Island. (A) Rainfall on Lizard Island between January 1, 1996, and April 1, 1996 (15). (B) Measured seawater surface temperatures and salinities in the lagoon located between Lizard Island, South Island, and Palfrey Island (see Fig. 1) between June 1, 1993 and June 1, 1996 (16); values shown during the experimental period were obtained on March 18, 1996, 4 days after measurements began and 3 days before measurements were completed. A salinity of 32‰ was recorded on March 13, 1996 (11).
We began floating the instrument package across the reef flat with the incoming tide shortly after the heavy rain ceased (Fig. 2A). The package was floated across the reef at various times of the day and night to determine diel rates of benthic photosynthesis, respiration, calcification, and solution of reef rock. These parameters were estimated from equations for the seawater carbonate equilibrium (17) wherein ΔpH describes the total change in all CO2 species resulting from both organic and inorganic carbon metabolism and ΔO2 multiplied by the appropriate metabolic quotient describes the change in CO2 due to organic metabolism (12). The advantage of this pH–O2 technique is that all necessary measurements can be made with electrodes. Its disadvantage is that the photosynthetic and respiratory quotients (PQ and RQ) must be used to convert measurements of ΔO2 to ΔCO2. These quotients are estimated by adjusting their values until estimates of calcification at saturating light intensities and at night accord with total alkalinity-based (TA) measurements (calcification = 0.5ΔTA) (13). Accordingly, water samples for determination of TA were taken at 100-m intervals as the instrument package crossed the reef on two transects around noon and on two transects shortly after dusk.
Subsequent analysis of data revealed that the changes in TA of seawater crossing the reef flat at Lizard Island were so large that they provided unrealistic values for the metabolic quotients. We thus estimated organic and inorganic carbon metabolism from measurements of ΔO2 and ΔpH by assuming PQ = 1.0 or 1.1 and RQ = 0.9 or 1.0. These values encompass most previous estimates of PQ and RQ for coral reef communities (10). Data given herein are for PQ = RQ = 1.0. Use of PQ = 1.1 and RQ = 0.9 decreased hourly estimates of photosynthesis and respiration by ≤10% and peak rates of calcification by <20%. The global standards for coral reef metabolism are gross productivity (P) = 6–8 g of C per m2 per day and calcification (G) ≈ 3–5 kg of CaCO3 per m2 per year (10). Our pH–O2 data gave P = 9.3 g of C per m2 per day and G ≈ 9.3 kg of CaCO3 per m2 per year. Thus, our measurement of productivity was marginally high while that of calcification was about 2–3 times the rate expected on a healthy reef flat. Measurements of calcification from ΔTA gave a rate about 3–5 times higher (G ≈ 15.1 kg of CaCO3 per m2 per year) than the suggested standard.
Total calcification during daylight, estimated from pH–O2 data (41.0 g of CaCO3 per m2), was similar to that estimated from ΔTA (36.8 g of CaCO3 per m2). However, the two techniques gave very different rates of nighttime calcification (Fig. 3): pH–O2 data indicated solution of reef rock (−15.5 g of CaCO3 per m2), whereas TA data indicated net precipitation (4.7 g of CaCO3 per m2).
Figure 3.

Estimated cumulative precipitation (positive increments) or dissolution (negative increments) of CaCO3 across the reef flat transect during the middle of the day (mean irradiance = 1811 μmol per m2 per second) (A) and the first part of the night (B). Small symbols indicate estimates based upon pH–O2 measurements assuming PQ = RQ = 1.0; large symbols indicate estimates based upon ΔTA measurements; vertical bars indicate 95% confidence intervals; fitted lines are third-degree polynomials.
Both techniques thus indicated that the impoverished heavily stressed reef flat at Lizard Island had a much higher rate of calcification than pristine reefs growing in clear ocean water (10). Moreover, these estimates were made by using the same techniques that have fired the coral reef CO2 source-sink debate. Measurements of coral reef CO2 dynamics are founded upon the assumption that changes in the inorganic carbon content of seawater over reefs are almost entirely due to photosynthesis, respiration, calcification, and solution of reef rock (18–21). Our results indicate that other biogeochemical processes can significantly alter the seawater carbonate equilibrium by affecting total alkalinity.
The most likely of these is organic matter decomposition (22–24) that leads to the formation of phosphoric acid,
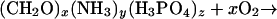 |
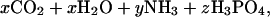 |
and a considerably greater amount of nitric acid via oxidation of ammonium by nitrifying bacteria,
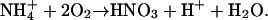 |
Substantial decomposition of organic matter was almost certainly occurring on the reef flat at Lizard Island because the various stresses, especially heavy rainfall and low salinity (Fig. 2), had caused widespread mortality of organisms upon and within the reef matrix.
At night, the pH–O2 technique measures absolute ΔO2 and, consequently, O2 consumed by nitrification is inadvertently assigned to respiration. The acid products of nitrification and respiration are, respectively, H+ and hydrated CO2. H+ and CO2 have similar effects upon seawater pH (25). The change in CO2 associated with nitrification is thus correctly predicted from O2 data in the dark. Thus, calcification is reliably approximated. This is not the case in the light when photosynthesis and respiration occur simultaneously because net ΔO2 is measured. Daytime measurements fail to record oxygen consumption by nitrification and its attendant effects on pH and total alkalinity. This causes net productivity to be underestimated and calcification to be overestimated. The underestimate of net productivity during the day, however, equals the overestimate of respiration at night provided that the rate of nitrification does not vary diurnally. Given significant nitrification, the pH–O2 method, therefore, reasonably estimates calcification in the dark and gross productivity over 24 h but incorrectly estimates respiration at night and net productivity and calcification during the day. This is not so with the alkalinity method: the reduction in total alkalinity associated with nitrification is incorrectly attributed to calcification irrespective of whether it is day or night.
This fundamental difference between the pH–O2 and TA techniques provides two ways to evaluate the error present in daytime estimates of calcification. If a reasonable value for RQ (e.g., 1.0) is used to convert nighttime measurements of ΔO2 into ΔCO2 in the pH–O2 method, the reduction in pH caused by nitrification is assigned to respiration and calcification is correctly estimated. Subtracting this rate of calcification from that indicated by ΔTA in the dark provides an estimate of the error due to nitrification. Alternatively, the error present in daytime estimates of calcification may be evaluated by assuming that PQ is the reciprocal of the determined value for RQ estimated by aligning pH–O2 data with ΔTA. Because this average value for RQ was 0.64, PQ was assumed to be 1.56. Applying these two methods of correction to the pH–O2 data yielded calcification rates of 1.9 and 2.1 kg of CaCO3 per m2 per year, respectively, for the reef flat at Lizard Island during the study.
Clearly, some broad assumptions have been made, the most significant being that nitrification was solely responsible for the inferred overestimate of calcification and that rates of nitrification do not change significantly between day and night. Inclusion of the effects of phosphoric acid production (23) and enhanced rates of inorganic nitrogen uptake during the day (26) would further reduce estimates of reef flat calcification unless, in the latter case, nitrate were taken up in preference to ammonium. We suspect, however, that neither phosphoric acid production nor diurnal variation in NH4+ uptake would seriously affect our estimates. This is because considerably less phosphoric acid than nitric acid is produced during organic decomposition and because rates of calcification in the dark were measured just after dusk when rates of nitrification were presumably closest to those of the day. Straightforward uptake of NH4+ by algae (including zooxanthellae) could have contributed to alkalinity depletion but this would have required a large exogenous supply of NH4+ to the reef† and it would not have accounted for the difference in nighttime estimates of calcification using the pH–O2 and ΔTA techniques.‡
Further error may have resulted from determination of RQ and nighttime rates of calcification near the end of the experimental period when seawater salinities had returned close to normal. Measurements made under high irradiance were mainly carried out near the beginning of the experimental period when suitable tidal cycles permitted deployment of the instrument package near the solar zenith. Probably not by coincidence, highest apparent rates of calcification were obtained at this time, when biogeochemical processes distorting the seawater carbonate equilibrium were likely to have been most intense.
In spite of these uncertainties, our data indicate that perturbation of reef ecosystems can produce significant anomalies in community metabolic performance. This conclusion derives from the following aspects of our data: (i) a need to use impossibly low values for RQ to make nighttime pH–O2 estimates of calcification equate with ΔTA measurements, (ii) unacceptably high rates of nighttime calcification indicated by ΔTA measurements, and (iii) unusually high rates of daytime calcification indicated by both pH–O2 and ΔTA measurements.
The kinds of processes that have been inferred here to explain the anomalies that we noted in reef flat community metabolism have been shown to exert considerable influence on seawater chemistry in other coastal and estuarine systems (e.g., refs. 27 and 28). Our data suggest that their significance in coral reef environments, particularly those fringing land, may have been underestimated. The notion that coral reefs are largely autonomous structures, dependent upon the meager supplies of nutrients carried by tropical ocean currents, may be incorrect. Unusual events that make available large amounts of nutrients to coral reefs may produce short-term impacts but long-term benefits, as they do to flood plains, estuaries, and certain other coastal ecosystems.
It has been noted that high rates of productivity and calcification are sometimes associated with proximity to land (10). Equivalent procedures to those used here also gave unusually high rates of calcification for fringing reefs at Moorea and Eilat (8.9 and 9.7 kg of CaCO3 per m2 per year, respectively) (8, 14), suggesting that similar biogeochemical processes may have been occurring on these near-shore reefs. Certainly, there is evidence that the fringing reef at Moorea has been degraded by human activities (29) and widely varying estimates of reef flat calcification and productivity have been obtained at different times (6). In contrast, a fringing reef at Ishigaki Island, Japan, apparently precipitated a “standard” 3.7 kg of CaCO3 per m2 per year but fixed only 2.5 g of C per m2 per day (1).
An interesting observation emerges from these data. Data from Moorea and Ishigaki Island are at the center of the debate as to whether reefs are sources of, or sinks for, atmospheric CO2. Shiraho Reef at Ishigaki Island exhibited “normal” calcification, carbon fixation at about one-third of the “standard” rate and it was apparently a sink for CO2. A fringing reef at Moorea had slightly above “normal” carbon fixation, calcification was around twice the “standard” rate, and it was apparently a source of CO2. These findings suggest that the opposing views in the “coral reefs as sources of or sinks for CO2 debate” may both be correct in terms of their respective data sets—but that neither data set truly represents the contribution of coral reef growth to global air–sea CO2 exchange. Data presented herein question the long-standing assumption that the carbonate equilibrium of seawater above most coral reefs is principally controlled by photosynthesis, respiration, calcification, and solution of reef rock. Standard pH–O2 and TA techniques may not provide accurate data for the metabolic performance of reefs that lie close to land. Further, these techniques are likely to provide inaccurate data on oceanic reefs in the event of significant nutrient import or turnover.
It may, nonetheless, be possible to use these very excursions from standard rates of production and calcification to deduce the causes of reef perturbation. For example, high productivity and low calcification may indicate strong algal uptake of nitrate supplied by agricultural land run-off or deep-water upwelling (note: uptake of NO3− lowers seawater total alkalinity), whereas high productivity and high calcification may indicate strong algal and microbial uptake of NH4+ supplied by organic matter decomposition or sewage discharge.
Most of these ideas rest on the notion that mature reefs exhibit similar rates of community metabolism except when perturbed. They further stand on the assumption that production and consumption are balanced in healthy reefs and thus that inorganic carbon metabolism drives air–sea fluxes of CO2. The assumption that reefs have P/R ratios that verge upon unity cannot be correct if they continue to increase in dimension. Reef flats, on the other hand, are likely to have P/R ratios close to 1.0 because they do not add significant biomass after reaching the sea surface. Reliable estimation of the role of reefs in air–sea CO2 exchange requires measurement of the metabolism of whole reefs that are not perturbed.
These arguments suggest that reef health might be most effectively investigated by analysis of their community metabolic quotients. In lieu of the development of a good field CO2 electrode, this requires measurement of O2, TA, and pH or O2, TA, and pCO2. These combined measurements are further needed to confirm the “normality” of a reef’s metabolic performance before extrapolating measured rates of air–sea CO2 exchange to reefs in general.
Overall, reef growth may serve to drive CO2 into the atmosphere but probably to a lesser extent than has been indicated by Lagrangian measurements of reef flat metabolism at Moorea (31 mmol per m2 per day) and Yonge Reef on the GBR (182 mmol per m2 per day) (6). Comparative studies using a Eulerian approach indicated rates of CO2 efflux that were lower by 1–2 orders of magnitude (1.81 mmol per m2 per day at Moorea and 5.1 mmol per m2 per day at Yonge) (9). If these latter data are the more representative, then inclusion of high net organic productivity on the seaward slopes of reefs (30) may indicate that whole reefs act as sinks for atmospheric CO2. The fact that carbonate rocks store 3 × 104 more inorganic carbon than the atmosphere (17) shows that reefs are sinks for CO2 over geological time.
Acknowledgments
We thank M. J. Devereux, B. A. King, J. M. Lough, B. J. Tobin, the Directors and Staff of the Lizard Island Research Station, and the AIMS Graphics Section for technical assistance. This work was carried out under cooperative research agreement between the Japanese Association of Marine Sciences and Technologies and the Australian Institute of Marine Science.
Footnotes
Bacterial decomposition of organic nitrogen produces ammonium and hydroxyl ions, whereas uptake of NH4+ by algae leads to formation of organic nitrogen and protons, thus significant alkalinity depression does not occur unless the processes of NH4+ formation and uptake are separated in space or time.
Nitrification consumes O2, whereas uptake of NH4+ by algae does not. Therefore, the resulting production of H+ and consequential drop in TA would not be concomitant with reductions in the values of RQ and PQ.
References
- 1.Kayanne H, Suzuki A, Saito H. Science. 1995;269:214–216. doi: 10.1126/science.269.5221.214. [DOI] [PubMed] [Google Scholar]
- 2.Gattuso J-P, Frankignoulle M, Smith S V, Ware J R, Wollast R. Science. 1996;271:1298. doi: 10.1126/science.271.5253.1298a. [DOI] [PubMed] [Google Scholar]
- 3.Buddemeier R W. Science. 1996;271:1298–1299. doi: 10.1126/science.271.5253.1298b. [DOI] [PubMed] [Google Scholar]
- 4.Kayanne H, Suzuki A, Saito H. Science. 1996;271:1299–1300. doi: 10.1126/science.271.5253.1299. [DOI] [PubMed] [Google Scholar]
- 5.Kinsey D W, Hopley D. Palaeogeogr Palaeoclimatol Palaeoecol. 1991;89:363–377. [Google Scholar]
- 6.Gattuso J-P, Pichon M, Dellesalle B, Canon C, Frankignoulle M. Mar Ecol Prog Ser. 1996;145:109–121. [Google Scholar]
- 7.Lewis J B. Biol Rev. 1977;52:305–347. [Google Scholar]
- 8.Gattuso J-P, Pichon M, Dellesalle B, Frankignoulle M. Mar Ecol Prog Ser. 1993;96:259–267. [Google Scholar]
- 9.Frankignoulle M, Gattuso J-P, Biondo R, Bourge I, Copin-Montégut G, Pichon M. Mar Ecol Prog Ser. 1996;145:123–132. [Google Scholar]
- 10.Kinsey D W. Proc Fifth Int Coral Reef Congress (Tahiti) 1985;4:505–526. [Google Scholar]
- 11.Chisholm, J. R. M., Barnes, D. J. & Devereux, M. J. (1996) Measurement and Analysis of Reef Flat Community Metabolism at Lizard Island, Great Barrier Reef, Australia, Phase 2 Report to Japanese Association of Marine Sciences and Technologies, Australian Institute of Marine Science Contribution 857.
- 12.Barnes D J. J Exp Mar Biol Ecol. 1983;66:149–161. [Google Scholar]
- 13.Barnes D J, Devereux M J. J Exp Mar Biol Ecol. 1984;79:213–231. [Google Scholar]
- 14.Barnes D J, Lazar B J. Exp Mar Biol Ecol. 1993;174:1–13. [Google Scholar]
- 15.Lizard Island Research Station, Private Mail Bag 37, Cairns, Q. 4870, Australia.
- 16.Great Barrier Reef Marine Park Authority, Sir Leslie Thiess Drive, Townsville, Q. 4810, Australia.
- 17.Skirrow G. In: Chemical Oceanography 2. Riley J P, Skirrow G, editors. London: Academic; 1978. pp. 1–192. [Google Scholar]
- 18.Kinsey D W. In: Coral Reef: Research Methods. Stoddart D R, editor. Paris: UNESCO; 1978. pp. 439–468. [Google Scholar]
- 19.Smith S V. Limnol Oceanogr. 1973;18:106–120. [Google Scholar]
- 20.Smith S V, Key G S. Limnol Oceanogr. 1975;20:493–495. [Google Scholar]
- 21.Smith S V, Kinsey D W. In: Coral Reef: Research Methods. Stoddart D R, editor. Paris: UNESCO; 1978. pp. 469–484. [Google Scholar]
- 22.Brewer P G, Wong G T F, Bacon M P, Spencer D W. Earth Planet Sci Lett. 1975;26:81–87. [Google Scholar]
- 23.Brewer P G, Goldman J C. Limnol Oceanogr. 1975;21:108–117. [Google Scholar]
- 24.Goldman J C, Brewer P G. Limnol Oceanogr. 1980;25:352–357. [Google Scholar]
- 25.Frankignoulle M. J Mar Syst. 1994;5:111–118. [Google Scholar]
- 26.Larkum A W D, Kennedy I R, Muller W J. Mar Biol. 1988;98:143–155. [Google Scholar]
- 27.Fenchel T, Blackburn T H. Bacterial and Mineral Cycling. New York: Academic; 1979. [Google Scholar]
- 28.Day J W, Jr, Hall C A S, Kemp W M, Yáñez-Arancibia A. Estuarine Ecology. New York: Wiley; 1989. pp. 111–116. [Google Scholar]
- 29.Wolanski E, Dellesalle B, Dufour V, Aubanel A. Proc 11th Australasian Conference Coastal Ocean Engineering. 1993;4:583–588. [Google Scholar]
- 30.Kinsey D W, Davies P J. In: Biogeochemical Cycling of Mineral-Forming Elements. Trudinger P A, Swaine D J, editors. Amsterdam: Elsevier; 1979. pp. 131–162. [Google Scholar]