

Abstract
The classic view for hypothalamic regulation of anterior pituitary (AP) hormone secretion holds that release of each AP hormone is controlled specifically by a corresponding hypothalamic-releasing hormone (HRH). In this scenario, binding of a given HRH (thyrotropin-, growth hormone-, corticotropin-, and luteinizing hormone-releasing hormones) to specific receptors in its target cell increases the concentration of cytosolic Ca2+ ([Ca2+]i), thereby selectively stimulating the release of the appropriate hormone. However, “paradoxical” responses of AP cells to the four well-established HRHs have been observed repeatedly with both in vivo and in vitro systems, raising the possibility of functional overlap between the different AP cell types. To explore this possibility, we evaluated the effects of HRHs on [Ca2+]i in single AP cells identified immunocytochemically by the hormone they stored. We found that each of the five major AP cell types contained discrete subpopulations that were able to respond to several HRHs. The relative abundance of these multi-responsive cells was 59% for lactotropes, 33% for thyrotropes, and in the range of 47–55% for gonadotropes, corticotropes, and somatotropes. Analysis of prolactin release from single living cells revealed that each of the four HRHs tested were able to induce hormone release from a discrete lactotrope subpopulation, the size of which corresponded closely to that in which [Ca2+]i changes were induced by the same secretagogues. When viewed as a whole, our diverse functional measurements of multi-responsiveness suggest that hypothalamic control of pituitary function is more complicated than previously envisioned. Moreover, they provide a cellular basis for the so-called “paradoxical” behavior of pituitary cells to hypothalamic hypophysiotropic agents.
Keywords: calcium, thyrotropin-releasing hormone, growth hormone, prolactin
The pituitary gland is a major neuroendocrine modulator that regulates many peripheral glands and tissues through the secretion of anterior pituitary (AP) hormones. These hormones include prolactin (PRL), growth hormone (GH), thyrotropin (TSH), adrenocorticotropin, and the gonadotropins [follicle-stimulating hormone (FSH) and luteinizing hormone (LH)]. The secretory activity of the gland is controlled, in turn, by signals derived from the hypothalamus in the form of hypothalamic-releasing hormones (HRHs) such as thyrotropin-releasing hormone (TRH), growth hormone-releasing hormone (GHRH), corticotropin-releasing hormone (CRH), and gonadotropin-releasing hormone (LHRH). The classic view for hypothalamic control of AP hormone secretion holds that each major HRH modulates the secretion of a single pituitary hormone (1), but there are some notable exceptions to this rule. For example, TRH is acknowledged to be a physiological modulator of both TSH and PRL secretion, just as LHRH controls the release of both LH and FSH. In addition to these acknowledged exceptions, there have been sporadic reports about “paradoxical” responses to various HRHs. Such observations of AP hormone release elicited by a noncorresponding HRH derive from a broad spectrum of studies involving both normal and pathological AP tissue evaluated in vivo or in vitro (2, 3). The reasons underlying these effects are not known, although they frequently are attributed to the lack of proper control mechanisms in established tumor cell lines or in cells derived from pathological specimens.
Quite recently two groups (including our own) have obtained evidence to suggest that “paradoxical” responses to HRHs may be as much the rule as the exception. More specifically, when attempts were made to characterize AP cell phenotypes on the basis of their cytosolic calcium concentration ([Ca2+]i) responses to HRH stimulation, many cells were found to respond to two or more of these agents (4, 5). These findings not only cast doubts on the utility of [Ca2+]i responses as an index of AP cell phenotype, but also raised a couple of interesting questions. Is multi-responsiveness restricted to hormone-producing cells of a given type? Do such pleiotropic [Ca2+]i responses extend to hormone release by living AP cells? In the present study we attempted to address these intriguing questions by combining calcium imaging and immunocytochemistry with the aim of correlating multi-responsiveness to cellular phenotype or by quantifying HRH-induced hormone release from single living cells through the use of reverse hemolytic plaque assay.
MATERIALS AND METHODS
AP cells were obtained from 8- to 10-week-old male Wistar rats and attached to poly-l-lysine-coated glass coverslips as described previously (4). The cells then were either used directly for [Ca2+]i measurements or cultured in RPMI 1640 medium (Sigma) containing 10% fetal calf serum (GIBCO) and antibiotics for 2–3 days. Measurements of [Ca2+]i were performed in cells loaded with fura-2 (6) by digital-imaging fluorescence microscopy. All experiments were conducted at 37°C. The cells were under continuous perfusion with either control or test solutions at 2–3 ml/min. This rate of perfusion allowed >95% exchange of the medium within 5–10 s. The standard control solution had the following composition: 145 mM NaCl, 5 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 10 mM glucose, 10 mM Na+-Hepes, pH 7.4. Test solutions contained in addition either GHRH, LHRH, or CRH at 4 nM or TRH at 100 nM. The cells were epi-illuminated alternately at 340 and 380 nm, and light emitted above 520 nm was recorded by using a Magical Image Processor (Applied Imaging, Newcastle, U.K.). Pixel-by-pixel ratios of consecutive frames obtained at 340 and 380 nm excitation were produced, and [Ca2+]i was estimated from these ratios by comparison with fura-2 standards. Additional details can be found elsewhere (4, 7, 8).
At the end of the [Ca2+]i measurements, AP cells in the microscopic field were typed by immunofluorescence by using antibodies raised against one of the pituitary hormones, either as described previously (9) or in accordance with the abbreviated protocol that follows. Cells were fixed with 4% paraformaldehyde in PBS for 10 min, permeabilized with 0.3% Triton X-100 in the above solution for 3 min, and washed with PBS for 5 min. Then 10% goat serum in PBS was added. After 5 min the antibody against one of the AP hormones labeled with Oregon green 488 (see below) was added and the incubation continued for 15–30 min. After washing, a fluorescence image was captured (excitation, 490 nm; emission, >510 nm) with the image processor. Occasionally, this process was repeated for a second fluorescent antibody against another AP hormone. Then nuclei were stained with Hoechst 33258 (0.5 μg/ml, 10 min), and another fluorescence image was acquired (excitation, 340 nm; emission, >420 nm). The image from the fluorescently stained nuclei facilitated definition of cellular boundaries in cells that were physically close.
Single-cell measurements of PRL release were performed by reverse hemolytic plaque assay in cells prepared from male rats (250 g, Harlan Sprague–Dawley, Holtzman; Madison, WI) as described previously (10). Briefly, AP cells cultured for 2 days on plastic Petri dishes (5 × 105 cells/ml) in DMEM (GIBCO) supplemented with 0.1% BSA, antibiotics, and 10% fetal bovine serum were resuspended by a brief trypsinization, rinsed with DMEM-0.1% BSA, mixed with protein-A-coated ovine erythrocytes, and infused into Cunningham chambers at an AP cell density of 30 cells/μl. After a 1-hr attachment period (37°C; 95% air-5% CO2), chambers were rinsed with assay medium (DMEM-0.1% BSA), flooded with 150 μl of assay medium containing PRL antiserum (1:80) alone or in combination with different treatments (three chambers each), and incubated for the specified times (1 or 2 hr). Guinea pig complement (1:80, 50 min) was added to develop the plaques. After fixation overnight with 2% glutaraldehyde and staining with toluidine blue, the percentage of all pituitary cells that formed PRL plaques was determined after a 1-hr antibody incubation conducted in the absence (control) or presence of 100 nM of each HRH. At least 300 AP cells were counted in each chamber. For measurements of plaque areas, cells were incubated with antibody for 2 hr under the same conditions as before. At this time the percentage of cells forming PRL plaques was maximum as determined in preliminary experiments. The measurements of plaque areas were made by use of a calibrated ocular reticule. The areas of at least 50 plaques/slide were quantified to derive the mean plaque area and frequency distributions of plaque sizes. The data from plaque assays are the result of five (percentage of PRL secretors) and four (plaque areas) independent experiments in which there were three replicates for each treatment. A two-way ANOVA was used to assess statistical significance, and treatment means were compared by use of the Bonnferroni’s Multiple Comparisons Test. Differences were considered to be significant at P < 0.05.
Antisera against rat PRL (rabbit, AFP425–10−91), β-TSH (rabbit, AFP1274789), GH (monkey, AFP4115), β-FSH (guinea pig, AFP85GP9691BFSHB), and anti-human adrenocorticotropin (rabbit, AFP39013082) were generous gifts from the National Hormone and Pituitary Program, the National Institute of Diabetes and Digestive and Kidney Diseases, the National Institute of Child Health and Human Development, and the U.S. Department of Agriculture, Rockville, MD. Fluorescein-labeled anti-rabbit, anti-guinea pig, or anti-monkey IgGs, the hypothalamic-releasing hormones (GHRH, TRH, LHRH, and CRH) and the LHRH antagonist [d-Phe2,6,Pro3]-LHRH all were obtained from Sigma. Fura-2/AM and Oregon green-isothiocyanate were purchased from Molecular Probes.
RESULTS
Many AP Cells Respond to More Than One HRH.
In the experiment illustrated by representative example in Fig. 1, AP cells were consecutively stimulated with each of the four HRHs. Cells A and B responded with an increase in [Ca2+]i to only one of the releasing hormones (TRH and GHRH, respectively). In contrast, cell C responded to three of the HRHs and cell D to all four. The cumulative results obtained in 25 similar experiments with AP cells cultured for 2–3 days (2,133 cells analyzed in total) are summarized in the last column of Table 1. Shown here are the ranges for different combinations of induced responses obtained in three sets of 6–10 experiments carried out over a 2-year period. Consistent with our previous observations (4), these cells (secreting hormone phenotypes not determined) distributed into three subpopulations of roughly equal size. One subgroup was comprised of cells that did not respond to any of the HRHs (nonresponsive cells in Table 1) and accounted for 19–31% of all AP cells. Interestingly, most of the cells in this subgroup (85–95%) exhibited a [Ca2+]i rise after depolarization with high K+ (50 mM) solutions (data not shown). A second subgroup (36–50% of all cells) responded selectively to just one of the HRHs (mono-responsive cells). The final group (29–31%) was made up of cells that responded to more than one HRH (multi-responsive cells) and included some (3–7%) that were responsive to all four. The frequencies found for each of the possible combinations of HRH-responsive cells also are detailed in Table 1. As established in our earlier study (4), cells responding to more than one HRH almost always responded to TRH. Overall, most AP cells were found to be responsive to TRH (53 ± 2%), followed by GHRH (36 ± 1%), CRH (15 ± 2%), and LHRH (14 ± 1%) (mean ± SE of the three sets of experiments referred to above).
Figure 1.
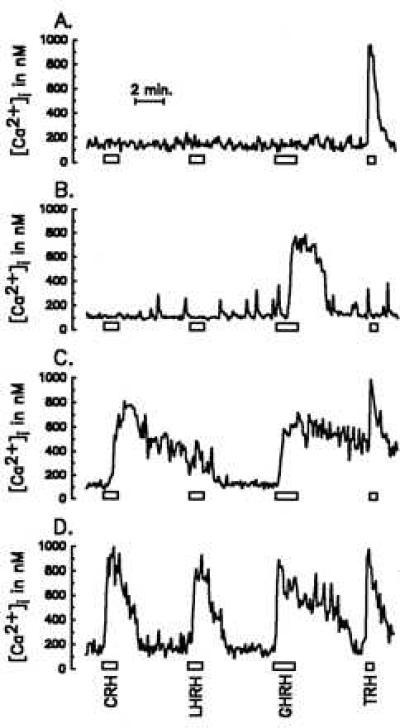
Effects of HRHs on [Ca2+]i in four representative single cells. Cells were stimulated with solutions containing 4 nM CRH, LHRH, or GHRH, and 100 nM TRH, as shown.
Table 1.
Responses of hormone-containing cell types to HRHs
| Cell types | % PRL cells | % GH cells | % ACTH cells | % FSH cells | % TSH cells | % All* cells |
|---|---|---|---|---|---|---|
| Nonresponsive cells | 13 [3] | 13 [9] | 14 | 19 | 9 | 19–31 [22] |
| ΣMono-responsive cells | 28 [66] | 31 [61] | 33 | 33 | 57 | 36–50 [51] |
| CRH | 0 [0] | 0 [1] | 17 | 3 | 0 | 3–7 [3] |
| LHRH | 3 [0] | 3 [4] | 5 | 19 | 9 | 3 [4] |
| GHRH | 0 [1] | 11 [54] | 0 | 0 | 0 | 9–14 [16] |
| TRH | 25 [65] | 17 [2] | 11 | 11 | 48 | 17–30 [28] |
| ΣMultiresponsive cells | 59 [30] | 55 [30] | 54 | 47 | 33 | 29–31 [27] |
| CRH-LHRH | 0 [0] | 0 [0] | 2 | 0 | 0 | <1 [0] |
| CRH-GHRH | 2 [0] | 0 [3] | 6 | 0 | 0 | 0–2 [1] |
| CRH-TRH | 2 [8] | 1 [0] | 14 | 0 | 6 | 2–3 [3] |
| LHRH-GHRH | 0 [0] | 3 [7] | 0 | 3 | 0 | 0–1 [2] |
| LHRH-TRH | 6 [7] | 9 [0] | 0 | 19 | 18 | 1–5 [5] |
| GHRH-TRH | 14 [7] | 23 [13] | 6 | 0 | 0 | 12–16 [8] |
| CRH-LHRH-GHRH | 0 [0] | 0 [3] | 0 | 0 | 0 | <1 [1] |
| CRH-LHRH-TRH | 5 [2] | <1 [0] | 5 | 0 | 3 | 0–3 [2] |
| CRH-GHRH-TRH | 2 [3] | 0 [<1] | 11 | 0 | 0 | 2–3 [1] |
| LHRH-GHRH-TRH | 7 [1] | 6 [2] | 2 | 8 | 3 | 1–3 [2] |
| All four | 23 [2] | 8 [1] | 8 | 17 | 3 | 3–7 [2] |
Proportional abundance of [Ca2+]i responses to HRHs in different AP cell types, identified post facto by the hormone they stored. The different HRH responsiveness inferred from the [Ca2+]i are shown in the first column. Nonresponsive cells indicates no response to any of the HRHs. Numbers correspond to the percentages within each cell type obtained in sets of 3–10 different experiments in AP cells cultured for 2–3 days for cells storing PRL (P, n = 128), GH (n = 79), adrenocorticotropin (ACTH cells, n = 64), FSH (n = 42), or TSH (n = 33). The values in the last column (All cells) correspond to the ranges obtained in three sets of 6–10 experiments, each one (n = 591, 682 and 960 cells) performed at different times during a 2-year period. Cell type was not identified in these experiments. The numbers in brackets correspond to results obtained in 5–7 experiments using freshly prepared AP cells for PRL cells (n = 236), GH cells (n = 200) or all cells without identification (n = 761).
Hormone-secreting phenotype not identified.
Multi-Responsiveness Exists within All Hormone-Containing Cell Types.
There are at least two plausible explanations for multi-responsiveness to HRHs. In one scenario, such apparent indiscrimination might be restricted to hormone-secreting cells of a given type (e.g., lactotropes). Alternatively, several AP cell types might contribute to this phenomenon. In an attempt to distinguish between these possibilities, we immunocytochemically identified AP cell types at the end of the [Ca2+]i measurements, and Fig. 2 illustrates the way in which this was achieved. First, the cells were stimulated sequentially with the four HRHs as before (Fig. 1). Fig. 2A shows three images of [Ca2+]i, coded in gray levels, taken either at rest (CONT.) or during stimulation with GHRH or TRH. At the end of the [Ca2+]i measurements, the cells were fixed and stained with a fluorescent antibody against one of the AP hormones. In routine experiments, only one antibody was used, but in some instances two were used successively (Fig. 2B). As shown by representative example in Fig. 2, one of the cells contained GH and responded only to GHRH, whereas the cell containing PRL responded just to TRH.
Figure 2.
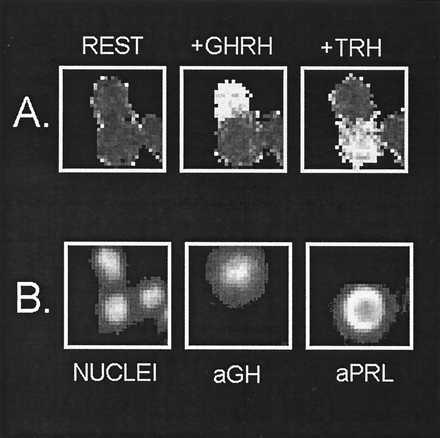
Protocol for combining calcium imaging with immunocytochemical identification of the cell studied. A single microscopic field (30 × 30 μm) containing three single AP cells is shown. (A) Images were captured during the measurement of [Ca2+]i, either before stimulation (REST) or during stimulation with either 4 nM GHRH or 100 nM TRH. Higher [Ca2+]i appears brighter in the gray image. (B) Images were taken during the immunocytochemical identification protocol, after staining with anti-GH-Oregon green (aGH) or after staining with anti-PRL-Oregon green (aPRL). NUCLEI, nuclear localization after staining with Hoescht 33258. See Materials and Methods for details.
Fig. 3 compares the percentages of phenotypically identified AP cells that responded to each of the four HRHs. On the whole, each cell type responded better to its classical, corresponding HRH than it did to the other secretagogues, with the exception of somatotropes where a higher fraction responded to TRH than to GHRH. Surprisingly, multi-responsiveness to HRHs was exhibited by all AP cell types, and TRH along with LHRH was the most likely to induce [Ca2+]i responses regardless of cell phenotype. Among the AP cell types, thyrotropes exhibited the most stringent degree of selectivity for HRHs in that good responses were obtained only with TRH and LHRH.
Figure 3.
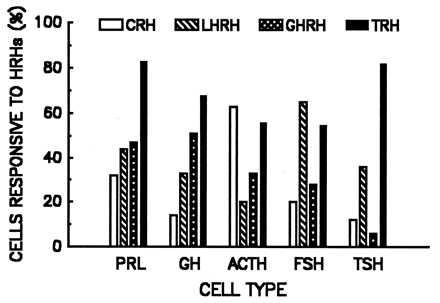
Responses of the AP cell types to each of the HRHs tested. Cells were considered responsive to a given HRH when an increment of [Ca2+]i exceeding 50 nM was obtained in experiments similar to those illustrated in Fig. 1. The number of cells analyzed (3–10 experiments for each cell type) was: PRL, 128; GH, 79; adrenocorticotropin (ACTH), 64; FSH, 42; TRH, 33.
The first five columns in Table 1 provide additional details regarding the combinations of responses exhibited by the different cell types. Cells responding to more than one HRH (multi-responsive cells in Table 1) accounted for roughly half (47–59%) of all lactotropes, somatotropes, corticotropes, and gonadotropes and a smaller proportion of thyrotropes (33%). The percentage of cells responding to all four HRHs was particularly high for lactotropes (23%), negligible for thyrotropes (3%), and intermediate for the other cell types (8–17%). The association of GHRH and TRH responsiveness (particularly the variants GHRH-TRH and LHRH-GHRH-TRH) was quite common in somatotropes and lactotropes. Thyrotropes, on the other hand, were the only cells in which the dominant response was restricted to a single HRH (TRH, 48% of the TSH cells).
Multi-Responsiveness Also Exists with Respect to Hormone Release.
Up to this point, we used changes of [Ca2+]i as a biological end-point with which to assess the effects of HRHs. Unfortunately, such changes may not necessarily culminate in a relevant physiologic response. In pursuit of a more distal biological end-point, we decided to evaluate the effects of HRHs on hormone release from single, living cells by using the reverse hemolytic plaque assay (10). To this end, we focused on a single, representative cell type, the lactotrope, because it exhibited the highest degree of HRH multi-responsiveness in terms of [Ca2+]i changes. Accordingly, we subjected AP cells from male rats to a plaque assay for PRL in the absence or presence of various HRHs, and then measured two relevant parameters. The first of these was the percentage of all AP cells that formed a plaque after a 1-hr incubation with PRL antibody. The rationale here is that a threshold amount of PRL must be released to induce plaque formation. In the case of lactotropes, a 2-hr antibody incubation is required for all cells that release PRL to induce plaque formation. Thus, only a fraction of PRL plaques can be detected after 1 hr under basal conditions, but the addition of HRHs, which accelerate the rate of PRL release, also can accelerate the rate of plaque development. In short, this strategy can be used as a very sensitive assay for identifying regulatory secretagogues, particularly if such agents affect only a subpopulation of cells. The other parameter measured was that of plaque area (after a 2-hr antibody incubation), which provides a reliable index of the cumulative amount of hormone released (10).
The effects of various HRHs on the rate of PRL plaque development is illustrated in Fig. 4A. Clearly, all the HRHs had a quantifiable effect on PRL release, but as anticipated from the [Ca2+]i results, TRH evoked the greatest response. To confirm that the “paradoxical” effect of at least one HRH was mediated by its corresponding receptor, we attempted to block the action of LHRH (100 nM) by concurrent exposure to the LHRH antagonist [d-Phe2,6,Pro3]-LHRH (300 nM). As shown in Fig. 4B, the antagonist diminished the rate of LHRH-induced PRL release by 60%, a value that compares favorably with the 50% reduction of LH release observed (11) when the antagonist was used at an identical molar ratio (3:1). Neither the LHRH antagonist nor somatostatin (a nonspecific peptide control) had any effect on the basal secretion of PRL when treated alone (Fig. 4B). Finally, the percent of plaque-forming cells significantly was increased by all the four HRHs and effect of the different HRHs did not change on varying the dilution of AP cells (Fig. 5). These results suggest that the PRL secretion induced by HRHs was not because of paracrine effects of substances released by cells other than lactotropes, which would be expected to decrease with cell dilution, but to direct action of HRHs on lactotropes. Fig. 6 shows the effects of the four HRHs on the frequency distribution of plaque areas. Each of the four HRHs evoked an increase in the relative amount of PRL release by single cells, as evidenced by the shift to the right of plaque areas in the plots. In all cases, except for TRH, the frequency distribution revealed after stimulation was bimodal, indicating that only a subpopulation of lactotropes was stimulated to release PRL by HRHs other than TRH.
Figure 4.
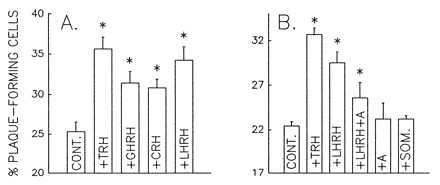
Effects of HRHs on PRL secretion as quantified by reverse hemolytic plaque assay. Cells were incubated concurrently with PRL antibody and the stated HRHs (100 nM) for 60 min. (see Materials and Methods). (A) Effects of HRHs on the percentage of plaque-forming cells. Data are the mean ± SEM of five independent experiments. (B) Inhibition of the LHRH effect by the antagonist [d-Phe2,6,Pro3]-LHRH (+A) at 300 nM; +SOM, effects of somatostatin at 100 nM. Data are representative of two similar experiments. Differences were considered significant (∗) at P < 0.05.
Figure 5.
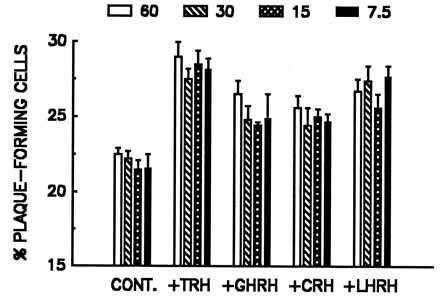
Effects of cell dilution on the secretion of PRL induced by different HRHs. AP cells were infused into the Cunningham chambers at four different dilutions (7.5–60 cells/μl, as shown). Averages of three replicates within a single experiment are shown. All the values obtained with the four different HRHs differed significantly (P < 0.05) from the control values. Profiles are representative of results obtained in two similar experiments. Other details are as in Fig. 4.
Figure 6.
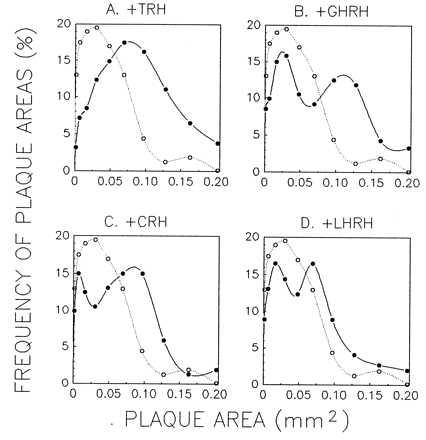
Effects of HRHs on frequency distribution of plaque areas. The sizes of plaques were quantified after a 2-hr antibody incubation with the different HRHs (100 nM). The frequency distributions for control, unstimulated cells (open circles and dotted line) and cells challenged with one of the four HRHs (closed circles and continuous line) are compared. Averages of three replicates within a single experiment are shown. Profiles are representative of results obtained in four similar experiments.
Multi-Responsive Cells Are Not An Artifact of Culture.
It could be argued that AP cells maintained in primary culture for 2–3 days may undergo changes that render them poorly representative of their physiologic condition in the intact animal. In consideration of this possibility, we repeated selected experiments with freshly dispersed cells, which were allowed to attach to poly-l-lysine-coated coverslips for only 1 hr before use. Proportional abundance values for mono- and multi-responsive AP cells from fresh dispersions are presented within brackets in Table 1. Note that the percentages found in the whole population (Table 1, extreme right column) were very similar to those above for cells cultured for 2–3 days. In lactotropes and somatotropes, however, the subpopulations of cells responding to only one HRH was greater in the freshly prepared cells than in their counterparts. This increase in mono-responsive cells was balanced by a decreases in both cells nonresponsive to HRHs and the multi-responsive cells, not just one or the other.
DISCUSSION
According to the classic view for hypothalamic control of AP hormone secretion, each HRH modulates the release of a single AP hormone. It follows logically, therefore, that each AP cell type should be responsive to just one HRH. However, our findings indicate clearly that this is not the case. In fact, when HRH stimulation was assessed by [Ca2+]i dynamics, only one-third of the AP cells were actually responsive to just one HRH, which was not necessarily their classical, corresponding HRH. Another third of the cells did not respond to any HRH, even though stimulation with high-K+ solutions revealed that they were excitable cells, and immunocytochemistry demonstrated that many of them contained AP hormones. More surprisingly, the remaining third of AP cells responded to more than one or all HRHs. Immunocytochemical identification of the stored hormone revealed that multi-responsive cells were not restricted to any particular cell type. Instead, a significant population of multi-responsive cells were present within each of the five AP phenotypes. It could be argued that HRHs might crossreact nonspecifically with several HRH receptors. However, the different sizes and chemical structures of the four HRHs, as well as the low HRH concentrations used in this study, are not consistent with this possibility. It likewise could be argued that multi-responsiveness and/or paradoxical responses to HRHs develop as an artifact of primary culture, and therefore are not reflective of the normal, physiological situation. We have shown, however, that multi-responsive cells were also present in freshly dispersed AP preparations, although their frequencies were somewhat lower than in those in cultures maintained for 2–3 days. The possibility that such seemingly “inappropriate” responses actually derive from activation of “appropriate” receptors is supported by recent reports that roughly half of the AP mRNA for the TRH receptor is localized to somatotropes (12) and that the mRNA encoding the receptor for somatostatin (the classic hypothalamic GH inhibiting factor) is distributed among all five major cell types of the AP (13). In addition, it has been reported recently that 38% percent of somatotropes bind biotinylated GnRH, demonstrating that a representative subpopulation of GH cells bears GnRH receptors (14). When viewed as a whole, our results strongly support the view that multi-responsiveness to hypothalamic hormones is a genuine property of normal AP cells, and not just an anomaly restricted to pathophysiological conditions.
Our present data demonstrate that a variety of HRHs can increase [Ca2+]i within a given hormone-secreting cell type. Inasmuch as [Ca2+]i is believed to mediate the secretory actions of HRHs (15–17), it seemed reasonable to propose that multiple HRHs should converge on a single AP cell to modulate the release of each hormone. We decided to test this line of reasoning on lactotropes, the AP cell type that exhibited the largest fraction of multi-responsive cells. By using the reverse hemolytic plaque assay, we found that all four HRHs increased both the rate and the cumulative amount of PRL released from single, living lactotropes. Analysis of frequency distributions of plaque areas revealed that TRH stimulated most of the lactotropes to release PRL, whereas the other three HRHs induced PRL export from just select subpopulations of cells. Thus the effects of the “appropriate” HRHs were more universal than those of the “inappropriate” counterparts, although the latter were clearly unequivocal. As was the case for [Ca2+]i mobilization, the effects of HRHs on PRL secretion appeared to be because of specific stimulation of corresponding HRH receptors on lactotropes, because they were not reproduced by peptides of comparable sizes and structures and the effects of LHRH were inhibited by a specific LHRH antagonist (Fig. 4). It could be argued that paracrine effects could contribute to stimulation by noncorresponding HRHs. Although this possibility cannot be absolutely ruled out, the results from the cell dilution experiments (Fig. 5) suggest that this mechanism does not play a major role. The finding that multi-responsive cells were not restricted to a given hormone-secreting cell type suggests that secretion of AP hormones other than PRL also may be controlled in a multi-factorial manner.
The results of the present study raise an interesting and provocative question: If multi-responsiveness of AP cells is as much the rule as the exception, then why have others not reported it previously? In fact, there have been numerous reports entirely consistent with this possibility, but interpretation of those findings appears to have been influenced by the conventional wisdom of the time. For example, it has been reported that GHRH is capable of eliciting secretion of LH and FSH from rat anterior pituitaries in vitro (18), and that TRH is almost as potent as LHRH in evoking LH release (19). In addition, TRH has been proposed to be a GH-releasing factor (2) on the basis of the ability to induce GH release by pituitary cells from rats and other mammals (20). There also have been reports that LHRH can stimulate secretion of GH and TSH from AP cell cultures (21) as well as PRL release from aggregate AP cells (22). Reports about so-called “paradoxical” responses to HRHs are likewise quite common for healthy and infirm human subjects (2), but such observations are not fully interpretable, owing to the potential for indirect effects of HRHs administered in vivo. In summary, our findings that secretion of a given AP hormone can be regulated directly by several HRHs are not isolated; the results of the numerous studies by others support the same conclusion.
The multi-responsive nature of AP cells invites speculation that HRHs subserve regulatory roles that transcend hormonal synthesis, release, and mitotic rates within their traditional target cells. One possibility is that in addition to regulating the acute release of its corresponding hormone, a given HRH could exert a chronic influence on the proportion of cells that secrete a different hormone. In this manner, changes in physiologic requirements for the product of a particular cell type could be met without a net increase in the size of the AP gland. An example of such phenotypic plasticity is the functional interconversion (i.e., transdifferentiation) of mammotropes and somatotropes that is induced by LHRH (see ref. 23 for a review). A variation on this theme is that cells with “inappropriate” receptors might comprise a reserve population, which, when activated by the “inappropriate” HRH, can be recruited to secrete the corresponding hormone. This possibility is predicated on the notion that established AP cell types contain trace amounts of “inappropriate” hormones, and reports about immunocytochemically identified gonadothyrotropes and somatogonadotropes (14) along with corticotropes that co-stain for LH, FSH, TSH, or PRL (24) are certainly consistent with this idea. A final possibility worthy of consideration is that combinations of two or more HRHs might comprise codes that are deciphered by target cells to effect a specific response. Such a modulated response might take the form, for example, of the induction of mono-hormonal secretion by a bi-hormonal cell. The specific biological endpoint notwithstanding, it is clear that hypothalamic control of AP function is considerably more complicated than previously envisioned.
Acknowledgments
Financial support from the Spanish Dirección General de Investigación Científica y Técnica (DGICYT, Grant PB92-0268) and from the National Institutes of Health (Grant DK38251) is gratefully acknowledged.
Footnotes
This paper was submitted directly (Track II) to the Proceedings Office.
Abbreviations: AP, anterior pituitary; [Ca2+]i, cytosolic calcium concentration; HRH, hypothalamic-releasing hormone; PRL, prolactin; GH, growth hormone; TSH, thyrotropin; FSH, follicle-stimulating hormone; LH, luteinizing hormone; TRH, TSH-releasing hormone; GHRH, GH-releasing hormone; CRH, corticotropin-releasing hormone; LHRH, luteinizing hormone-releasing hormone or gonadotropin-releasing hormone.
References
- 1.Harris G W, Reed M, Fawcett C P. Br Med Bull. 1966;196:266–272. doi: 10.1093/oxfordjournals.bmb.a070485. [DOI] [PubMed] [Google Scholar]
- 2.Harvey S. J Endocrinol. 1990;125:345–358. doi: 10.1677/joe.0.1250345. [DOI] [PubMed] [Google Scholar]
- 3.Amsterdam J D, Winokur A, Lucki I, Snyder P, Harris R I, Caroff S, Rickels K. Psychoneuroendocrinology. 1982;7:177–184. doi: 10.1016/0306-4530(82)90010-5. [DOI] [PubMed] [Google Scholar]
- 4.Villalobos C, Núñez L, García-Sancho J. FASEB J. 1996;10:654–660. doi: 10.1096/fasebj.10.5.8621065. [DOI] [PubMed] [Google Scholar]
- 5.Kasahara K, Tasaka K, Masumoto N, Mizuki J, Tahara M, Miyake A, Tanizawa O. Biochem Biophys Res Commun. 1994;199:1436–1441. doi: 10.1006/bbrc.1994.1391. [DOI] [PubMed] [Google Scholar]
- 6.Grynkiewicz G, Poenie M, Tsien R Y. J Biol Chem. 1985;260:3440–3450. [PubMed] [Google Scholar]
- 7.Villalobos C, Fonteriz R, López M G, García A G, García-Sancho J. FASEB J. 1992;6:2742–2747. doi: 10.1096/fasebj.6.9.1319362. [DOI] [PubMed] [Google Scholar]
- 8.López M G, García A G, Artalejo A R, Neher E, García-Sancho J. J Physiol (London) 1995;482:15–27. doi: 10.1113/jphysiol.1995.sp020496. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Núñez L, De la Fuente M T, Garcia A G, García-Sancho J. Am J Physiol. 1995;269:C1540–C1546. doi: 10.1152/ajpcell.1995.269.6.C1540. [DOI] [PubMed] [Google Scholar]
- 10.Boockfor F R, Hoeffler J P, Frawley L S. Neuroendocrinology. 1986;42:64–70. doi: 10.1159/000124250. [DOI] [PubMed] [Google Scholar]
- 11.Bowers C Y, Humphries J, Wasiak T, Folkers K, Reynolds G A, Reichert L E., Jr Endocrinology. 1980;106:674–683. doi: 10.1210/endo-106-3-674. [DOI] [PubMed] [Google Scholar]
- 12.Konaka S, Yamada M, Satoh T, Ozawa H, Watanabe E, Takata K, Mori M. Endocrinology. 1997;138:827–830. doi: 10.1210/endo.138.2.5030. [DOI] [PubMed] [Google Scholar]
- 13.O’Carroll A-M, Krempels K. Endocrinology. 1995;136:5224–5227. doi: 10.1210/endo.136.11.7588261. [DOI] [PubMed] [Google Scholar]
- 14.Childs G V, Miller B T, Miller W L. Endocrinology. 1997;138:1577–1584. doi: 10.1210/endo.138.4.5090. [DOI] [PubMed] [Google Scholar]
- 15.Gershengorn M C. Annu Rev Physiol. 1986;48:515–526. doi: 10.1146/annurev.ph.48.030186.002503. [DOI] [PubMed] [Google Scholar]
- 16.Kato M, Hoyland J, Sikdar S K, Mason W T. J Physiol (London) 1992;447:171–189. doi: 10.1113/jphysiol.1992.sp018997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Shangold G A, Murphy S N, Miller R J. Proc Natl Acad Sci USA. 1988;83:6566–6570. doi: 10.1073/pnas.85.17.6566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Knepel W, Schwaninger M, Wesemeyer K D, Döhler K D, Sandow J. Endocrinology. 1987;120:732–738. doi: 10.1210/endo-120-2-732. [DOI] [PubMed] [Google Scholar]
- 19.Fujihara N, Shiinol M. Can J Physiol Pharmacol. 1983;61:186–189. doi: 10.1139/y83-029. [DOI] [PubMed] [Google Scholar]
- 20.Welsh J B, Cuttler L, Szabo M. Endocrinology. 1986;119:2368–2375. doi: 10.1210/endo-119-5-2368. [DOI] [PubMed] [Google Scholar]
- 21.Ghosh B R, Wu J C, Strahl B D, Childs G V, Miller W L. Endocrinology. 1996;137:5144–5154. doi: 10.1210/endo.137.11.8895389. [DOI] [PubMed] [Google Scholar]
- 22.Robberecht W, Andries M, Denef C. Neuroendocrinology. 1992;56:185–194. doi: 10.1159/000126227. [DOI] [PubMed] [Google Scholar]
- 23.Frawley L S, Boockfor F R. Endocr Rev. 1991;12:337–355. doi: 10.1210/edrv-12-4-337. [DOI] [PubMed] [Google Scholar]
- 24.Childs G V. Trends Endocrinol Metab. 1991;2:112–117. doi: 10.1016/s1043-2760(05)80007-4. [DOI] [PubMed] [Google Scholar]