

Abstract
In angiosperms, the functional enucleate sieve tube system of the phloem appears to be maintained by the surrounding companion cells. In this study, we tested the hypothesis that polypeptides present within the phloem sap traffic cell to cell from the companion cells, where they are synthesized, into the sieve tube via plasmodesmata. Coinjection of fluorescently labeled dextrans along with size-fractionated Cucurbita maxima phloem proteins, ranging in size from 10 to 200 kDa, as well as injection of individual fluorescently labeled phloem proteins, provided unambiguous evidence that these proteins have the capacity to interact with mesophyll plasmodesmata in cucurbit cotyledons to induce an increase in size exclusion limit and traffic cell to cell. Plasmodesmal size exclusion limit increased to greater than 20 kDa, but less than 40 kDa, irrespective of the size of the injected protein, indicating that partial protein unfolding may be a requirement for transport. A threshold concentration in the 20–100 nM range was required for cell-to-cell transport indicating that phloem proteins have a high affinity for the mesophyll plasmodesmal binding site(s). Parallel experiments with glutaredoxin and cystatin, phloem sap proteins from Ricinus communis, established that these proteins can also traffic through cucurbit mesophyll plasmodesmata. These results are discussed in terms of the requirements for regulated protein trafficking between companion cells and the sieve tube system. As the threshold value for plasmodesmal transport of phloem sap proteins falls within the same range as many plant hormones, the possibility is discussed that some of these proteins may act as long-distance signaling molecules.
The phloem system plays a pivotal role in the delivery of resources (photosynthate, amino acids, and mineral nutrients) to heterotrophic plant tissues. In angiosperms, the phloem is comprised of sieve tube members and their associated companion cells. During differentiation, each sieve tube member enters a highly specialized developmental program in which the vacuole is removed, the plastids become highly modified and reduced in number, the nucleus degenerates, ribosomes are reduced (or absent), and, although retained, the endoplasmic reticulum is confined to a parietal position in association with the plasma membrane (1). As part of this maturation process, plasmodesmata interconnecting individual sieve tube members undergo structural modifications to produce open plasma membrane-lined pores within developing sieve plates and lateral sieve areas. Subsequently, a pressure gradient within the mature, functional, sieve tube causes a time-dependent volume displacement of the contents of individual sieve tube members in the direction of this gradient.
The functionality of the higher plant phloem sieve tube system is highly dependent on the maintenance of membrane integrity; loss of ability to establish osmotic gradients would quickly abolish the capacity for translocation. As individual enucleate sieve tubes appear to retain functional status for weeks to many months, it would seem logical to assume that, during the evolution of this advanced sieve tube system, aspects of cellular maintenance were transferred to neighboring cells (2). As individual sieve tube members remain symplasmically connected to companion cells via specialized plasmodesmata, these cells were likely recruited to perform such maintenance functions (3). As protein and lipid turnover would be involved in this process, this evolutionary step most likely required that plasmodesmata, linking companion cells to sieve tube members, developed the capacity to mediate the trafficking of specialized proteins (4).
Until quite recently, plasmodesmata were thought only to establish a symplasmic pathway for the cell-to-cell exchange (by diffusion) of small molecules, such as biochemical intermediates, photosynthates, amino acids, and ions. However, based on pioneering studies performed by using viral-encoded movement proteins (MPs; refs. 5–9), and more recently with plant proteins (10, 11), it is now clear that higher plant plasmodesmata have evolved the capacity to engage in the transport of macromolecules (proteins and nucleic acids). Direct microinjection experiments, performed with viral MPs and plant proteins, have established that cell-to-cell trafficking of macromolecules involves (i) protein-mediated increase in plasmodesmal size exclusion limit (SEL), (ii) protein transport, and (iii) protein-mediated trafficking of RNA/DNA (9, 12). Thus, there is now an experimental foundation for the hypothesis that either protein or mRNA, expressed in one cell or group of cells (e.g., the phloem companion cells), can exert control over developmental and/or physiological processes within surrounding cells (e.g., sieve tube members), via selective plasmodesmal-mediated transport (11).
Additional evidence in support of the hypothesis that protein trafficking occurs between companion cells and sieve tube members has been provided by studies performed on the phloem sap collected from a number of plant species. For example, Fisher et al. (13) established that wheat phloem sap, collected from excised aphid stylets, contained more than 200 proteins that were present within the translocation stream. Analysis of this wheat phloem sap indicated that the majority of these proteins fell within the size range of 10–40 kDa, but a number of larger proteins (60–70 kDa) were also detected. The presence of a similar range of proteins has now been established for the phloem sap of rice (14, 15) and Ricinus (16, 17). It is also important to note that, in the studies performed on wheat, it was clear that a constant exchange of proteins between the sieve tube member and the companion cell occurred along the translocation pathway. This finding raised the possibility that some of the proteins present in the phloem sap may be components of a long-distance signaling mechanism involved in coordinating various physiological processes between plant organs (18, 19).
In the present study, we provide direct experimental evidence that proteins present within the phloem sap of Cucurbita maxima and Ricinus communis have the capacity to interact with plasmodesmata to (i) induce a significant increase in SEL (greater than 20 but less than 40 kDa) and (ii) mediate their own cell-to-cell transport. Our microinjection experiments also revealed that phloem proteins ranging from 10 to 200 kDa induced an increase in SEL to the same extent, suggesting that protein unfolding may be an essential step in plasmodesmal trafficking. In addition, we report on the threshold level (20–100 nM) of protein required to initiate transport through plasmodesmata. These results are discussed in terms of the role of plasmodesmata in phloem and plant function.
MATERIALS AND METHODS
Plant Material.
C. maxima Duch. cv Big Max plants were grown during the summer in a pathogen-free greenhouse under natural daylight conditions. Plants for use in microinjection experiments were grown under controlled environment conditions.
Collection and Fractionation of Cucurbit Phloem Proteins.
Phloem exudate (sap) was obtained from cut stems of 2-month-old cucurbit plants. Microcapillary pipettes were used to collect the phloem exudate that was then transferred to a Tris buffer (50 mM, pH 7.5) solution containing 0.5% (vol/vol) 2-mercaptoethanol. Phloem sap was then dialyzed for 2–3 h against this same buffer before being stored at −80°C. Dialyzed cucurbit phloem sap proteins were separated by SDS/PAGE and stained with Coomassie brilliant blue. Gels were then dissected to yield size-fractionated proteins that were as follows: fraction I, 10–20 kDa; fraction II, 20–25 kDa; fraction III, 25–50 kDa; fraction IV, 50–80 kDa; fraction V, 80–100 kDa; and fraction VI, 100–200 kDa. Proteins from each gel slice were initially electroeluted into 50 mM ammonium bicarbonate containing 0.1% (wt/vol) SDS (for 2 h) by using the protocol provided by Bio-Rad. The elution buffer was then changed to 50 mM ammonium bicarbonate and the proteins electroeluted for a further 3–5 h. Eluted proteins were then concentrated by partial lyophilization or by vacuum evaporation at 4°C, to yield final stock concentrations ranging from 0.1–0.5 μg⋅μl−1. In addition, individual protein bands having apparent molecular weights of 17, 24, 30, 40, and 52 kDa were cut from SDS/PAGE gels and protein electroeluted and concentrated as described above. Note that, as previously established by Bostwick et al. (20), the 24-kDa band was shown by Western bot analysis to contain phloem protein II (PP2). In addition, a polyclonal antibody raised against glutaredoxin from Ricinus phloem sap (21) cross-reacted with the 52-kDa protein, suggesting that these two proteins may be related.
Finally, for some experiments, PP2 was purified directly from the cucurbit phloem exudate by using the ovomucoid-acrylic bead method described by Bostwick et al. (20).
Escherichia coli Expression and Labeling of Proteins for Use in Microinjection Studies.
Different E. coli strains were employed to express a range of proteins used in microinjection experiments. The PP2 ORF was cloned into a pRSET-B plasmid vector and transformed into E. coli BL21. PP2 expression (as an N-terminal histidine-tagged fusion protein), purification, and renaturation for microinjection experiments were as described by Noueiry et al. (6). PP2 was also expressed in E. coli strain JM109 by using M13 phage according to the manufacturer’s instructions (Invitrogen). Routinely, a Ni-NTA agarose column (Qiagen, Chatsworth, CA) was used to purify the histidine-tagged PP2. Removal of the N-terminal histidine-tag was achieved, with the enterokinase cleavage site on the fusion protein, by first equilibrating the PP2 bound to the Ni-NTA column with a 50 mM Tris buffer (pH 8) solution containing 1 mM CaCl2 and 0.1% (vol/vol) Tween 20 followed by digestion with EKMax (Invitrogen) at 25°C for 20 h. The cleaved PP2 was eluted with 0.1 M NaHCO3, pH 9 and 2% (wt/vol) SDS, and cleavage was confirmed by SDS/PAGE. This PP2 was then prepared for microinjection by using either the procedures described by Noueiry et al. (6) or the above-described protocols developed for fractionation of phloem proteins.
Two mutant forms of PP2 were also expressed in E. coli strain BL21. The strategy for subcloning was as follows: cPC13 was digested with either Sst/EcoRI or XhoI/NcoI to yield fragments coding an N-terminal 77 amino acid and a C-terminal 40 amino acid PP2 deletion mutant, respectively.
The ORFs of glutaredoxin (21) and cystatin (J. Szederkényi and C.S., unpublished results), two R. communis phloem sap proteins, were cloned into the BamHI/HindIII sites of pQE30 (Qiagen) and expressed in M15 cells following the manufacturer’s instructions.
Two E. coli strains carrying either the Arabidopsis thaliana ubiquitin coding region or a ubiquitin deletion mutant, which lacked the four C-terminal residues, in pET3c (Novagen), were kindly provided by Judy Callis (University of California, Davis). Expression of ubiquitin was carried out by using the protocols of Beers and Callis (22). The histidine-tagged proteins were purified on a Ni-NTA agarose columns as previously described.
The maize homeobox protein, KNOTTED1 (KN1; wild-type and dysfunctional mutant), and the 3a MP of cucumber mosaic virus (CMV-MP) were similarly expressed in E. coli (7, 10) for use in control microinjection experiments.
As only limited quantities of purified cucurbit phloem sap proteins could be prepared, it was not feasible to label all proteins with fluorescein isothiocyanate (FITC) for microinjection experiments. Thus, for initial experiments, size-fractionated proteins obtained from the cucurbit phloem sap were mixed (1:1 volumes) with a 2 mM FITC-labeled dextran solution (10, 20, or 40 kDa; Molecular Probes) prepared in 5 mM KHCO3. All proteins obtained by using the various E. coli expression systems were fluorescently labeled with FITC according to the methods outlined by Noueiry et al. (6). In addition, for SEL experiments, unlabeled protein was mixed with FITC-labeled dextrans, as described above. Lucifer yellow CH (LYCH, Sigma) was prepared as a 1 mM solution in 5 mM KHCO3. Immediately prior to use in microinjection experiments, all solutions were filtered through a 0.1-μm pore polyvinylidene difluoride membrane filter (Ultrafree C3 VV; Millipore) and then stored on ice until used. Fluorescent probes (FITC-dextrans) and fluorescently labeled proteins used in microinjection experiments were between 1–2 mM and 1–5 μg⋅μl−1 (unless otherwise noted), respectively.
Microinjection Procedures.
Cotyledons of 10- to 14-day-old cucurbit plants were used for all microinjection studies, with a minimum of five plants being used per protein tested. Details regarding the instrumentation and procedures for pressure-mediated microinjection and movement of probes were as described (23).
RESULTS
Cucurbit Phloem Sap Proteins Can Interact with Mesophyll Plasmodesmata.
Analysis of the cucurbit phloem exudate revealed that it contained a large number of proteins ranging in molecular weight from ≈10 to 200 kDa (Fig. 1). The relative concentrations of proteins present within the cucurbit phloem sap covered a wide range. However, the pattern and relative abundance of these proteins was found to be very stable from plant to plant. Major protein bands at 24 kDa and 90 kDa represent PP2 and phloem protein I (PP1), respectively. Given that such a large number of proteins are present in the long-distance translocation stream, they must either enter the phloem during the process of sieve tube maturation, or traffic, via plasmodesmata, from the site of synthesis within the surrounding companion cells into the sieve tube system.
Figure 1.
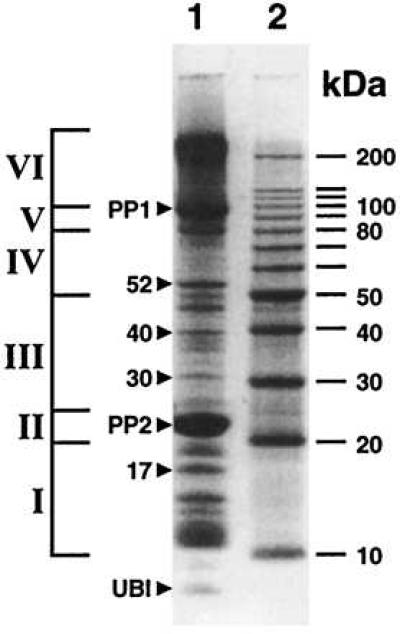
Phloem exudate collected from the stem of 2-month-old C. maxima plants for use in microinjection studies. Shown is the protein pattern obtained by SDS/PAGE (lane 1). Lane 2 displays the molecular weight markers. Distribution of phloem proteins present in size-fractionated samples are evident: fraction I, 10–20 kDa; fraction II, 20–25 kDa; fraction III, 25–50 kDa; fraction IV, 50–80 kDa; fraction V, 80–100 kDa; fraction VI, 100–200 kDa. Individual proteins used in microinjection experiments are indicated by darts. UBI, ubiquitin.
To test the plasmodesmal trafficking hypothesis, the cucurbit phloem sap proteins were size fractionated and then employed directly in microinjection experiments at concentrations approximating those present in the phloem exudate. Earlier studies had established that the companion cell-sieve tube system could be accessed by microinjection methods (24, 25). However, initial experiments performed on petiole and stem tissues established the very low efficacy of this approach (see also refs. 25 and 26). This was particularly problematic given the large number of protein samples to be employed in the present study. Thus, we were forced to seek an alternative experimental tissue on which to perform these phloem sap protein experiments. In earlier experiments conducted in our laboratory we demonstrated that KN1, a homeobox protein that functions in the meristems of Zea mays (10, 27), could traffic through plasmodesmata located in other maize tissues (mesophyll and bundle sheath) as well as through plasmodesmata in heterologous tissues (tobacco mesophyll) (10). In view of these results, we focused the present studies on mesophyll cells located within the cotyledons of C. maxima.
Coinjection of fraction I proteins and 10-kDa FITC-dextran resulted in the movement of fluorescence from the injected mesophyll cell into the surrounding tissue. Experiments performed with 20-kDa FITC-dextran yielded identical results, whereas when the cucurbit fraction I proteins were coinjected with 40-kDa FITC-dextran the associated fluorescence remained within the injected cell (Table 1). In control experiments in which 10-, 20-, or 40-kDa FITC-dextran was injected alone, fluorescence was confined to the target mesophyll cell (data not shown). These experiments were repeated with fraction II proteins, which was largely comprised of PP2. As illustrated in Table 1, these cucurbit phloem proteins also mediated the cell-to-cell movement of both the 10- and 20-kDa FITC-dextrans, but, as with fraction I proteins, the 40-kDa FITC-dextran was confined to the target cell. In coinjection experiments with both fraction I and II proteins, fluorescence was detected in at least 10 cells within 1–2 min. Thus, protein(s) within each fraction must have the capacity to interact with mesophyll plasmodesmata to induce an increase in SEL and mediate its (their) cell-to-cell transport.
Table 1.
Ability of cucurbit phloem sap proteins to increase plasmodesmal SEL and traffic cell to cell
| Phloem exudate proteins
|
Microinjections*
|
||||
|---|---|---|---|---|---|
| Fraction no. | Size range, kDa | Concentration,† μg⋅μl−1 | 10-kDa FITC-dextran | 20-kDa FITC-dextran | 40-kDa FITC-dextran |
| I | 10–20 | 0.05 | 11 (15; 73%) | 11 (15; 73%) | 0 (10; 0%) |
| II | 20–25 | 0.05 | 13 (16; 81%) | 12 (15; 80%) | 0 (10; 0%) |
| III | 25–50 | 0.05 | 11 (14; 79%) | 10 (14; 71%) | 0 (11; 0%) |
| IV | 50–80 | 0.25 | 10 (14; 71%) | 11 (14; 79%) | 1 (10; 10%) |
| V | 80–100 | 0.10 | 10 (15; 67%) | 11 (14; 79%) | 0 (12; 0%) |
| VI | ≥100 | 0.05 | 11 (14; 79%) | 10 (15; 67%) | 2 (12; 17%) |
Size-fractionated proteins and FITC-dextrans were coinjected into mesophyll cells within the cotyledons of C. maxima plants. Control injections performed with LYCH established that the plasmodesmata, within the injection site, permitted rapid and extensive movement of this small membrane-impermeable probe.
Number of injections in which the fluorescently labeled probe moved from the injected cell into the surrounding tissue (total number of injections in each experiment and percent movement given in parentheses). Fluorescence associated with the injected FITC-dextran began to move into neighboring cells upon delivery into the target cell and, within 1-2 min, it had spread to at least 10 cells.
Represents phloem sap protein concentration back-loaded into the microinjection pipette.
Microinjection experiments performed on fraction III (27–50 kDa), IV (50–80 kDa), V (80–100 kDa), and VI (≥100 kDa) proteins established that the capacity to induce an increase in plasmodesmal SEL was a common feature of all protein fractions extracted from the cucurbit phloem. The microinjection data presented in Table 1 revealed that the phloem proteins, present within each fraction, increased the mesophyll plasmodesmal SEL to a value greater than 20 kDa but less than 40 kDa. Here it is important to note that the detected increase in SEL was not correlated with the actual size of the proteins present within the fractions; i.e., 10–20 kDa through to 200-kDa proteins had the same effect on plasmodesmal SEL. Furthermore, cell-to-cell movement of phloem proteins was detected even though the protein level within the injection pipette was between 0.05 to 0.25 μg⋅μl−1 (Table 1). This protein level is an order of magnitude lower than that used in our previous studies on viral MPs and KN1. Hence, these microinjection experiments provided direct experimental support for the hypothesis that numerous (all?) proteins present within the phloem sap have the ability, when present at low concentrations, to interact with mesophyll plasmodesmata to increase SEL and traffic cell to cell.
To ascertain the ability of individual phloem proteins to potentiate their cell-to-cell transport, we next performed experiments on protein fractions obtained by cutting specific protein bands from the SDS/PAGE gels. For these studies we chose proteins reflecting a range of levels, from high to medium abundance (see Fig. 1). Coinjection of the 17-, 24-, 30-, and 52-kDa proteins with 20-kDa FITC-dextran resulted in cell-to-cell movement that was identical in nature to that observed with fraction I–VI proteins (see Tables 1 and 2). Although the 40-kDa protein band moved in some injection experiments, the frequency (31%) and extent of this movement (2–5 cells) was clearly below that observed for the other four proteins examined (frequencies, 70–90%; movement, 10–20 cells).
Table 2.
Capacity of specific cucurbit phloem sap proteins to interact with plasmodesmata to induce an increase in SEL and mediate their own cell-to-cell transport in cotyledons of C. maxima
| Injected material | Protein concentration,* μg⋅μl−1 | Microinjections† |
|---|---|---|
| LYCH | — | 15 (15; 100%) |
| 20-kDa FITC-dextran | — | 0 (15; 0%) |
| 17-kDa protein fraction‡ | 0.6 | 12 (17; 71%) |
| 24-kDa protein fraction | 1.0 | 14 (15; 93%) |
| 30-kDa protein fraction | 1.5 | 11 (14; 79%) |
| 40-kDa protein fraction | 1.4 | 5 (16; 31%) |
| 52-kDa protein fraction | 1.2 | 12 (15; 80%) |
Represents phloem sap protein concentration back-loaded into the microinjection pipette.
Details as in Table 1.
Each protein fraction was coinjected with 20-kDa FITC-dextran.
Comparative Studies on Phloem Sap and E. coli-Expressed PP2.
The PP2 coding sequence (21) with an N-terminal histidine-tag was transformed into E. coli to permit protein expression, purification, FITC-labeling, and renaturation of PP2. This material was then used to perform comparative studies on the manner in which E. coli-expressed and phloem-derived PP2 interact with mesophyll plasmodesmata. As a control for these experiments, parallel microinjection studies were carried out by using the 3a MP of cucumber mosaic virus (CMV-MP) and KN1. The CMV-MP was chosen because C. maxima is a natural host for CMV, and thus its MP should be well adapted for cell-to-cell transport through cotyledonary mesophyll plasmodesmata. The rationale for using KN1 was that this plant transcription factor has been shown to be capable of moving in heterologous tissues from a range of plant species, including Z. mays and Nicotiana tabacum (10), and Nicotiana benthamiana, Vigna unguiculata (cowpea), and Antirrhinum majus (snapdragon) (S.B. and W.J.L., unpublished results). To allow direct comparisons between experiments, all injected proteins were prepared and back-loaded into the micropipette at a concentration of 1.0–2.0 μg⋅μl−1.
Microinjection experiments performed with FITC-CMV-MP revealed that this viral MP has the capacity to mediate its cell-to-cell transport through mesophyll plasmodesmata in cucurbit cotyledons; similar results were obtained with FITC-KN1 (Table 3). Note that, as in our previous study (10), the mutant form (MT6) of KN1 was unable to move from the injected cell (on the one occasion when fluorescence was observed to move, it was confined to two adjacent cells). These control experiments established that FITC-CMV-MP and FITC-KN1 behave in an identical manner (very similar efficacy of movement from the injected cell as well as extent of cell-to-cell movement) in the experimental tissue chosen for cucurbit phloem protein injection studies.
Table 3.
Cell-to-cell movement capacity of cucurbit phloem sap and E. coli-expressed PP2
| Injected material | Microinjections* | Extent of movement |
|---|---|---|
| LYCH | 59 (63; 94%) | 20–30 cells |
| 20-kDa FD | 5 (57; 9%) | 1–4 cells† |
| FITC-CMV-MP | 15 (22; 68%) | 20–30 cells |
| FITC-KN1 | 14 (19; 74%) | 20–30 cells |
| FITC-KN1 (MT6)‡ | 1 (10; 10%) | 2 cells† |
| FITC-PP2 (PS)§ | 22 (28; 79%) | 20–30 cells |
| FITC-PP2 (Ec)§ | 14 (19; 74%) | 20–30 cells |
| PP2 (PS) + 20-kDa FD | 20 (26; 77%) | 20–30 cells |
| PP2 (Ec) + 20-kDa FD | 15 (18; 83%) | 20–30 cells |
| PP2 (PS)¶ + 20-kDa FD | 8 (10; 80%) | 20–30 cells |
| PP2 (PS) + 40-kDa FD | 0 (14; 0%) | 0 cells |
| PP2 (Ec) + 40-kDa FD | 2 (15; 13%) | 2–5 cells† |
| FITC-PP2 (MT1)‡ (Ec) | 7 (13; 54%) | 10–12 cells |
| PP2 (MT1) (Ec) + 20 kDa FD | 6 (13; 46%) | 10–12 cells |
| PP2 (MT1) (Ec) + 40 kDa FD | 0 (10; 0%) | 0 cells |
| PP2 (MT2)‡ (Ec) | 9 (14; 64%) | 20–30 cells |
| PP2 (MT2) (Ec) + 20 kDa FD | 22 (30; 73%) | 20–30 cells |
| PP2 (MT2) (Ec) + 40 kDa FD | 0 (11; 0%) | 0 cells |
In these experiments either FITC-labeled proteins or unlabeled proteins plus FITC-dextrans (FD) were pressure-injected into mesophyll cells within the cotyledons of C. maxima plants. Control experiments included the CMV-MP and KN1 from Z. mays.
Details as in Table 1. Proteins prepared from E. coli (CMV-MP, KN1, and PP2) were back-loaded into the pipette tip at concentrations from 1.5 to 2.0 μg⋅μl−1. PP2 (PS) was used at 1.0 μg⋅μl−1.
When movement of fluorescence was detected, it was usually confined to adjacent cells.
KN1 (MT6) is described in ref. 10. PP2 MT1 and PP2 MT2, proteins in which the N-terminal 77 amino acids and C-terminal 40 amino acid residues were deleted, respectively.
(PS) and (Ec) refer to proteins obtained from cucurbit phloem sap and expressed in E. coli, respectively.
PP2 purified from PS using ovomucoid-acrylic beads was used at a concentration of 0.05 μg⋅μl−1.
As illustrated by the data presented in Table 3, FITC-PP2 prepared from either cucurbit phloem sap or from the E. coli expression system reacted in a similar manner in microinjection experiments. In addition, these parallel studies established that the capacity of PP2 to induce an increase in plasmodesmal SEL was the same, irrespective of the source of this protein. Furthermore, experiments performed with PP2 purified directly from the cucurbit phloem exudate, by using the ovomucoid-acrylic bead method (20), established that cell-to-cell movement of this PP2 preparation was comparable to that observed with PP2 that was electroeluted from SDS/PAGE gels. This result is very significant, as it resolved our concern that PP2 treatment with SDS, or the presence of residual levels of SDS within the PP2 preparations, could induce artifacts in the microinjection experiments. Finally, a comparison between the phloem protein fraction II data presented in Table 1 and the results obtained by using FITC-labeled PP2, collected from the phloem sap, indicated that the FITC-dextran coinjection experiments accurately report on the capacity of protein trafficking through mesophyll plasmodesmata.
The ability of PP2 N- and C-terminal deletion mutants to move cell to cell was also examined (Table 3). Only the N-terminal mutant (PP2 MT1) displayed a significant alteration in movement properties. Although PP2 MT1 was still able to increase plasmodesmal SEL and move cell to cell, its efficacy and extent of movement was significantly lower than the C-terminal deletion mutant (PP2 MT2) or wild-type PP2. Based on this preliminary analysis, it would appear that neither the N-terminal 77 nor the C-terminal 40 amino acid residues are essential for PP2 to interact with plasmodesmata. Presumably, the essential motif(s) must reside within the central region of this protein.
PP2 Threshold Concentration for Cell-to-Cell Transport.
Cell-to-cell movement of 10- and 20-kDa FITC-dextran was potentiated by low concentrations of size-fractionated phloem proteins (Table 1). To identify the threshold protein concentration required for successful interaction with mesophyll plasmodesmata, a series of microinjection experiments was conducted in which the level of 20-kDa FITC-dextran was held constant whereas the amount of PP2 was serially reduced. The results from these experiments are presented in Fig. 2. Interestingly, phloem sap-purified PP2 had a threshold concentration of between 0.025 and 0.04 μg⋅μl−1, whereas the value for E. coli-expressed PP2 was between 0.4 to 0.8 μg⋅μl−1; i.e., some 20 times higher. As similar threshold values were obtained with PP2 preparations that either contained the histidine-tag or for which this tag had been enzymatically cleaved (by using enterokinase), this difference cannot be attributed to the additional histidine residues.
Figure 2.
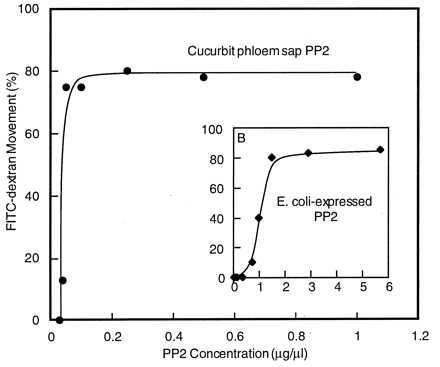
Microinjection experiments performed to determine the threshold level of PP2 required to potentiate cell-to-cell movement of coinjected 20-kDa FITC-dextran. PP2 used in these experiments was either purified from cucurbit phloem exudate or expressed in and extracted from E. coli (Inset). In each microinjection series, the level of FITC-dextran was held constant (1 mM) whereas the level of PP2 was serially reduced until fluorescence associated with the FITC-dextran did not move out from the injected cell.
Phloem Sap Proteins Traffic in Different Species.
To examine whether the phloem sap proteins from one species have the capacity to move through plasmodesmata in a different species, a series of microinjection experiments was performed on Ricinus and cucurbit phloem proteins. Microinjection of FITC-PP2 into mesophyll cells within mature leaves of N. tabacum, N. benthamiana, and A. majus resulted in cell-to-cell movement equivalent to that previously observed in cucurbit cotyledons (data not shown). Similarly, glutaredoxin (21) and cystatin (J. Szederkényi and C.S., unpublished results), which are abundant proteins in Ricinus phloem sap, were both able to increase plasmodesmal SEL (≥20 to ≤40 kDa) and move cell to cell when microinjected into mesophyll cells of cucurbit cotyledons (Table 4). However, in these experiments, the efficacy and extent of this movement was less than that obtained when cucurbit phloem proteins were employed in similar experiments. Interestingly, the data obtained in the Ricinus glutaredoxin experiments were very similar to the results from PP2 MT1 studies (see Tables 3 and 4).
Table 4.
Capacity of E. coli-expressed R. communis phloem sap proteins to move from cell to cell when microinjected into mesophyll cells within the cotyledons of C. maxima plants
| Injected material | Protein concentration, mg⋅ml−1 | Micro- injections* | Extent of movement |
|---|---|---|---|
| LYCH | — | 18 (18; 100%) | 20–30 cells |
| 20-kDa FD | — | 0 (18; 0%) | 0 cells |
| FITC-glutaredoxin | 5.0 | 10 (17; 59%) | 10–12 cells |
| Glutaredoxin + 20-kDa FD | 2.5 | 6 (12; 50%) | 10–12 cells |
| Glutaredoxin + 40-kDa FD | 2.5 | 1 (12; 8%) | 4 cells† |
| FITC-cystatin | 2.3 | 5 (11; 45%) | 10–12 cells |
| Cystatin + 20-kDa FD | 1.7 | 6 (12; 50%) | 10–12 cells |
| Cystatin + 40-kDa FD | 1.7 | 1 (10; 10%) | 3 cells† |
| Ubiquitin‡ + 20-kDa FD | 1.0 | 1 (10; 10%) | 3 cells† |
| FITC-ubiquitin§ | 3.0 | 0 (16; 0%) | 0 cells |
| FITC-ΔC-ubiquitin§ | 3.0 | 2 (12; 16%) | 3–4 cells† |
| FITC-ΔC-ubiquitin§ + PP2 | 2.0 + 2.0 | 0 (8; 0%) | 0 cells |
| FITC-ΔC-ubiquitin§ + KN1 | 2.0 + 2.0 | 1 (10; 10%) | 3 cells† |
FD, FITC-dextran.
Details as in Table 1.
When movement of fluorescence was detected, it was confined to adjacent cells.
Ubiquitin extracted from the phloem sap of R. communis.
E. coli-expressed A. thaliana ubiquitin (wild-type) and ΔC-ubiquitin (C-terminal deletion mutant).
Coinjection of ubiquitin with 20-kDa FITC-dextran revealed that this 8.5-kDa protein, present in the phloem sap of both Ricinus and cucurbits, was incapable of increasing plasmodesmal SEL (Table 4). This result may have been caused by (i) ubiquitin trafficking without an increase in SEL, or (ii) rapid conjugation of ubiquitin to proteins within the injected mesophyll cell which would have made it unavailable for movement through plasmodesmata. To explore these possibilities, Arabidopsis thaliana ubiquitin (wild-type) and a ΔC-ubiquitin mutant (C-terminal deletion mutant that is rendered incapable of protein conjugation) were expressed in E. coli and FITC-labeled proteins used in microinjection experiments. Again, these forms of ubiquitin were confined to the injected cell (Table 4). In view of this result, we next coinjected FITC-ΔC-ubiquitin with either PP2 or KN1 to ascertain whether cell-to-cell movement of the unreactive form of ubiquitin could be potentiated during the trafficking of these proteins. Interestingly, as shown by the data presented in Table 4, neither PP2 nor KN1 facilitated the movement of FITC-ΔC-ubiquitin. These experiments suggest that ubiquitin may require the presence of a companion cell-specific protein(s) to mediate its transport into the sieve tube system.
DISCUSSION
Plasmodesmal Trafficking of Phloem Proteins.
We here establish, via microinjection experiments carried out on cucurbit mesophyll tissue, that phloem-derived proteins have the capacity to function in a manner analogous to viral MPs. It is unfortunate that technical difficulties precluded experiments being performed on the companion cell-sieve tube member complex. Nevertheless, extensive microinjection studies carried out on neighboring mesophyll cells unambiguously established that cucurbit phloem sap proteins can interact with plasmodesmata to both induce an increase in SEL and potentiate their cell-to-cell transport through this tissue. Similar studies performed on tobacco and snapdragon revealed that these cucurbit phloem sap proteins exhibited the same properties in terms of their ability to traffic through mesophyll plasmodesmata. Furthermore, parallel experiments conducted on cucurbit cotyledons with specific Ricinus phloem sap proteins provided further confirmation that phloem proteins (from different species) have the capacity to traffic through plasmodesmata located within nonphloem tissues. Thus, the present results, in conjunction with a recent report that 10-kDa FITC-dextran can move between companion cells and the sieve tube members of Vicia faba (26) and the exciting finding that SUT1 mRNA can be detected in plasmodesmata between companion cells and sieve tube members (28), provide strong support for the hypothesis that, in the higher plant phloem, extensive macromolecular trafficking occurs between companion cells and sieve tube members.
Immunolocalization and in situ studies performed on wheat (13), cucurbit (20, 29), rice (30), and tobacco, potato, and tomato (28) have established that, when expressed in vivo, the sieve tube proteins are confined to the companion cell-sieve tube complex. Thus, our present findings reveal two important facets associated with plasmodesmal function: (i) the plasmodesmal supramolecular structure does not constitute, in itself, the only discrimination site for cell-to-cell transport of macromolecules; and (ii) additional cell-specific factors (cytoplasmic or plasmodesmal) are likely involved in the regulation of macromolecular trafficking across specific cellular boundaries (e.g., the companion cell-sieve tube member interface) (11). A general corollary would be that, in the absence of specific regulatory factors, plasmodesmata will permit the cell-to-cell movement of bona fide trafficking proteins when they are introduced into cells located outside of their normal domain (either by ectopic expression or microinjection).
In comparison to cucurbit phloem proteins, glutaredoxin and cystatin from the Ricinus phloem were somewhat less effective in terms of mediating their cell-to-cell transport through cucurbit mesophyll plasmodesmata. This reduction in efficacy (percent of movement as well as extent of trafficking) could well reflect a minor divergence between homologous proteins within cucurbit and Ricinus plasmodesmata (but see later discussion on E. coli-expressed proteins). In future studies we will test the ability of specific phloem proteins to traffic through plasmodesmata in more divergent species. Potentially, such experiments could provide an effective means of mapping the evolution of plasmodesmal macromolecular trafficking in the plant kingdom.
Protein Unfolding Appears Essential for Plasmodesmal Transport.
Our microinjection studies established that many cucurbit and Ricinus phloem sap proteins have the capacity to increase plasmodesmal SEL to a value greater than 20 but less than 40 kDa (Tables 1, 2, 3, 4). This finding is significant, for it demonstrates that protein trafficking through mesophyll plasmodesmata is accompanied by an increase in SEL, but that the absolute value does not depend on the actual size of the molecule being trafficked. The simplest interpretation of these results is that bona fide trafficking proteins must be partially unfolded prior to, or during, their transport through the plasmodesmal microchannel. Such an unfolding event, during transport, and refolding on exit from the plasmodesmal microchannel, would likely involve chaperones. In this regard, it is interesting to note that chaperones have been identified in Ricinus phloem sap (17). Although it has yet to be determined whether these phloem sap chaperones function at the level of plasmodesmal trafficking, it is possible that special families of plasmodesmal chaperones may have evolved to mediate both protein unfolding/folding and sorting/delivery to plasmodesmata located at specific cellular boundaries.
Phloem Sap Proteins Are Active at Low Concentrations.
With the exception of PP1 and PP2, the proteins in the cucurbit phloem sap are present at quite low levels. In the size-fractionation experiments presented in Table 1, the protein levels employed were adjusted to reflect the levels of these less abundant proteins within the original phloem sap. As the level of protein present in each of these fractions was sufficient to allow cell-to-cell movement of the coinjected FITC-dextran, it is clear that the cucurbit phloem proteins display a very high affinity for the endogenous components involved in trafficking of macromolecules through mesophyll plasmodesmata.
Based on our PP2 threshold studies, it is possible to gain an estimate of the concentration required, within the injected mesophyll cell, to allow PP2 movement into the neighboring cells. As shown in Fig. 2, PP2-mediated cell-to-cell movement of FITC-dextran occurred until the PP2 concentration in the microinjection pipette was reduced to ≈0.03 μg⋅μl−1; at 0.02 μg⋅μl−1 no movement was detected. Thus, PP2 (24 kDa) must be present, in the pipette tip, at a concentration of ≥1 × 10−12 mol/μl (1 μM) to allow a threshold quantity of protein to be microinjected into a target cell. For the present discussion, we will simply assume that, based on our observations of the relative levels of fluorescence present within the pipette tip and in the injected cell, a 10-fold dilution of PP2 occurs during the injection process. By using this estimate, the PP2 threshold for plasmodesmal transport would be in the range of 100 nM.
A second estimate for this threshold value can be gained based on analysis of experiments in which PP2 was injected at 0.05 μg⋅μl−1. Under these conditions, the 20-kDa FITC-dextran moved into at least 10 cells. Thus, based on our earlier analysis, the initial level of PP2 in the target cell would be 0.2 μM. If cell-to-cell movement stopped once the PP2 was further diluted within the 10 cells to the threshold level, this would yield a value of 20 nM. Certainly a threshold value for PP2 in the range of 20–100 nM would be consistent with the range of cell-to-cell movement reported in Table 3.
It is important to note that our analysis of PP2 movement has assumed that all of the PP2, present within the injection solution, is competent to traffic through plasmodesmata. The data presented in Fig. 2 for PP2 expressed in and purified from E. coli indicate that, when prepared by using this method, PP2 exhibits a 20 times lower affinity for plasmodesmal transport compared with PP2 extracted from the phloem sap. Although there may be a number of causes for this important difference, it may well reflect the presence of partially (improperly?) folded PP2 in the E. coli preparation. In view of this finding, the possibility cannot be discounted that a small fraction of partially folded PP2 may similarly exist in the PP2 sample prepared from the phloem sap. If this were the case, our estimates concerning the PP2 threshold concentration would represent an upper limit.
Threshold values for the less abundant phloem proteins are likely to be even lower than the value for PP2. This result can be seen by analysis of size fraction III which contained a number (≥10) of low-abundance proteins having a molecular weight from 25 to 50 kDa. As cell-to-cell movement was recorded by using a total protein concentration of 0.05 μg⋅μl−1, individual proteins would be present in the injection pipette at concentrations from 100 to 200 nM. Thus, the threshold values for individual fraction III proteins may be considerably below 10–20 nM.
As the threshold values for plasmodesmal transport of the phloem sap proteins fall within the same concentration range as many plant hormones, the possibility exists that some of these proteins may act as long-distance signaling molecules (18, 19). Plasmodesmata connect the functional sieve tube members to the surrounding cells/tissues along the entire long-distance translocation pathway. Thus, the site(s) at which a specific protein exits the translocation stream would be determined by the status of the plasmodesmata along the path; i.e., their capacity to allow selective trafficking of macromolecules. Although the molecular/cellular events underlying such regulation have yet to be elucidated, it is clear that such knowledge would provide important insights into the way in which higher plants orchestrate events at the whole-plant level.
Acknowledgments
Judit Szederkényi, Universität Bayreuth, Germany, provided the cystatin construct. This work was supported by Department of Energy Biosciences Grant DE-FG03-94ER20134 and Japan Tobacco Inc. Grant 94K077 (to W.J.L.). C.S. was supported by a fellowship from the German Research Community (SCHO 386/5-1).
ABBREVIATIONS
- MP
movement protein
- CMV-MP
cucumber mosaic virus movement protein
- FITC
fluorescein isothiocyanate
- LYCH
lucifer yellow CH
- PP1
phloem protein I
- PP2
phloem protein II
- SEL
size exclusion limit
References
- 1.Parthasarathy M V. In: Encyclopedia of Plant Physiology, New Series: Transport in Plants I. Phloem Transport. Zimmermann M H, Milburn J A, editors. Vol. 1. New York: Springer; 1975. pp. 3–38. [Google Scholar]
- 2.Raven J A. Plant Cell Environ. 1991;14:139–146. [Google Scholar]
- 3.Lucas W J, Ding B, van der Schoot C. New Phytol. 1993;125:435–476. doi: 10.1111/j.1469-8137.1993.tb03897.x. [DOI] [PubMed] [Google Scholar]
- 4.Lucas W J, Wolf S. Trends Cell Biol. 1993;3:308–315. doi: 10.1016/0962-8924(93)90013-q. [DOI] [PubMed] [Google Scholar]
- 5.Fujiwara T, Giesman-Cookmeyer D, Ding B, Lommel S A, Lucas W J. Plant Cell. 1993;5:1783–1794. doi: 10.1105/tpc.5.12.1783. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Noueiry A O, Lucas W J, Gilbertson R L. Cell. 1994;76:925–932. doi: 10.1016/0092-8674(94)90366-2. [DOI] [PubMed] [Google Scholar]
- 7.Ding B, Li Q, Nguyen L, Palukaitis P, Lucas W J. Virology. 1995;207:345–353. doi: 10.1006/viro.1995.1093. [DOI] [PubMed] [Google Scholar]
- 8.Nguyen L, Lucas W J, Ding B, Zaitlin M. Proc Natl Acad Sci USA. 1996;93:12643–12647. doi: 10.1073/pnas.93.22.12643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Lucas W J, Gilbertson R L. Annu Rev Phytopathol. 1994;32:387–411. doi: 10.1146/annurev.phyto.43.040204.135939. [DOI] [PubMed] [Google Scholar]
- 10.Lucas W J, Bouché-Pillon S, Jackson D P, Nguyen L, Baker L, Ding B, Hake S. Science. 1995;270:1980–1983. doi: 10.1126/science.270.5244.1980. [DOI] [PubMed] [Google Scholar]
- 11.Mezitt L A, Lucas W J. Plant Mol Biol. 1996;32:251–273. doi: 10.1007/BF00039385. [DOI] [PubMed] [Google Scholar]
- 12.Gilbertson R L, Lucas W J. Trends Plant Sci. 1996;1:260–267. [Google Scholar]
- 13.Fisher D B, Wu Y, Ku M S B. Plant Physiol. 1992;100:1433–1441. doi: 10.1104/pp.100.3.1433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Nakamura S, Hayashi H, Mori S, Chino M. Plant Cell Physiol. 1993;34:927–933. [Google Scholar]
- 15.Ishiwatari Y, Honda C, Kawashima I, Nakamura S, Hirano H, Mori S, Fujiwara T, Hayashi H, Chino M. Planta. 1995;195:456–463. doi: 10.1007/BF00202605. [DOI] [PubMed] [Google Scholar]
- 16.Sakuth T, Schobert C, Pecsvaradi A, Eichholz A, Komor E, Orlich G. Planta. 1993;191:207–213. [Google Scholar]
- 17.Schobert C, Groβmann P, Gottachalk M, Komor E, Pecsvaradi A, zur Nieden U. Planta. 1995;196:205–210. [Google Scholar]
- 18.Balachandran S, Hull R J, Vaadia Y, Wolf S, Lucas W J. Plant Cell Environ. 1995;18:1301–1310. [Google Scholar]
- 19.Lucas W J, Balachandran S, Park J, Wolf S. J Exp Bot. 1996;47:1119–1128. doi: 10.1093/jxb/47.Special_Issue.1119. [DOI] [PubMed] [Google Scholar]
- 20.Bostwick D W, Dannenhoffer J M, Skaggs M I, Lister R M, Larkins B A, Thompson G A. Plant Cell. 1992;4:1539–1548. doi: 10.1105/tpc.4.12.1539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Szederkényi J, Komor E, Schobert C. Planta. 1997;202:349–356. doi: 10.1007/s004250050137. [DOI] [PubMed] [Google Scholar]
- 22.Beers E P, Callis J. J Biol Chem. 1993;268:21645–21649. [PubMed] [Google Scholar]
- 23.Ding B, Haudenshield J, Hull R J, Wolf S, Beachy R N, Lucas W J. Plant Cell. 1992;4:915–928. doi: 10.1105/tpc.4.8.915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.van Bel A J E, Kempers R. Planta. 1991;183:69–76. doi: 10.1007/BF00197569. [DOI] [PubMed] [Google Scholar]
- 25.Kempers R, Prior D A M, van Bel A J E, Oparka K J. Plant J. 1993;4:567–575. [Google Scholar]
- 26.Kempers R, van Bel A J E. Planta. 1997;201:195–201. [Google Scholar]
- 27.Jackson D, Veit B, Hake S. Development (Cambridge, UK) 1994;120:405–413. [Google Scholar]
- 28.Kühn C, Franceschi V R, Schultz A, Lemoine R, Frommer W B. Science. 1997;275:1298–1300. doi: 10.1126/science.275.5304.1298. [DOI] [PubMed] [Google Scholar]
- 29.Dannenhoffer J M, Schultz A, Skaggs M I, Bostwick D E, Thompson G A. Planta. 1997;201:405–414. [Google Scholar]
- 30.Ishiwatari, Y., Fujiwara, T., McFarland, K. C., Nemoto, K., Hayashi, H., Chino, M. & Lucas, W. J. (1997) Planta, in press. [DOI] [PubMed]