
Table 2.
Parameters in the model. Time is expressed in years, weight in grams, and biomass density in grams per unit volume.
| individual physiology | ||
| α | 0.6 | assimilation efficiencya |
| n | 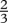 |
exponent of max. intakeb |
| h | 30 | factor for max. intakec |
| p | 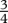 |
exponent of standard metabolismd |
| ks | 4 | factor for standard metabolisme |
| μ0 | 0.6 | background mortality factorf |
| η | 0.25 | size at maturation rel. to Mg |
| individual foraging | ||
| β | 100 | preferred predator–prey weight ratioh |
| σ | 1.3 | width of selection functioni |
| γ | 526 | factor for search volumej |
| q | 0.9 | exponent of search volumek |
| primary production | ||
| κr | 0.005 | resource spectrum carrying capacity |
| r0 | 4 | growth rate of resource spectruml |
bJobling (1994) states that 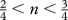 . We have used n=
. We have used n=  to be consistent with von Bertalanffy growth curves.
to be consistent with von Bertalanffy growth curves.
cAdjusted such that emergent growth rates are in the range of those observed in the North Sea (figure 1).
eThe data of Winberg (1956) indicate a standard (resting) metabolism factor for fish of about 4 g0.25 yr−1 at 10°C.
fThis has been adjusted to lead to a background mortality of the same order as the predation mortality, but still lower than the predation mortality.
iUrsin (1973) finds σ = 1 for a single species. To account for species diversity within an asymptotic size class, this has been increased to σ = 1.3.
jSee equation (2.1).
kConsiderations on the bioenergetic budget of swimming predict a value of q between  and 1 (Andersen & Beyer 2006).
and 1 (Andersen & Beyer 2006).
lSavage et al. (2004; temperature: 10°C).