
Figure 1.
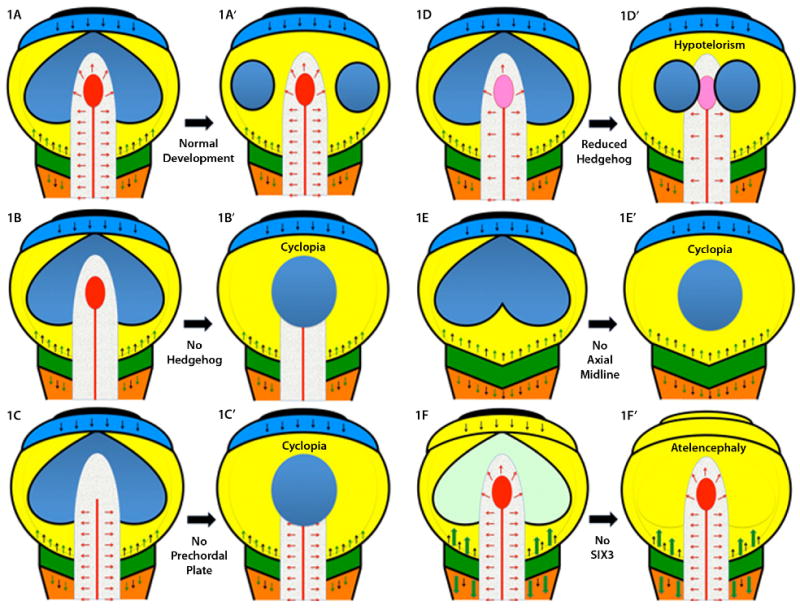
A: A schematic representation of a typical flat neural plate prior to the neurulation stage, viewed from above, is shown with key signaling centers indicated by various colors. Anterior is at the top and the hindbrain (orange) and spinal chord at the bottom. Each pair of figures illustrates the effects of the loss of key genes/structures in a simplified developmental progression. The most rostral center, the anterior neural ridge (ANR), is in black and secretes FGF8 (black arrows, also produced at the midbrain-hindbrain boundary [MHB], which is dark green) and the anti-WNT factor TLC (not shown) [Houart et al., 2002]. The ANR promotes the growth and expansion of the telencephalon (shown as an undivided light blue strip of tissue). Note that the eye field, shown in dark blue, is also contiguous across the midline just caudal to the telencephalon within the mesencephalon/midbrain territory (yellow). The MHB produces several important caudalizing factors, including WNTs (green arrows), whose actions are contained by rostral inhibitors (DKK1, TLC, SIX3, etc). The paraxial mesoderm (caudal to the MHB) is a source of retinoic acid that patterns the hindbrain, but is eliminated anterior to the MHB by the enzyme Cyp26. The axial midline (notochord as red line, and prechordal plate, PCP as red ellipse) is a source of hedgehog ligands (red arrows) as well as anti-BMP factors such as chordin, follistatin and noggin (not shown, see Klingensmith chapter). Definitive ectoderm is a source of BMP activity (thin black line surrounding each diagram) and assumes its exterior location, with neural tissue interior, following neurulation [England et al., 2006]. A′: If the PCP correctly migrates beneath the telencephalon, then secondary signals, including a secondary domain of hedgehog activity in the ventral telencephalon, accompanies the division of the eye field into two discrete eyes. B: If hedgehog signals (red arrows) are eliminated, genetically or pharmacologically, the eye field fails to divide resulting in cyclopia, see B′. Hedgehog activity is also required in the developing face [Cordero et al., 2004] linking additional signaling centers to the function of the PCP. C: Surgical ablation of the PCP also results in cyclopia [Shih and Fraser, 1995], see C′. D and D′: Attenuated hedgehog signaling is compatible with some eye field separation, although the eyes are closely spaced (hypotelorism). E and E′: If the axial midline fails to form, or degenerates, the resulting animals are uniformly cyclopic [Israeli et al., 1999; Karkera et al., 2002]. F: The importance of WNT inhibition is illustrated by model organisms lacking Six3 where the telencephalon is truncated under the un-restrained activity (large green arrows) of caudalization signals. To the extent that anterior eye field forms under these circumstances (faint green) these anterior structures soon disappear, see F′. Interestingly, one WNT inhibitor, DKK1, is not a common mutational target in humans [Roessler et al., 2000]. Excessive exposure to retinoids can also result in atelencephaly, or HPE [Sulik et al., 1995], possibly through a similar mechanism of caudalization.