
Figure 3.
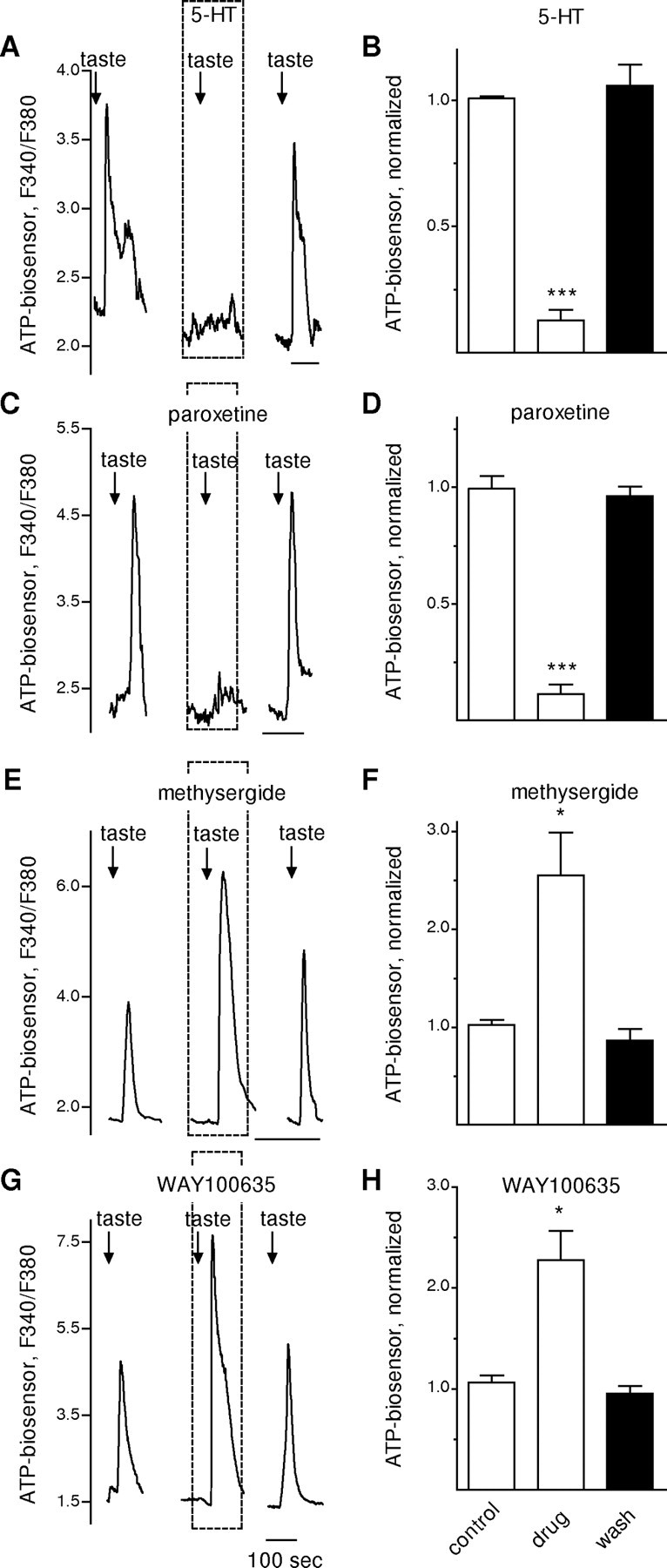
Manipulating extracellular serotonin or blocking 5-HT1A receptors in taste buds modulates ATP neurosecretion. Whole taste buds were isolated from mouse vallate papillae to retain cell-to-cell communication between receptor and presynaptic cells (Y. J. Huang et al., 2007). Traces show taste-evoked secretion of ATP from the receptor cells in a taste bud, measured with an ATP biosensor positioned against the taste bud. A, Responses of an ATP biosensor cell positioned next to a taste bud. Similar to the results shown in Figure 2A for isolated single receptor cells, the presence of bath-applied 5-HT (10 nm, perfused throughout the interval shown by dashed box) reversibly reduced taste-evoked ATP secretion from the receptor cells within the taste bud. Arrows, Taste stimulation. B, Summary of data from eight experiments. Bars show means ± SEM for ATP biosensor signals in response to taste stimulation (as in A), normalized to the average taste-evoked response in absence of drug (i.e., control). C, Paroxetine, which blocks 5-HT reuptake and thus would be expected to lead to an accumulation of extracellular 5-HT released during taste stimulation, mimics the actions of bath-applied 5-HT. As shown here, perfusion with paroxetine (100 nm, dashed box) reduced taste-evoked ATP secretion. D, Summary of data from five experiments, plotted as in B. E, Methysergide, a 5-HT1 and 5-HT2 receptor antagonist, greatly enhances ATP secretion from taste buds. Here, bath-applied methysergide (10 nm) nearly doubled taste-evoked ATP secretion from the taste bud. F, Summary of data from five experiments, plotted as in B. G, WAY100635, a 5-HT1A selective antagonist, also enhances taste-evoked ATP secretion. Here, WAY100635 (10 nm) produced a large increase in ATP secretion. H, Summary of data from five experiments, plotted as in B. *p < 0.05; ***p < 0.001; Student's paired t test.