

Abstract
Xanthophyll pigments have critical structural and functional roles in the photosynthetic light-harvesting complexes of algae and vascular plants. Genetic dissection of xanthophyll metabolism in the green alga Chlamydomonas reinhardtii revealed functions for specific xanthophylls in the nonradiative dissipation of excess absorbed light energy, measured as nonphotochemical quenching of chlorophyll fluorescence. Mutants with a defect in either the α- or β-branch of carotenoid biosynthesis exhibited less nonphotochemical quenching but were still able to tolerate high light. In contrast, a double mutant that was defective in the synthesis of lutein, loroxanthin (α-carotene branch), zeaxanthin, and antheraxanthin (β-carotene branch) had almost no nonphotochemical quenching and was extremely sensitive to high light. These results strongly suggest that in addition to the xanthophyll cycle pigments (zeaxanthin and antheraxanthin), α-carotene-derived xanthophylls such as lutein, which are structural components of the subunits of the light-harvesting complexes, contribute to the dissipation of excess absorbed light energy and the protection of plants from photo-oxidative damage.
The xanthophylls comprise a diverse group of oxygenated carotenoids with varied structures and multiple functions (1). In almost all photosynthetic eukaryotes, the majority of xanthophylls are bound with chlorophyll (Chl) molecules to proteins of integral membrane, light-harvesting complexes (LHCs) (2–5). The LHCs absorb and transfer excitation energy to the photosynthetic reaction centers to drive electron transport; these reactions convert light energy into chemical energy that is used to fix atmospheric CO2 into sugars. Xanthophylls can function as accessory light-harvesting pigments, as structural entities within the LHC, and as molecules required for the protection of photosynthetic organisms from the potentially toxic effects of light.
The importance of carotenoids in photoprotection is evident from the phenotypes of organisms that cannot synthesize carotenoids, either as a consequence of mutations or treatment with herbicides (e.g., norflurazon) that block carotenoid biosynthesis (6–9). There are several mechanisms by which carotenoids function to protect plants against photodamage. Specific xanthophylls are involved in the de-excitation of singlet Chl (1Chl) that accumulates in the LHC under conditions of excessive illumination (10–14). This de-excitation, measured as nonphotochemical quenching of Chl fluorescence (NPQ), depends on a large transthylakoid proton gradient that becomes established in excessive light. The development of NPQ correlates with the synthesis of zeaxanthin (Z) and antheraxanthin (A) from violaxanthin (V) via the xanthophyll cycle, which is depicted in Fig. 1. Estimates of excited state energy levels suggest that the lowest singlet state (21Ag or S1) of Z and A can accept excitation energy directly from 1Chl (15–17); the excited xanthophylls return to ground state by nonradiative heat dissipation. 1Chl can also enter the triplet state (3Chl) by intersystem crossing, and 3Chl can facilitate the formation of the highly toxic singlet oxygen molecule (1O2). The triplet states of xanthophylls can de-excite both 3Chl and 1O2 (18). The arrangement of the xanthophyll and Chl molecules in the LHC (19) allows for singlet and triplet energy transfer between these pigments either by a coulomb or a Dexter electron exchange mechanism (20). Finally, xanthophylls may be involved in inhibiting lipid peroxidation (21).
Figure 1.
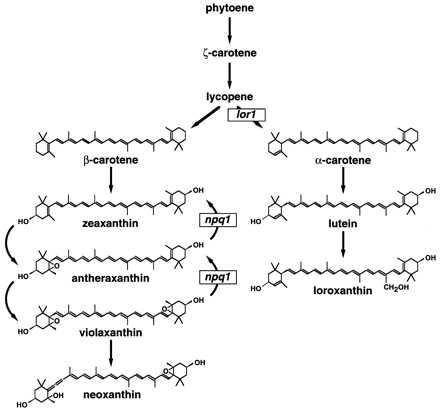
Carotenoid biosynthetic pathway in C. reinhardtii. The defects in xanthophyll metabolism in the npq1 and lor1 mutants are indicated.
To elucidate the roles of specific xanthophylls in photoprotection, we used a video imaging system to isolate mutants of the unicellular green alga Chlamydomonas reinhardtii that were impaired in NPQ and xanthophyll metabolism (22). Analysis of the npq1 mutant, which is unable to convert V to A and Z, demonstrated that operation of the xanthophyll cycle is not required for all pH-dependent NPQ in C. reinhardtii (22). In this report, we provide genetic evidence of a role in NPQ for xanthophylls derived from α-carotene, in addition to the xanthophyll cycle pigments, which are derived from β-carotene (Fig. 1). Characterization of single and double mutants has enabled us to define the specific xanthophylls that are required for protection of the photosynthetic apparatus from photo-oxidative damage in excessive light.
MATERIALS AND METHODS
Strains and Growth Conditions.
The npq1 mutant was isolated as described (22). The lor1 mutant (originally designated pg-101) (23, 24) was obtained from the Chlamydomonas Genetics Center (Duke University). Although it was first described as lacking only loroxanthin, lor1 also is unable to make α-carotene and lutein and is therefore likely defective in ɛ-cyclase activity. Genetic crosses were performed according to established methods (25). Mutants were backcrossed three times as mt− parents to the standard wild-type strain CC-125 (mt+) to eliminate other mutations that might be present in the different strain backgrounds, including the chloroplast sr-u-2–60 mutation that was in the background of the original npq1 isolate (22).
Cells were grown photoautotrophically in 100 ml minimal (high-salt) medium (25) with shaking in air in sterile beakers at 25°C with a 15 hr light/9 hr dark cycle (22). Viable cell number was determined by using a hemacytometer as described (26). Strain stocks were maintained in very low light (10 μmol photons m−2⋅s−1) at 27°C on agar medium containing acetate (Tris–acetate phosphate) (25).
Measurements of Fluorescence and Oxygen Evolution.
Cells (30 μg Chl) were deposited on a filter disc, and fluorescence parameters were measured in air (without CO2 enrichment) by using a modified pulse-amplitude modulation fluorometer system as described (22), except that the modulated measuring beam was provided by an LED with a peak output at 450 nm instead of 655 nm, and the duration of saturating light pulses (for determination of Fm or Fm′) was 250 ms instead of 1 s. Conventional fluorescence nomenclature was used (27). NPQ was calculated as (Fm − Fm′)/Fm′.
The light- and CO2-saturated rate of O2 evolution was measured by using a DW2/2 oxygen electrode chamber (Hansatech, Norfolk, U.K.) with 1.5 ml of cells (7.5 μg Chl/ml) and 4 mM NaHCO3 at 25°C. Actinic illumination (1090 μmol photons m−2⋅s−1) was provided by a 20 W halogen lamp via a fiber optic.
Pigment Determination.
Cells were frozen immediately in liquid nitrogen, and pigments were extracted with 90% (vol/vol) acetone in H2O. HPLC was performed by using a Spherisorb ODS-1 column as described (22). In addition, duplicate samples were analyzed on a Microsorb-MV (Rainin, Woburn, MA) column (28) to determine the proportion of neoxanthin and loroxanthin, which cochromatographed on the ODS-1 column. Pigments were eluted from the Microsorb-MV column at a flow rate of 1 ml/min with a 7-min linear gradient from 62% to 75% acetone, then an 8-min linear gradient from 75% to 80% acetone, followed by 7 min of 95% acetone, and finally a 3-min linear gradient from 95% to 100% acetone.
RESULTS AND DISCUSSION
A Role in NPQ for Xanthophylls Derived from α-Carotene.
The npq1 mutant is defective in the xanthophyll cycle and could not convert V to A and Z on exposure to excessive light (Figs. 1 and 2) (22). As a consequence, this mutant had a reduced ability to de-excite 1Chl (shown as reduced NPQ in Fig. 3). Nevertheless, excessive light did not impair the growth of npq1 (Fig. 4 and Table 1) under our growth conditions, probably because substantial NPQ still developed in this strain (22). These results with npq1 suggested that processes in addition to the xanthophyll cycle are involved in the de-excitation of 1Chl.
Figure 2.
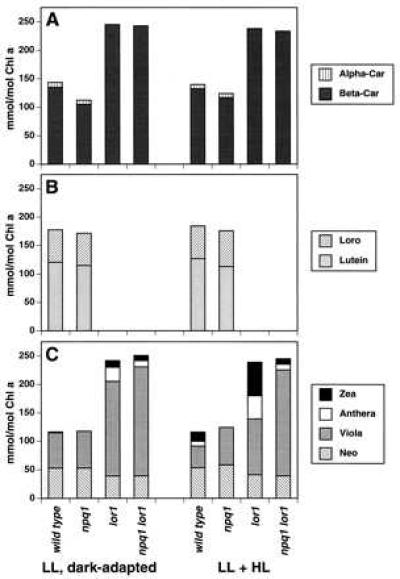
Carotenoid composition of C. reinhardtii strains. Cells were grown at an incident photon flux density (PFD) of 70 μmol photons m−2⋅s−1 (low light, LL) and then dark-adapted overnight. Carotenes (A), α-carotene-derived xanthophylls (B), and β-carotene-derived xanthophylls (C) were determined by HPLC analysis of cell extracts before (LL, dark-adapted) and after exposure to 1,160 μmol photons m−2⋅s−1 for 15 min [LL + high light (HL)] by using the same conditions as for measurements of fluorescence. Values are the means of two independent experiments. Loro, loroxanthin; Alpha-Car, α-carotene; Zea, zeaxanthin; Anthera, antheraxanthin; Viola, violaxanthin; Neo, neoxanthin; Beta-Car, β-carotene.
Figure 3.
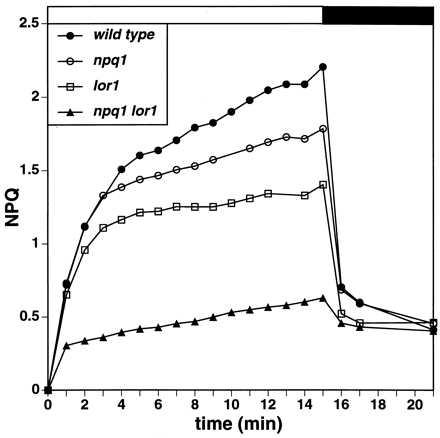
Induction of NPQ in C. reinhardtii strains. Cells were grown at an incident PFD of 70 μmol photons m−2⋅s−1 and dark-adapted overnight. Fluorescence was measured before, during, and after exposure to actinic light (1,160 μmol photons m−2⋅s−1) in air, and NPQ was calculated as (Fm − F′m)/F′m. The white bar above the graph indicates the period of illumination with high light (HL); the black bar indicates illumination with weak far red background light.
Figure 4.

Growth of C. reinhardtii strains. Cells were streaked on minimal agar medium, incubated overnight at 50 μmol photons m−2⋅s−1, and grown photoautotrophically for 6 days at the indicated PFD.
Table 1.
Growth and photosynthesis characteristics of C. reinhardtii strains
| Strain | Growth PFD, μmol photons m−2⋅s−1 | Chl doubling time, hr (n) | Fv/Fm (n) | O2 evolution, μmol O2 cell−1⋅hr−1(n) |
|---|---|---|---|---|
| Wild type | 70 | 20 ± 0 (4) | 0.763 ± 0.011 (6) | 1.21 ± 0.06 (2) |
| npq1 | 70 | 19 ± 2 (4) | 0.771 ± 0.009 (6) | 0.99 ± 0.07 (2) |
| lor1 | 70 | 23 ± 1 (4) | 0.793 ± 0.010 (6) | 0.96 ± 0.01 (2) |
| npq1 lor1 | 70 | 21 ± 1 (4) | 0.802 ± 0.004 (6) | 1.14 ± 0.16 (2) |
| Wild type | 350 | 28 ± 2 (4) | 0.769 ± 0.009 (3) | 0.90 ± 0.23 (2) |
| npq1 | 350 | 29 ± 3 (4) | 0.761 ± 0.004 (3) | 0.92 ± 0.06 (2) |
| lor1 | 350 | 34 ± 4 (4) | 0.718 ± 0.066 (3) | 0.36 ± 0.17 (2) |
| npq1 lor1 | 350 | 67 ± 16 (4) | 0.477 ± 0.050 (3) | 0.09 ± 0.00 (2) |
Each value is the mean ± SD (or for n = 2, mean ± difference from the mean). The maximum rate of O2 evolution was determined at light and CO2 saturation as described.
The major α-carotene-derived xanthophylls of the C. reinhardtii LHCs are lutein and loroxanthin. These xanthophylls are predicted to be energetically similar to A (15–17), suggesting that they might function in the direct de-excitation of 1Chl. This possibility is supported by the finding that the lor1 mutant of C. reinhardtii (23, 24), which cannot synthesize α-carotene, lutein, and loroxanthin (Figs. 1 and 2), had reduced NPQ (Fig. 3). Reduced NPQ in lor1 is unlikely to be caused by a lack of α-carotene, because wild-type cells (or npq1) accumulated very little α-carotene (Fig. 2), and a mutant that contained elevated levels of α-carotene did not exhibit increased NPQ (unpublished results). Hence, lutein, loroxanthin, or both must contribute to the de-excitation of 1Chl. The pH-dependent, rapidly reversible NPQ that remains in the lor1 strain can be attributed to the formation of Z and A via the xanthophyll cycle; these xanthophylls accumulated in lor1 to a greater extent than in wild-type cells (Fig. 2 and Table 2). Like the npq1 mutant, lor1 did not appear to be impaired significantly in photoautotrophic growth in high light (Fig. 4 and Table 1).
Table 2.
Pigment characteristics of C. reinhardtii strains
| Strain | Growth PFD, μmol photons m−2⋅s−1 | Chl a/Chl b (n) | (A+Z)/(V+A+Z) (n) | (V+A+Z)/Chl a, mmol/mol (n) | Lutein/Chl a, mmol/mol (n) | Loroxanthin/Chl a, mmol/mol (n) |
|---|---|---|---|---|---|---|
| Wild type | 70 | 2.64 ± 0.04 (6) | 0.066 ± 0.020 (2) | 72.1 ± 5.4 (2) | 115.2 ± 6.0 (2) | 58.9 ± 3.1 (2) |
| npq1 | 70 | 2.64 ± 0.03 (6) | 0.000 ± 0.000 (2) | 73.6 ± 5.1 (2) | 113.1 ± 6.3 (2) | 57.7 ± 2.8 (2) |
| lor1 | 70 | 4.02 ± 0.10 (6) | 0.213 ± 0.004 (2) | 221.5 ± 12.1 (2) | 0.0 ± 0.0 (2) | 0.0 ± 0.0 (2) |
| npq1 lor1 | 70 | 4.04 ± 0.07 (6) | 0.108 ± 0.001 (2) | 219.1 ± 15.1 (2) | 0.0 ± 0.0 (2) | 0.0 ± 0.0 (2) |
| Wild type | 350 | 2.87 ± 0.05 (3) | 0.598 ± 0.035 (2) | 114.3 ± 1.9 (2) | 209.8 ± 1.2 (2) | 20.2 ± 1.7 (2) |
| npq1 | 350 | 2.90 ± 0.02 (3) | 0.001 ± 0.001 (2) | 125.3 ± 12.1 (2) | 215.3 ± 20.4 (2) | 21.3 ± 0.1 (2) |
| lor1 | 350 | 4.17 ± 0.11 (3) | 0.696 ± 0.075 (2) | 320.8 ± 22.3 (2) | 0.0 ± 0.0 (2) | 0.0 ± 0.0 (2) |
| npq1 lor1 | 350 | 4.70 ± 0.29 (3) | 0.195 ± 0.001 (2) | 440.0 ± 46.8 (2) | 0.0 ± 0.0 (2) | 0.0 ± 0.0 (2) |
Pigment determination by HPLC was performed on samples of exponentially growing cells that were taken during the middle of the light period. Each value is the mean ± SD (or for n = 2, mean ± difference from the mean).
The C. reinhardtii lor1 mutant exhibited an elevated Chl a/Chl b ratio (Table 2) and decreased amount of neoxanthin (Fig. 2), which reflects a partial defect in assembly or stability of the peripheral LHCII (23). Similar pigment changes were reported for an analogous mutant of the green alga Scenedesmus obliquus (29). However, it is unlikely that the NPQ defect in lor1 is solely the consequence of altered assembly or stability of the peripheral LHCII; several observations suggest that lutein has a more direct role in NPQ. Recent data indicate that the inner rather than the peripheral LHC is the site of xanthophyll-related NPQ (30–35), and there is no evidence of a significant alteration of the inner LHC in the lor1 mutant. The Fv/Fm value of the lor1 mutant (and the lor1 npq1 double mutant) grown in low light was not decreased compared with that of wild-type cells (Table 1), suggesting that the lor1 lesion does not perturb the efficient transfer of absorbed light energy to the photosystem II reaction centers. As shown in Table 1, the Fv/Fm values of low light grown lor1 and npq1 lor1 were significantly greater than that of wild-type C. reinhardtii. Finally, the lut2 mutant of Arabidopsis thaliana, which is analogous to the lor1 mutant, has a normal Chl a/Chl b ratio (36), but is still impaired in NPQ (O.B., B. Pogson, D. DellaPenna, A.R.G., and K.K.N., unpublished data).
Specific Xanthophylls Required for Photoprotection.
Analysis of the npq1 lor1 double mutant demonstrated that blocking the synthesis of specific xanthophylls derived from both α- and β-carotene resulted in a cumulative reduction in NPQ. This double mutant was unable to synthesize α-carotene, lutein, and loroxanthin or to convert V to A and Z (Figs. 1 and 2). On exposure to high light, this strain displayed almost no reversible NPQ (Fig. 3), suggesting that it is severely impaired in the de-excitation of 1Chl. The reversible NPQ remaining in npq1 lor1 (Fig. 3) probably reflects the low levels of A and Z (Fig. 2 and Table 2) that accumulate as intermediates in the synthesis of V (Fig. 1).
The α-carotene-derived lutein and loroxanthin and the β-carotene-derived Z and A function redundantly in protecting C. reinhardtii from photo-oxidative damage. The npq1 and lor1 single mutants grew relatively normally in high light (Fig. 4 and Table 1), although the photosystem II efficiency (Fv/Fm) for lor1 appeared to be lower than that of wild-type cells (Table 1). In contrast, the npq1 lor1 double mutant, which attained a similar rate of growth in low light to wild-type cells, grew much more slowly than wild-type cells in high light (Table 1) and rapidly bleached (Fig. 4).
Increases in Z, A, and lutein during growth at elevated light intensities are consistent with a role of these xanthophylls in photoprotection. Wild-type C. reinhardtii grown at 350 μmol photons m−2⋅s−1 had greatly increased levels of Z and A, an increased xanthophyll cycle pool (V + A + Z), and elevated lutein relative to Chl a (Table 2). Increases in the xanthophyll cycle pool and lutein in high light occurred even in npq1, despite the inability of this strain to convert V to A and Z. Vascular plants and many algae grown in high light exhibit similar increases in xanthophyll cycle pigments (37–39). The greater accumulation of lutein in wild type and npq1 in high light was accompanied by a lower level of loroxanthin (Table 2). Light intensity-dependent changes in loroxanthin levels have been observed previously for the green alga Scenedesmus obliquus (40). The xanthophyll cycle pool in the lor1 strains was considerably elevated relative to that of wild-type or npq1 cells, even when lor1 was grown in low light (Table 2). This finding suggests that in the lor1 mutant the intermediates normally metabolized by the α-carotene biosynthetic pathway are diverted to the β-carotene branch; in agreement with this interpretation, the sum total of the carotenes and xanthophylls in all of the strains was remarkably similar for each specific growth condition. Together, these results suggest that Z, A, and lutein but not loroxanthin have critical functions in photoprotection and that the NPQ defect in the lor1 mutant is probably caused by the absence of lutein.
Multiple Levels of Xanthophyll-Dependent Photoprotection.
Z, A, and lutein have 10 or more conjugated double bonds and at least one cyclohexenyl ring with a single oxygen substituent (see Fig. 1). Xanthophylls with these characteristics could alter the fluidity of the photosynthetic membranes (41, 42) and/or the aggregation state of the LHCs within the membranes (43). The lack of an epoxide on at least one cyclohexenyl ring of Z, A, and lutein may facilitate a direct photochemical reaction with 1O2 (44), whereas having additional conjugated double bonds might make these xanthophylls more effective in preventing lipid peroxidation (21). Furthermore, the energies of the lowest singlet and triplet excited states decrease with increasing conjugation length. It is more energetically feasible for a direct transfer of excitation energy to occur from 1Chl to the singlet states of Z, A, and lutein than to V or neoxanthin (15, 16), which have fewer than 10 conjugated double bonds. Direct interaction between Chl and xanthophylls is consistent with the results of our genetic analysis, which revealed roles for Z, A, and lutein in the de-excitation of 1Chl in vivo. Alternatively, unique structural features of Z, A, and lutein may be required for protonation-induced conformation changes of specific inner LHC proteins to which these xanthophylls are bound; these changes could result in de-excitation of 1Chl that does not involve direct transfer of excitation energy from Chl to xanthophyll (14, 45).
The absence of xanthophyll-dependent de-excitation of 1Chl in the LHCs of the npq1 lor1 mutant would lead to elevated formation of 3Chl and 1O2. The latter species could cause irreversible photo-oxidative damage unless it were de-excited by interactions with carotenoids or scavenged by antioxidants such as α-tocopherol. β-Carotene, V, and neoxanthin (plus very low levels of A and Z) are essentially the only carotenoids in the npq1 lor1 mutant. The predominant carotenoids associated with the LHCs of this strain are likely to be V and neoxanthin, because these two xanthophylls, but not β-carotene, can substitute for lutein in LHC reconstitution assays (46). Both V and neoxanthin are probably able to function in photoprotection as they can de-excite 3Chl, thereby minimizing 1O2 production (18). However, V and neoxanthin have fewer than 10 conjugated double bonds, making these pigments less effective in the de-excitation of any 1O2 that does form in the npq1 lor1 strain (47). In contrast, Z, A, and lutein would be efficient scavengers of both 3Chl and 1O2; this is supported by the finding that C. reinhardtii mutants that are unaffected in their xanthophyll composition, but that cannot quench 1Chl (they exhibit essentially no NPQ) (22), can still survive high intensity illumination (unpublished results). Hence, the accumulation of V and neoxanthin in the npq1 lor1 strain is not sufficient for photoprotection in high light (Fig. 4); photoprotection in this strain is impaired at several levels because of a lack of Z, A, and lutein.
A Model for NPQ with Roles for Xanthophylls Derived from Both α- and β-Carotene.
The finding that lutein plays an important role in NPQ emphasizes how the generation and characterization of mutants can be used to dissect photoprotection in plants. The results presented here have enabled us to develop a model for NPQ that incorporates functions for both the α- and β-carotene-derived xanthophylls. Because LHCs can assemble in mutants lacking lutein [although there is some difference in the extent of assembly of LHCs in algal and plant mutants deficient in lutein (24, 29, 36, 48)], other xanthophylls must be able to fulfill the structural role of lutein in LHC assembly. In the case of the npq1 lor1 double mutant (and the lor1 single mutant grown in low light), V and neoxanthin (rather than lutein) must be the xanthophylls that form the “cross brace” in each LHC monomer (19). Because V can replace lutein in the LHCs, we propose that the xanthophylls derived from α- and β-carotene bind to the same sites in the LHCs of wild type. Based on the substoichiometric levels of β-carotene-derived xanthophylls per LHC monomer, it was suggested that these xanthophylls bind to sites on the LHC distinct from those to which lutein binds (19, 49). As an alternate interpretation, the substoichiometric amounts of some of the xanthophylls may simply reflect the heterogeneous xanthophyll population that can assemble with the LHC monomers (e.g., two luteins, one lutein and one V, or one lutein and one Z) (30). Because V is de-epoxidated efficiently in the lor1 mutant (Fig. 2), the V that assembles with the LHC must be accessible, either directly while still bound to the LHC polypeptides, or on exchange with a pool of free pigment, to the de-epoxidase that catalyzes the conversion of V to Z. This binding site would position Z (or lutein) in close proximity to the Chl a molecules that assemble with the LHC polypeptides and allow for the direct transfer of excitation energy from 1Chl to Z (or lutein).
Xanthophyll-dependent de-excitation of 1Chl also depends on acidification of the thylakoid lumen and/or localized thylakoid membrane domains (10–12, 14). Protonation of the LHC, probably specific polypeptides of the inner LHC (29–34), may promote a conformation change (14, 50) that favors the transfer of excitation energy from 1Chl to the xanthophylls that are bound to these particular proteins. Therefore, only a subset of the Z, A, and lutein pools are directly involved in the de-excitation mechanism. Because the lowest singlet energy state of lutein is likely to be higher than that of Z (and therefore closer to the lowest energy state of 1Chl), lutein may be a weaker quencher at pH values not saturating for NPQ. A greater proton gradient may be required to facilitate energy transfer from 1Chl to lutein either by causing a more substantial conformation change that brings the lutein closer to the Chl molecules or by changing the local electric field, which could provide sufficient spectral overlap between the lutein and proximal Chl molecules or alter the dipole strength of the lutein S1 to S0 transition (15).
This model accommodates roles in NPQ for both α- and β-carotene-derived xanthophylls and explains both in vivo and in vitro results, some of which have suggested that all NPQ depends on Z and A. From experiments with isolated thylakoid membranes, Gilmore and Yamamoto (51) derived the linear equation NPQ = χ1[H+][Z + A] + χ2[H+] + c. This equation includes the term χ2[H+] that relates NPQ to the proton concentration and is independent of [Z] and [A]. This term may reflect the contribution of the invariant α-carotene-derived xanthophylls such as lutein. Therefore, we suggest that the equation be modified to NPQ = χ1[H+][Z + A] + χ3[H+][L] + c, where [L] is the lutein concentration and χ3[L] = χ2. Assuming that χ3 is relatively small compared with χ1, which is consistent with lutein being a weaker quencher than Z, lutein-dependent NPQ would be most apparent at a high [H+], which occurs during the induction of photosynthesis on sudden illumination of dark-adapted cells with high light (the way in which the experiments in this study were performed). This interpretation is also consistent with the observation that only the rapid phase of NPQ is observed on illumination of the npq1 mutant (Fig. 2) (22). Others have noted xanthophyll cycle-independent NPQ under conditions likely to generate a high lumenal [H+], such as during the illumination of leaves in an atmosphere of 1% O2 and 0% CO2 (52). The contribution of the lutein concentration to NPQ would be less when the lumenal [H+] is only moderately high, conditions that occur during steady-state photosynthesis in high light or when the ATP synthase operates in reverse in isolated thylakoids in the dark. These latter conditions were used in many experiments that led to the conclusion that essentially all NPQ was dependent on Z and A (53–55).
Measurements of Chl fluorescence lifetime distributions in isolated thylakoids have distinguished two distinct components that contribute to a ΔpH-dependent decrease in the fluorescence yield (32, 54). A xanthophyll cycle pigment-independent shift in the fluorescence lifetime from 2.0 to 1.6 ns occurs on acidification of isolated thylakoid membranes. This shift, which we propose is a consequence of the de-excitation of 1Chl by lutein, involves the entire LHC population, probably because all of the LHC monomers contain at least one lutein (two luteins, lutein and V, or lutein and neoxanthin). When Z and A are present, a new low pH-dependent fluorescence lifetime component of 0.4 ns appears at the expense of the 1.6-ns component. The fractional intensity of this 0.4-ns component is correlated with the concentration of Z and A. This result is explained by our model, because Z and A would replace lutein, V, or neoxanthin in the LHC and lead to the more efficient de-excitation of 1Chl, thereby decreasing the fluorescence lifetime from 1.6 ns to 0.4 ns.
A similar process may occur when xanthophylls are added to isolated LHC particles and fluorescence quenching is measured on acidification (34, 56). The LHC particles used in these experiments contained mainly lutein and some V. They exhibited a certain level of NPQ that increased when Z was added to the preparations and decreased when V was added. According to our model the addition of Z would enhance quenching by exchanging for lutein and V within the LHC. Addition of V would inhibit quenching because some lutein bound in the LHC would be replaced by V, which is unable to de-excite 1Chl efficiently. Addition of lutein to the preparation would be predicted to have little or no effect.
In summary, the work presented here resolves some important questions concerning the xanthophyll dependence of NPQ and suggests a model for NPQ that accounts for several recent observations and that can now be tested further by using mutants. The analysis of mutants so far has demonstrated that almost all rapidly reversible NPQ depends on specific xanthophylls derived from both α- and β-carotene. Lutein is present constitutively in the LHCs and may function in NPQ mainly at high [H+]. These conditions activate the conversion of V to Z; the latter is a more effective acceptor of excitation energy from 1Chl, thereby allowing dynamic increases in the extent of photoprotection in excessive light. However, the results also emphasize that the xanthophylls can interact with 1Chl, 3Chl, and 1O2, proffering several lines of defense against photo-oxidative damage.
Acknowledgments
We thank Connie Shih for excellent technical assistance, Elizabeth Harris (Chlamydomonas Genetics Center, Duke University, Durham, NC) for strains, and Catharina Casper-Lindley and Dennis Wykoff for reviewing the manuscript. This work was supported by National Science Foundation Grant IBN 950-6254 (to A.R.G. and O.B.). K.K.N. is a Department of Energy–Energy Biosciences Fellow of the Life Sciences Research Foundation. K.K.N. was also supported by a fellowship from the Department of Energy/National Science Foundation/U.S. Department of Agriculture Training Program in Plant Biology at Stanford University. This is Carnegie Institution of Washington, Department of Plant Biology, publication no. 1346.
ABBREVIATIONS
- A
antheraxanthin
- Chl
chlorophyll
- LHC
light-harvesting complex
- NPQ
nonphotochemical quenching of Chl fluorescence
- PFD
photon flux density
- V
violaxanthin
- Z
zeaxanthin
References
- 1.Britton G. FASEB J. 1995;9:1551–1558. [PubMed] [Google Scholar]
- 2.Green B R, Durnford D G. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:685–714. doi: 10.1146/annurev.arplant.47.1.685. [DOI] [PubMed] [Google Scholar]
- 3.Grossman A R, Bhaya D, Apt K E, Kehoe D M. Annu Rev Genet. 1995;29:231–288. doi: 10.1146/annurev.ge.29.120195.001311. [DOI] [PubMed] [Google Scholar]
- 4.Jansson S. Biochim Biophys Acta. 1994;1184:1–19. doi: 10.1016/0005-2728(94)90148-1. [DOI] [PubMed] [Google Scholar]
- 5.Paulsen H. Photochem Photobiol. 1995;62:367–382. [Google Scholar]
- 6.Griffiths M, Sistrom W R, Cohen-Bazire G, Stanier R Y. Nature (London) 1955;176:1211–1215. doi: 10.1038/1761211a0. [DOI] [PubMed] [Google Scholar]
- 7.Sager R, Zalokar M. Nature (London) 1958;182:98–100. doi: 10.1038/182098a0. [DOI] [PubMed] [Google Scholar]
- 8.Anderson I C, Robertson D S. Plant Physiol. 1960;35:531–534. doi: 10.1104/pp.35.4.531. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Böger P, Sandmann G. Photosynthetica. 1994;28:481–493. [Google Scholar]
- 10.Demmig-Adams B. Biochim Biophys Acta. 1990;1020:1–24. [Google Scholar]
- 11.Demmig-Adams B, Adams W W., III Annu Rev Plant Physiol Plant Mol Biol. 1992;43:599–626. [Google Scholar]
- 12.Demmig-Adams B, Gilmore A M, Adams W W., III FASEB J. 1996;10:403–412. doi: 10.1096/fasebj.10.4.8647339. [DOI] [PubMed] [Google Scholar]
- 13.Gilmore A M. Physiol Plant. 1997;99:197–209. [Google Scholar]
- 14.Horton P, Ruban A V, Walters R G. Annu Rev Plant Physiol Plant Mol Biol. 1996;47:655–684. doi: 10.1146/annurev.arplant.47.1.655. [DOI] [PubMed] [Google Scholar]
- 15.Owens T G. In: Photoinhibition of Photosynthesis: From Molecular Mechanisms to the Field. Baker N R, Bowyer J R, editors. Oxford: BIOS; 1994. pp. 95–109. [Google Scholar]
- 16.Frank H A, Cua A, Chynwat V, Young A, Gosztola D, Wasielewski M R. Photosynth Res. 1994;41:389–395. doi: 10.1007/BF02183041. [DOI] [PubMed] [Google Scholar]
- 17.Chow W S. In: Advances in Molecular and Cell Biology. Bittar E E, Barber J, editors. Vol. 10. London: JAI; 1994. pp. 151–196. [Google Scholar]
- 18.Frank H A, Cogdell R J. In: Carotenoids in Photosynthesis. Young A, Britton G, editors. London: Chapman & Hall; 1993. pp. 252–326. [Google Scholar]
- 19.Kühlbrandt W, Wang D N, Fujiyoshi Y. Nature (London) 1994;367:614–621. doi: 10.1038/367614a0. [DOI] [PubMed] [Google Scholar]
- 20.Dexter D L. J Chem Phys. 1953;21:836–850. [Google Scholar]
- 21.Frank H A, Cogdell R J. Photochem Photobiol. 1996;63:257–264. doi: 10.1111/j.1751-1097.1996.tb03022.x. [DOI] [PubMed] [Google Scholar]
- 22.Niyogi K K, Björkman O, Grossman A R. Plant Cell. 1997;9:1369–1380. doi: 10.1105/tpc.9.8.1369. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Eichenberger W, Boschetti A, Michel H P. Physiol Plant. 1986;66:589–594. [Google Scholar]
- 24.Chunaev A S, Mirnaya O N, Maslov V G, Boschetti A. Photosynthetica. 1991;25:291–301. [Google Scholar]
- 25.Harris E H. The Chlamydomonas Sourcebook. San Diego: Academic; 1989. [Google Scholar]
- 26.Davies J P, Yildiz F H, Grossman A. EMBO J. 1996;15:2150–2159. [PMC free article] [PubMed] [Google Scholar]
- 27.van Kooten O, Snel J F H. Photosynth Res. 1990;25:147–150. doi: 10.1007/BF00033156. [DOI] [PubMed] [Google Scholar]
- 28.Plumley F G, Schmidt G W. Plant Cell. 1995;7:689–704. doi: 10.1105/tpc.7.6.689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Bishop N I, Urbig T, Senger H. FEBS Lett. 1995;367:158–162. doi: 10.1016/0014-5793(95)00510-g. [DOI] [PubMed] [Google Scholar]
- 30.Bassi R, Pineau B, Dainese P, Marquardt J. Eur J Biochem. 1993;212:297–303. doi: 10.1111/j.1432-1033.1993.tb17662.x. [DOI] [PubMed] [Google Scholar]
- 31.Härtel H, Lokstein H. Biochim Biophys Acta. 1995;1228:91–94. [Google Scholar]
- 32.Gilmore A M, Hazlett T L, Debrunner P G, Govindjee Photosynth Res. 1996;48:171–187. doi: 10.1007/BF00041007. [DOI] [PubMed] [Google Scholar]
- 33.Jahns P. Plant Physiol. 1995;108:149–156. doi: 10.1104/pp.108.1.149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ruban A V, Young A J, Horton P. Biochemistry. 1996;35:674–678. doi: 10.1021/bi9524878. [DOI] [PubMed] [Google Scholar]
- 35.Walters R G, Ruban A V, Horton P. Eur J Biochem. 1994;226:1063–1069. doi: 10.1111/j.1432-1033.1994.01063.x. [DOI] [PubMed] [Google Scholar]
- 36.Pogson B, McDonald K A, Truong M, Britton G, DellaPenna D. Plant Cell. 1996;8:1627–1639. doi: 10.1105/tpc.8.9.1627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Thayer S S, Björkman O. Photosynth Res. 1990;23:331–343. doi: 10.1007/BF00034864. [DOI] [PubMed] [Google Scholar]
- 38.Demmig-Adams B, Adams W W., III Plant Cell Environ. 1992;15:411–419. [Google Scholar]
- 39.Maxwell D P, Falk S, Huner N P A. Plant Physiol. 1995;107:687–694. doi: 10.1104/pp.107.3.687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Senger H, Wagner C, Hermsmeier D, Hohl N, Urbig T, Bishop N I. J Photochem Photobiol B. 1993;18:273–279. [Google Scholar]
- 41.Gruszecki W I, Strzalka K. Biochim Biophys Acta. 1991;1060:310–314. [Google Scholar]
- 42.Havaux M, Gruszecki W I. Photochem Photobiol. 1993;58:607–614. [Google Scholar]
- 43.Ruban A V, Phillip D, Young A J, Horton P. Biochemistry. 1997;36:7855–7859. doi: 10.1021/bi9630725. [DOI] [PubMed] [Google Scholar]
- 44.Schubert H, Kroon B M A, Matthijs H C P. J Biol Chem. 1994;269:7267–7272. [PubMed] [Google Scholar]
- 45.Young A J, Frank H A. J Photochem Photobiol B. 1996;36:3–15. doi: 10.1016/S1011-1344(96)07397-6. [DOI] [PubMed] [Google Scholar]
- 46.Plumley F G, Schmidt G W. Proc Natl Acad Sci USA. 1987;84:146–150. doi: 10.1073/pnas.84.1.146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Foote C S, Chang Y C, Denny R W. J Am Chem Soc. 1970;92:5216–5218. doi: 10.1021/ja00720a036. [DOI] [PubMed] [Google Scholar]
- 48.Bishop N I. J Photochem Photobiol B. 1996;36:279–283. [Google Scholar]
- 49.Lee A I, Thornber J P. Plant Physiol. 1995;107:565–574. doi: 10.1104/pp.107.2.565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Bilger W, Björkman O. Planta. 1994;193:238–246. [Google Scholar]
- 51.Gilmore A M, Yamamoto H Y. Photosynth Res. 1993;35:67–78. doi: 10.1007/BF02185412. [DOI] [PubMed] [Google Scholar]
- 52.Adams W W, III, Demmig-Adams B, Winter K. Plant Physiol. 1990;92:302–309. doi: 10.1104/pp.92.2.302. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Gilmore A M, Yamamoto H Y. Proc Natl Acad Sci USA. 1992;89:1899–1903. doi: 10.1073/pnas.89.5.1899. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Gilmore A M, Hazlett T L, Govindjee Proc Natl Acad Sci USA. 1995;92:2273–2277. doi: 10.1073/pnas.92.6.2273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Demmig-Adams B, Adams W W., III Trends Plant Sci. 1996;1:21–26. [Google Scholar]
- 56.Phillip D, Ruban A V, Horton P, Asato A, Young A J. Proc Natl Acad Sci USA. 1996;93:1492–1497. doi: 10.1073/pnas.93.4.1492. [DOI] [PMC free article] [PubMed] [Google Scholar]