
Figure 4.
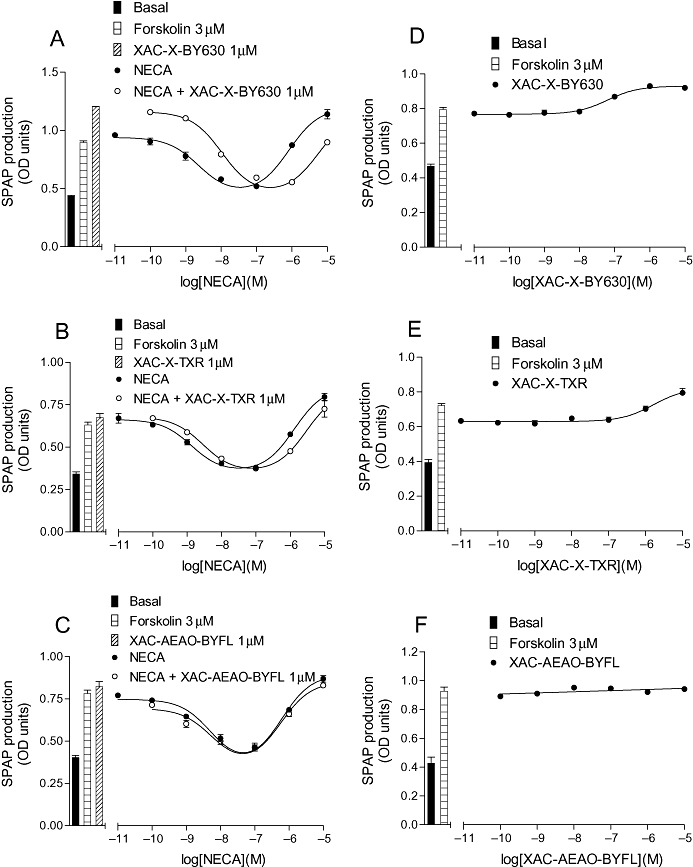
CRE-SPAP production in CHO-A1 cells in response to (A) NECA in the presence and absence of 1 µM XAC-X-BY630 (14); (B) NECA in the presence and absence of 1 µM XAC-X-Texas Red (XAC-X-TXR; 15); (C) NECA in the presence and absence of 1 µM XAC-AEAO-BYFL (17); (D) XAC-X-BY630 (14); (E) XAC-X-Texas Red (XAC-X-TXR) and (F) XAC-AEAO-BYFL (17). Bars show basal CRE-SPAP production, that in response to 3 µM forskolin, and for A–C, that in response to 1 µM of the XAC-derivative in the presence of 3 µM forskolin. Data points are mean ± SEM from triplicate values from a single experiment. These separate experiments are representative of (A) 9, (B) 6, (C) 10, (D) 9, (E) 6 and (F) 6 separate experiments. AEAO, 8-(2-aminoethylamino)-8-oxooctanoyl; CHO, Chinese hamster ovary; CRE-SPAP, cyclic 3′,5′ adenosine monophosphate response element-secreted placental alkaline phosphatase; NECA, 5′ (N-ethylcarboxamido)adenosine; X, 6-aminohexanoyl; XAC, xanthine amine congener.