

Abstract
The sentences we process in normal conversation tend to refer to information that we are familiar with rather than abstract, unrelated information. This allows for the use of knowledge stores to help facilitate comprehension processes. In many sentence comprehension studies, the stimuli are designed such that the use of world knowledge is limited. Here we investigated how the semantic relatedness of sentence constituents influences sentence processing. A three factor design was employed in which processing phase (sentence vs. probe), syntactic complexity (object-relative vs. conjoined active) and the semantic relatedness of the nouns within the sentence was examined. We found a differential effect in two sub-regions of the left inferior frontal gyrus (LIFG). BA 44 revealed an effect of syntactic complexity while inferior portions of the LIFG (BA 47) revealed an effect of relatedness as well as an interaction between complexity and relatedness during the probe phase. In addition, significant differences in activation were observed when comparing the sentence processing and probe phases with the sentence phase eliciting stronger semantic related activation while the probe phase elicited stronger working memory related activation.
Keywords: semantics, syntax
1. Introduction
The language processing system has been characterized by a set of separate, modular, processing systems which include systems that process orthographic, phonological, syntactic, and semantic information (Jackendoff, 2002, Levelt, 1999). There has been significant progress made in our understanding of these component processes of language comprehension. While there has been great advancement in our understanding of each individual processing system, how these systems work together, or collaborate, has received less attention. The goal of the current study was to explore the interaction between semantic and syntactic processing during sentence comprehension; specifically the study was designed to examine the influence semantics has on syntactic analysis. During normal communication sentence constituents are related to each other and as a result allow for the use of prior knowledge to aid in comprehension. For example, if discussing an event that occurred in a hospital it would be odd to encounter the word elephant but not so odd to encounter the words doctor or nurse. This is the bases of the current study with the general hypothesis tested being that when the sentence constituents are related syntactic analysis is facilitated.
There are a number of studies that have found that animacy information (a semantic feature) interacts with syntactic processing (Deutsch et al., 1999; Hoeks et al., 2004; Kim & Osterhout, 2005; Kolk et al., 2003; Kuperberg et al., 2007, 2006, 2005). Animacy information has been shown to play a role in syntactic analysis, particularly in assigning thematic roles (Kuperberg et al., 2005; Ferreira and Clifton, 1986; Chen et al., 2006). For example, in an fMRI study a differential effect of animacy on the processing of syntactically complex sentences was observed in a number of regions, including Broca’s area (Chen et al., 2006). Also, the P600, which has been observed during the processing of syntactic violations (Friederici et al., 1996; Hagoort et al., 1993; Kuperberg et al., 2005), was found to be influenced by animacy violations (Hoeks et al., 2004; Kim & Osterhout, 2005; Kolk et al., 2003; Kuperberg et al., 2005, 2007). Assuming that the P600 is a syntactic effect, finding this response for semantic violations suggests that semantic information can have an immediate effect on word processing within a sentence, and the integration of each word within the sentence.
While a P600 effect has been consistently observed for semantic violations, there seem to be little consensus as to what semantic factors result in the effect. One recent account, based on the extended Argument Dependency Model (eADM; Bornkessel-Schlesewsky & Schlesewsky, 2008), suggests that the P600 results from a mismatch of information obtained from the prominence and plausibility processing stages at the generalized mapping or well-formedness stage. According to eADM, prominence information (language specific information such as animacy and voice) is processed independently and parallel to plausibility (as well as world knowledge and discourse context) information. These two pieces of information are integrated at the generalized mapping stage when the final argument interpretation is determined and it is here where, if there is a conflict or mismatch, a P600 response will be generated (see Bornkessel-Schlesewsky & Schlesewsky, 2008 for details). This theory is important here for two reasons. First it argues that world knowledge/plausibility information is processed separately from other language-related information such as prominence information. This means that these two types of semantic information may have separable, independent influences on sentence processing. In fact, Bornkessel has demonstrated that prominence information is processed earlier than context/world knowledge information and that the neuro-anatomical correlates of prominence and plausibility processing are distinct (Bornkessel et al., 2003). For example, prominence information has been found to be associated with processing in the pars opercularis region of the left inferior frontal gyrus (BA 44) and the posterior superior temporal cortex (pSTC; Bornkessel & Schlesewsky, 2006; Grewe et al., 2006) while manipulations of plausibility have impacted the pars triangularis region of left IFG (BA 45; Friederici et al., 2003; Hagoort et al., 2004; Newman et al., 2003).
Just as animacy (prominence) has been shown to influence sentence comprehension, the semantic relatedness of sentence constituents (plausibility) may also affect sentence processing. ERP data suggest that the N400 is sensitive to the ease with which information is accessed in long-term memory (Kutas & Federmeier, 2000; Fischler et al., 1985). The sentence verification studies by Fischler and colleagues found that the N400 response was sensitive to category membership regardless of the truth value of the sentence. In other words, the N400 did not care if the sentences were true or not, only that there was a semantic relationship between the constituents of the sentence. These results suggest that the relationship between items in long-term memory seems to influence sentence processing which supports the current proposal. Therefore, the comprehension of sentences in which the sentence constituents are related to each other may be aided by long-term memory processes.
The current study was designed to explore how the semantic relatedness of the sentence constituents impacts comprehension. Many of the previous studies examining the interaction between syntax and semantics have used anomaly detection tasks instead of comprehension of grammatically intact sentences. Therefore, one of the goals of current study is to add to this literature by 1) examining comprehension processes and 2) manipulating semantic processing by varying the semantic relatedness of the nouns within the sentence. Manipulating the semantic relatedness is expected to impact plausibility processing and as has been shown previously plausibility can impact syntactic processing. Again, according to Bornkessel-Schlesewsky’s eADM, semantic relatedness, or world knowledge information is not equivalent to prominence information and has been shown to affect different cortical systems. The model (as does Hagoort and van Berkum, 2007) also argues that information such as semantic relatedness impacts lexical level processes and does not immediately impact sentence-level processing.
Bornkessel-Schlesewsky and colleagues (2009) have theorized that the prototypicality of the agent and patient may influence syntactic processing and it has been shown that the manipulation of prominence features modulates activation in the left IFG (Grodzinsky and Friederici, 2006; Grewe et al., 2006, 2007), and the left pSTC; both are regions that have been strongly linked to syntactic processing (Caplan et al., 2001; Caplan & Waters, 1999; Carpenter, et al., 1999; Cooke et al., 2006; Fiebach et al., 2001; Fiebach et al., 2005; Friederici et al, 2006; Friederici et al., 2003; Just et al., 1996; Keller et al., 2001; Newman et al., 2003). For example, a study conducted by Grewe and colleagues (2006) found that subject-initial sentences show increased activation in the left IFG compared to object-initial sentences when an inanimate nominative subject precedes an animate dative object which violates the more prominent animate-before-inanimate order. Grewe argues that the left IFG is involved in integrating both word order information as well as semantic information when constructing a representation of the sentence. The pSTC, on the other hand, has been linked to the assessment of prototypicality during comprehension and suggests that the region plays a role in generating the “relational construction of an argument hierarchy” (p. 40; Bornkessel-Schlesewsky, 2009).
Our hypotheses for the current study are as follows. The semantic relatedness of sentence constituents will facilitate sentence processing. It is unclear as to whether syntactic analysis will be facilitated in addition to semantic level processing. The eADM predicts that syntactic analysis per se would not be influenced by world knowledge. If there is facilitation of syntactic processing it would be expected to be observed in BA 44. This portion of Broca’s area has been implicated in syntactic processing (Fiebach et al. 2005; Friederici et al. 2003; Newman et al. 2003) and in the processing of prominence information (Bornkessel & Schlesewsky, 2006; Grewe et al., 2006). In addition, the activation of BA 45/47 is predicted to be modulated by relatedness. This is because the semantic relatedness manipulation used here - varying the semantic association of the sentence constituents - has some overlapping characteristics with the semantic plausibility manipulations discussed above (e.g., “The roofer that the chemist liked tossed a map out the window” Newman et al., 2009). Finally, previous studies have implicated the pSTG in both semantic and syntactic processing (Cooke et al., 2001; Friederici et al., 2003; Newman et al., 2009) and as a result the activation of this region may be expected to be modulated by both syntactic complexity and semantic relatedness. In order to test these region specific hypotheses a two-factor fMRI design in which semantic relatedness and syntactic complexity were within-participant variables was used. Semantic relatedness was manipulated by varying the relatedness of the first and second nouns within the sentences and complexity was manipulated by comparing object-relative and conjoined active sentences. Following the presentation of each sentence was a comprehension probe. A 6-sec delay was placed between the sentence and the probe to allow for the examination of brain activation associated with the sentence and the probe separately.
2. Results
2.1. Behavioral Results
The reaction time and error rate were analyzed (see Figure 1). Significant effects of complexity and relatedness were observed for both the reaction time [complexity: F(1,19) = 107.64, p < 0.0001; relatedness: F(1,19) = 14.26, p < 0.005] and error measures [complexity: F(1,19) = 37.25, p < 0.0001; relatedness: F(1,19) = 15.96, p < 0.001]. Neither measures revealed an interaction between the two variables (Fs<1).
Figure 1.
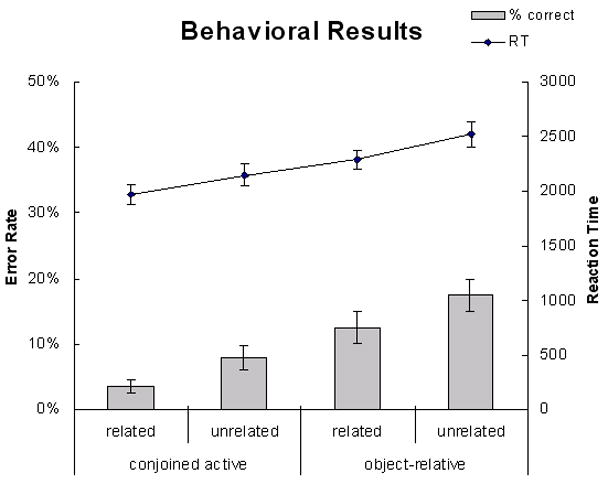
Behavioral data. Error bars depict the standard error.
2.2 fMRI Results
Analysis was performed on the sentence and probe phases separately. Contrasts were computed to examine the main effects of complexity, relatedness, and relatedness in each of the two processing phases as well as processing phase (sentence vs. probe).
2.2.1 Main Effect of Complexity
The three regions of interest, BA 44, BA 47, and the pSTG all revealed significant effects of complexity for both the sentence and the probe phases. No additional regions revealed a significant effect for the whole brain analysis using FDR (p< 0.05) (see Figure 2 and Table 1).
Figure 2.
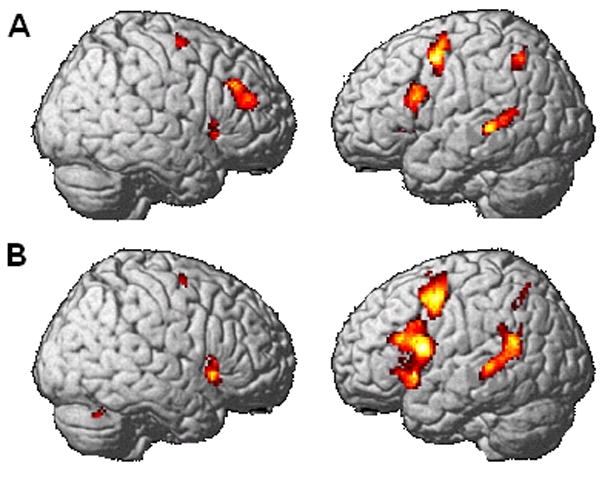
Main effect of syntactic complexity for the sentence phase (A) and the probe phase (B). The object-relative minus conjoined active contrast was performed using an uncorrected p<0.001 and an extent threshold of 20 voxels.
Table 1.
Main effect of complexity and relatedness in regions of interest
| Region | Cluster size | t-value | Coordinates x,y,z | |
|---|---|---|---|---|
|
Sentence Phase Complexity | ||||
| Left Inferior Frontal Gyrus | 44 | 1122 | 4.28 | −40,14,24 |
| Left Inferior Frontal/Insula | 47/13 | 4.34 | −30,22,0 | |
| Left Temporal | 22/21 | 356 | 4.55 | −58,−36,2 |
|
Sentence Phase Unrelated minus Related | ||||
| Left Inferior Frontal/Insula | 45/47 | 204 | 4.13 | −42,24,10 |
|
Probe Phase Complexity | ||||
| Left Inferior Frontal Gyrus | 44 | 2618 | 4.49 | −54,8,16 |
| Left Inferior Frontal/Insula | 47/13 | 5.43 | −32,20,4 | |
| Left Temporal | 22/21 | 1190 | 4.18 | −50,−38,0 |
|
Probe Phase Unrelated minus Related | ||||
| Left Inferior Frontal/Insula | 45/47 | 726 | 7.89 | −44,24,4 |
| Left Inferior Frontal Gyrus | 44 | 1185 | 7.56 | −56,14,28 |
2.2.2 Main Effect of Relatedness
BA 47 revealed a significant effect of relatedness for both the probe and sentence phases- greater activation for the unrelated compared to the related condition – for both the sentence and the probe processing phases. BA 44 also revealed an effect for the probe phase (see Figure 3 and Table 1). The whole brain analysis, using FDR to correct for multiple comparisons, revealed a number of other regions showing an effect of relatedness for the probe phase but no other regions were observed to have a relatedness effect for the sentence phase (see Table 2).
Figure 3.
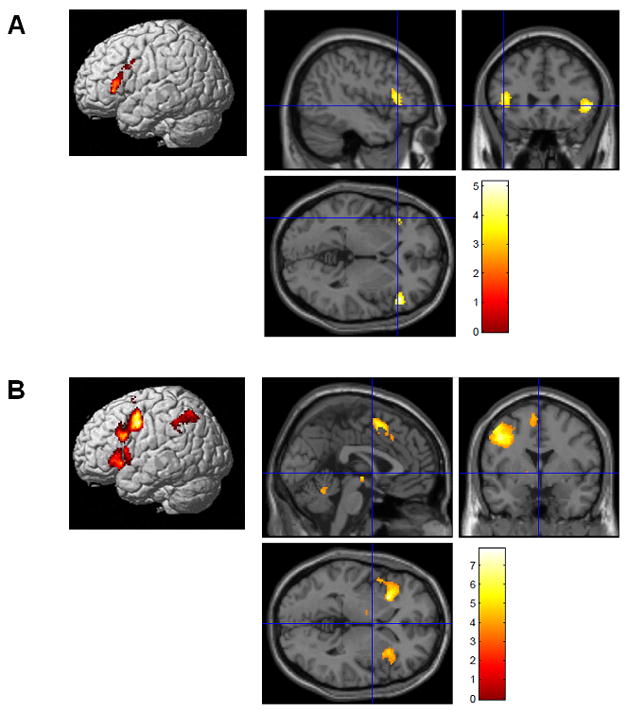
Main effect of relatedness for the sentence phase (A) and the probe phase (B). The unrelated minus related contrast was performed using an uncorrected p<0.001 and an extent threshold of 20 voxels.
Table 2.
Probe Phase Unrelated minus Related (FDR corrected, p<0.05)
| Region | Cluster size | t-value | Coordinates x,y,z | |
|---|---|---|---|---|
| Left Inferior Frontal/Insula | 13 | 726 | 7.89 | −28,18,−4 |
| 47 | 4.81 | −44,24,4 | ||
| Left Superior Frontal Gyrus | 6 | 782 | 5.09 | −8,6,56 |
| Left Precentral Gyrus | 6 | 1185 | 7.56 | −44,−2,40 |
| Left Superior Parietal | 7 | 812 | 6.64 | −26,−58,42 |
| Left Cerebellum | 69 | 5.06 | −2,−58,−20 | |
| Basal Ganglia | 86 | 4.89 | −6,−16,−12 | |
| Right Insula | 13 | 313 | 4.81 | 34,22,−2 |
| Right Prefrontal | 46 | 186 | 4.81 | 42,22,24 |
| Right Superior Parietal | 7 | 39 | 4.5 | 30,−58,42 |
| Right Precentral Gyrus | 4 | 41 | 4.41 | 40,−16,62 |
2.2.3 Complexity by Relatedness Interaction
An overadditive interaction between complexity and relatedness was observed in BA 47, which extends into the anterior insula, for the probe phase only (see Figure 4 and Table 3). The whole brain analysis, using FDR to correct for multiple comparisons, showed that the sentence phase elicited an interaction in a number of other regions within posterior cortex, including occipital regions such as the fusiform and lingual gyri (see Figure 4). The whole brain analysis did not reveal additional effects during the probe phase.
Figure 4.
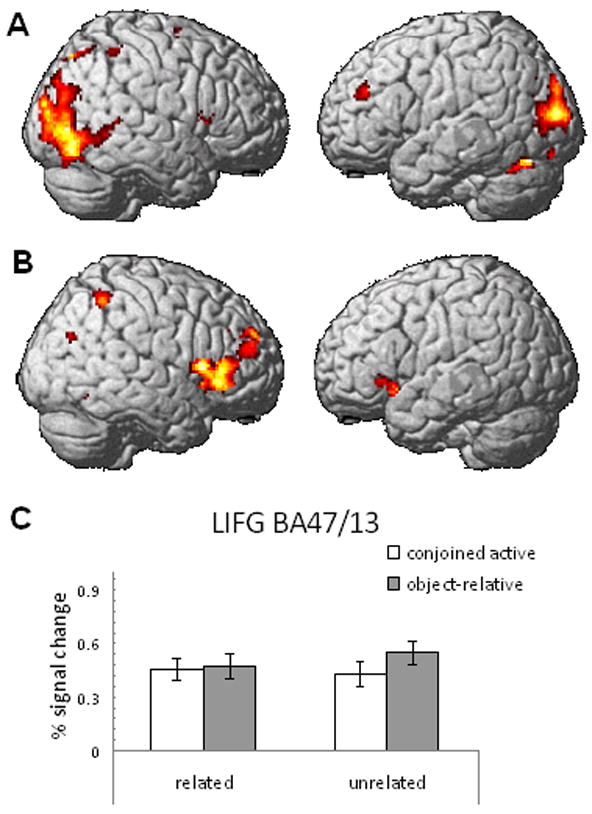
Interaction between complexity and relatedness during the sentence phase (A) and the probe phase (B). The contrast was performed using an uncorrected p<0.001 and an extent threshold of 20 voxels. (C) Depicts the nature of the interaction observed in BA 47/13 by showing the percent signal change in the region during the probe phase.
Table 3.
Interaction between complexity and relatedness in regions of interest
| Region | Cluster size | t-value | Coordinates x,y,z | |
|---|---|---|---|---|
|
Probe Phase | ||||
| Left Inferior Frontal | 47/13 | 154 | 4.76 | −32,18,−8 −36,30,−4 |
2.2.4 Effect of Processing Phase
In previous studies we have observed significant processing differences when comparing the sentence and the probe phases (Lee & Newman, 2010; Newman et al., 2009). Similar results were found here for the probe minus sentence contrast – greater activation in regions that have been linked to working memory and motor processes like the inferior parietal cortex, anterior cingulate and the cerebellum (see Figure 5 and Table 4). However, unlike in the previous study, the current study found that the sentence phase elicited greater activation in temporal, occipital and basal ganglia regions.
Figure 5.

The effect of processing phase: sentence minus probe (A) and probe minus sentence (B). The contrast was performed using an uncorrected p<0.001 and an extent threshold of 20 voxels.
Table 4.
Main effect of processing phase (FDR corrected, p < 0.05)
| Region | Cluster size | t-value | Coordinates x,y,z | |
|---|---|---|---|---|
|
Sentence minus Probe | ||||
| Right Cuneus | 17/18 | 13900 | 12.56 | 18,−94,2 |
| Left Inferior Temporal | 21 | 216 | 6.89 | −58,−10,−12 |
| Left Basal Ganglia (Caudate) | 1070 | 5.14 | −2,12,10 | |
| Left Superior Temporal | 22 | 74 | 4.61 | −48,−46,8 |
| Left Parietal | 3 | 607 | 4.56 | −52,−10,50 |
| Right Precentral | 6 | 402 | 4.03 | 46,−8,34 |
| Right Middle Temporal | 21 | 131 | 4.13 | 62,−8,−12 |
|
Probe minus Sentence | ||||
| Left Insula/Parietal | 13/40 | 8820 | 11.54 | −40,−6,2 |
| Left Superior Frontal | 10 | 8421 | 6.93 | −22,48,4 |
| Left Middle Frontal | 8/6 | 298 | 5.02 | −42,22,40 |
| Anterior Cingulate | 24 | 1113 | 5.76 | −10,−2,40 |
| Right Inferior Parietal | 40 | 7169 | 8.79 | 56,−58,40 |
| Right Middle Frontal | 8/9 | 935 | 8.14 | 42,12,44 |
| Left Cerebellum | 190 | 4.75 | −20,−50,−30 | |
| Right Cerebellum | 127 | 4.69 | 34,−68,−40 | |
2.3. Ancillary Behavioral Study
39 participants (mean age = 21.71, SD = 3.22), all of whom gave informed written consent, took part in a behavioral study. The same stimuli were used in a self-paced, word by word reading study. Stimuli were presented one word at a time in the center of the screen and participants pressed the space bar after each word was read in order to obtain word reading times. The probe was presented after each sentence in a whole sentence presentation format. When examining the verbs, both verb1 and verb2 revealed significant effects of syntactic complexity [F(1,38) = 13.44, p < 0.001; F(1,38) = 36.46, p < 0.001, respectively]. However, only verb2 revealed a significant effect of relatedness with the semantically related condition showing a faster reading time than the unrelated condition, particularly for the object-relative sentences [verb1: F<1; verb2: F(1,38) = 4.48, p < 0.05], see Figure 6. Neither revealed a significant interaction between complexity and relatedness.
Figure 6.
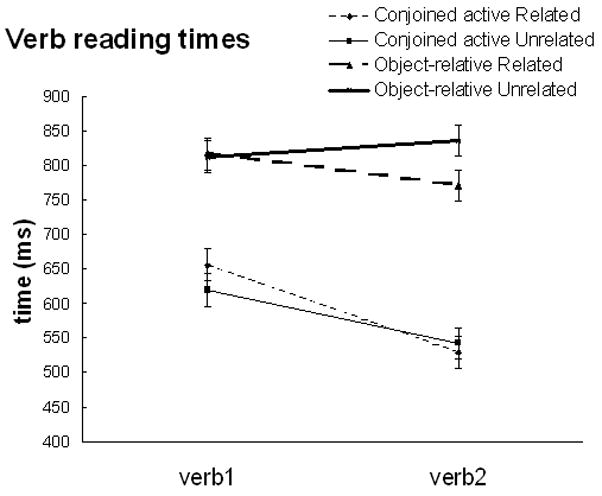
Ancillary Behavioral Study results. Error bars depict the standard error.
3. Discussion
The aim of the current study was to explore how the semantic relatedness of sentence constituents interacts with syntactic processes. Here we found syntactic complexity effects in BA 44, the posterior temporal cortex and in inferior parietal cortex. We also found that inferior portions of the LIFG revealed a significant effect of relatedness as well as an interaction between relatedness and complexity during the probe phase. These results seem to support the claim made by eADM that semantic information such as world knowledge is processed separately from prominence information and does not significantly affect on-line syntactic processing. In addition, we extended previous findings that revealed differential processing during the sentence and the probe phase (Lee & Newman, 2010; Newman et al., 2009). In the paragraphs below we discuss these findings in greater detail.
3.1. Interaction between semantics and syntax
Previous studies have provided evidence to support the hypothesis that semantic information is available earlier than some syntactic information (Freedman and Forster, 1985; Boland 1993; Murray and Rowan, 1998) suggesting that it could have a significant impact on syntactic analysis. In the current study we failed to find evidence that semantic relatedness directly impacts syntactic analysis in that the regions that have been most strongly implicated in syntactic processing, BA 44 and pSTC, both failed to show a significant interaction between complexity and relatedness. The IFG region where relatedness effects were observed was BA 47. There effects of relatedness and an interaction between relatedness and complexity were observed. The observation of these effects in BA 47 provides support for the eADM claims. First, the model claims that world knowledge information such as that manipulated here is processed separately from prominence information. Given that prominence manipulations have been found to modulate the activation of BA 44, not BA 47, this first claim is supported here. Second, the model postulates that the effect of world knowledge information on sentence processing is rather late. While fMRI does not provide the temporal resolution to examine early versus late processing, here we found an interaction between complexity and relatedness during the probe, not the sentence phase, suggesting that this effect may be on the well-formedness processing stage. In fact, Bornkessel-Schlesewsky & Schlesewsky (2008) argued that the semantic P600 observed in many studies (e.g., Kuperberg et al., 2003; Kim & Osterhout, 2005; Kolk et al., 2003) may be explained by a disruption of the well-formedness processing stage, a processing stage that occurs late in the model.
The main verb is a critical point in comprehension and it has been shown that for object-relative sentences (and subject-relative) the processing time is significantly longer than for simple syntactic structures such as conjoined actives (King & Just, 1991; Christiansen & Chater, 1999). One possible reason for the increased processing time is that thematic role assignments are made (and anticipated) when the main verb is encountered (Tenenhaus et al., 1989; Kamide et al., 2003). Here we found that in the ancillary behavioral study the processing time at the second verb in object-relative sentences is significantly reduced in semantically related sentences. This suggests that semantic information is indeed coming on-line early enough to possibly constrain the thematic role assignment process, at least for the second verb in object relative sentences. In fact, a similar region, BA 45/47, has been implicated in the process of thematic role assignments (Newman et al., 2003). For example, in a previous study in which we examined anomalous sentences where the anomaly (an extra verb) disrupted thematic role assignment, increased activation was observed in BA 45/47 (Newman et al., 2003). An overlapping region, BA 47 (and the anterior insula), was found to show a significant effect of relatedness with the unrelated condition eliciting greater activation than the related condition. It may be that the semantic relatedness of the nouns is priming the verb representation (also although we did not test this, the verbs were also related to the nouns). If this is the case, then the argument structure information that is stored with the verb is also primed, which may result in the facilitation of syntactic processing, in this case thematic role assignment which may be occurring in BA 47. Further work is necessary to detangle this relationship.
3.2. Processing phase differences
In the current study we examined the sentence and probe phases separately. As in our previous studies we observed processing differences across phase (Lee et al., 2008; Newman et al., 2009). In those studies we observed increased activation for the probe phase compared to the sentence phase in regions related to working memory processing but no regions revealed greater involvement during the sentence phase. In contrast, here we found that the temporal cortex along with the basal ganglia show greater activation during the sentence phase than the probe phase. The temporal cortex has been linked to memory retrieval processing for lexical-semantic knowledge (Friederici et al. 2003; Keller et al. 2001) and thematic role knowledge (Chatterjee et al. 1995; Kable et al. 2002; Wu et al. 2007). Therefore, finding greater involvement of this region during the sentence than the probe, when there is greater lexical access demands – due to the repetition of lexical items in the probe phase - may be expected. The role of the basal ganglia (BG) in language functioning, however, is not clear. There are a number of studies that show damage to the region results in language deficits (Copland, 2003; Cappa & Vallar, 1992; Wallesch & Papagno, 1988; Nadeau & Crosson, 1997 for review) as well as neuroimaging studies showing activation of the region during language tasks (Ni et al., 2000; Price et al., 1997; Kuperberg et al., 2000; Fiebach et al., 2002). Two language impairments have been observed in patients with BG damage, including Parkinson’s patients: lexical/semantic processing deficits such as difficulty with confrontation naming and word finding difficulties (Cappa & Vallar, 1992; Wallesch & Papagno, 1988); and a syntactic processing deficit (Grossman et al., 2003; Lieberman et al., 1992; Frisch et al., 2003). While it is difficult to determine based on the current study whether the region is more involved in semantics or syntax, the current study adds to this literature in that here, we too, observed the involvement of the basal ganglia as well as its modulation as a function of semantics.
4. Methods
4.1. Participants
A total of 20 students (15 females, mean age =22.9, SD =3.13) from the Indiana University community participated the study. All participants were required to have English as their native language. Written informed consent that was approved by the university IRB was obtained prior to participation.
4.2. Materials
The current fMRI experiment used a single trial event-related design in which each trial was treated as an event block (Kruggel and von Cramon 1999; Zarahn et al. 1997; Zarahn, 2000; Fig. 1). A trial could be divided into two phases; a 5 sec sentence reading phase and a 5 sec responding to a comprehension probe phase. A 6 sec delay was placed between the sentence and probe to allow for the examination of sentence and probe separately. Participants were instructed to read each sentence thoroughly and respond as quickly and accurately as possible to probes. Participants were told to place a greater weight on accuracy than speed of responding.
The semantically unrelated sentence materials were taken from Keller et al. (2001) which were derived originally from Just et al. (1996). Syntactic complexity (conjoined active and object-relative sentences) was a within-participant variable. The object-relative sentences are syntactically more complex (Caplan et al., 1999; Just et al. 1996; Keller et al. 2001). Stimuli were equated across conditions for frequency, word length, sentence length and animacy.
Before the semantically related sentence stimuli were constructed a survey was conducted by 305 individuals in which participants rated the semantic relatedness of noun pairs using a scale of 1–4. The noun pairs in the survey were prospective nouns that were used to generate sentence stimuli as well as the nouns that were used in the Keller et al. sentences. This information was then used to generate the semantically related sentences presented here. Example stimuli include:
| Conjoined Active | |
| Related Noun | The inspector questioned the suspect and made the report for the trial. |
| The inspector questioned the suspect. | |
| Unrelated Noun | The photographer hugged the bride and caused the riot in the city. |
| The photographer caused the riot. | |
| Object Relative | |
| Related Noun | The businessman that the investor financed bought the company with the stock. |
| The investor financed the businessman. | |
| Unrelated Noun | The roofer that the chemist liked tossed a map out the window. |
| The roofer tossed a map. | |
As demonstrated in the example stimuli, the unrelated sentences, by virtue of the sentence constituents not being related, are somewhat anomalous compared to the related sentences. However, these sentences have been and are currently being used in research. As such, it is of interest to determine how they differ from the more natural sentences that we encounter in our everyday lives.
After the sentences were generated a survey was again conducted that contained the first and second noun of each sentence. The relatedness scores were significantly different (p<0.001) for the related and unrelated noun conditions but not for the conjoined active and object-relative conditions (p>0.07) and there was no interaction found between the two factors (p>0.3).
There were 20 trials per condition which were evenly and randomly presented across 4, eight minute functional runs. A trial consisted of a sentence and a comprehension probe. The probes were true/false statements based on the sentence. The probes inquired about who did what to whom and references either the first or second mentioned verb. False statements were created by stating that the wrong noun performed the given action. In addition, each run contained 3, 28 sec fixation periods located at the beginning, middle and end of each run. The specifics of the presentation methods have been published previously in Newman et al., 2009.
4.3. fMRI acquisition and analysis
Functional MRI was conducted on a 3T Siemens TRIO scanner with an 8-channel radio frequency head coil located in the Imaging Research Facility at Indiana University. Functional images were obtained in eighteen oblique axial slices with 5mm thickness and a 1mm gap (TR = 1000 msec, TE = 25 msec, flip angle = 60°, matrix size = 64×64, FOV = 240×240 mm2) by a gradient echo planar imaging (EPI) sequence. Analysis methods have been published previously in Newman et al., 2009.
Regions of interest definitions were based on previous studies of syntactic complexity (Lee & Newman, 2010; Newman et al., 2009) and included BA 44, BA 45/47, BA 13, and posterior superior temporal cortex (pSTG). The correction for multiple comparisons for the a priori predicted activation in the LIFG and pSTG was carried out using an uncorrected p value of 0.005 and a cluster size threshold of 20; this corresponds to a per-voxel false-positive probability of less than 0.000001 (Forman et al., 1995). This method has been utilized previously (Knutson et al., 2006; Knutson et al., 2004; Wood et al., 2003; Konishi et al., 1998; Poldrack et al., 1999). For the whole-brain analyses, correction for multiple comparisons was carried out using the false discovery rate (FDR) approach (Benjamini & Yekutieli, 2001; Yekutieli & Benjamini, 1999).
Acknowledgments
This research was supported by a National Institute of Health grant (R03 HD051579–01). We would like to thank Ben Pruce, Jr., John Greco, Tara Muratore and Kristen Ratliff for all of their help with data collection.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Awh E, Smith EE, Koeppe RA, Schumacher EH, Katz S. Dissociation of storage and rehearsal in verbal working memory: evidence from PET. Psychol Sci. 1996;7:125–131. [Google Scholar]
- 2.Benjamini Y, Yekutieli D. The control of false discovery rate in multiple testing under dependency. Annals of Statistics. 2001;29:1165–1188. [Google Scholar]
- 3.Boland JE. The relationship between syntactic and semantic processes in sentence comprehension. Language and Cognitive Processes. 1997;12:423–484. [Google Scholar]
- 4.Bornkessel I, Schlesewsky M. The Extended Argument Dependency Model: A neurocognitive approach to sentence comprehension across languages. Psychological Review. 2006;113:787–821. doi: 10.1037/0033-295X.113.4.787. [DOI] [PubMed] [Google Scholar]
- 5.Bornkessel I, Schlesewsky M, Friederici AD. Eliciting thematic reanalysis effects: the role of syntax-independent information during parsing. Language and Cognitive Processes. 2003;18:268–298. [Google Scholar]
- 6.Bornkessel-Schlesewsky I, Schlesewsky M. An alternative perspective on “semantic P600“ effects in language comprehension. Brain Research Reviews. 2008;59:55–73. doi: 10.1016/j.brainresrev.2008.05.003. [DOI] [PubMed] [Google Scholar]
- 7.Bornkessel-Schlesewsky I, Schlesewsky M. The role of prominence information in the real-time comprehension of transitive constructions: a cross-linguistic approach. Language and Linguistics Compass. 2009;3/1:19–58. [Google Scholar]
- 8.Bornkessel-Schlesewsky I, Schlesewsky M. An alternative perspective on ‘semantic P600’ effects in language comprehension. Brain Research Reviews. 2008;59:55–73. doi: 10.1016/j.brainresrev.2008.05.003. [DOI] [PubMed] [Google Scholar]
- 9.Caplan D, Alpert N, Waters G. PET studies of sentence processing with auditory sentence presentation. Neuroimage. 1999;9:343–351. doi: 10.1006/nimg.1998.0412. [DOI] [PubMed] [Google Scholar]
- 10.Caplan D, Vijayan S, Kuperberg G, West C, Waters G, Greve D, Dale AM. Vascular response to syntactic processing: an event-related fMRI study of relative clauses. Hum Brain Mapp. 2001;15:26–38. doi: 10.1002/hbm.1059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Caplan D, Alpert N, Waters G. Effects of syntactic structure and propositional number on patterns of regional cerebral blood flow. J Cogn Neurosci. 1998;10:541–552. doi: 10.1162/089892998562843. [DOI] [PubMed] [Google Scholar]
- 12.Caplan D, Alpert N, Waters G. PET studies of sentence processing with auditory sentence presentation. Neuroimage. 1999;9:343–351. doi: 10.1006/nimg.1998.0412. [DOI] [PubMed] [Google Scholar]
- 13.Caplan D, Waters G. Verbal working memory and sentence comprehension. Behav Brain Sci. 1999;22:114–126. doi: 10.1017/s0140525x99001788. [DOI] [PubMed] [Google Scholar]
- 14.Cappa SF, Vallar G. Neuropsychological disorders after subcortical lesions: implications for neural models of language and spatial attention. In: Vallar G, Cappa SF, Wallesch C-W, editors. Neuropsychological disorders associated with subcortical lesions. Oxford, UK: Oxford University Press; 1992. pp. 7–41. [Google Scholar]
- 15.Carpenter PA, Just MA, Reichle ED. Working memory and executive function: evidence from neuroimaging. Curr Opin Neurobiol. 2000;10:195–199. doi: 10.1016/s0959-4388(00)00074-x. [DOI] [PubMed] [Google Scholar]
- 16.Chatterjee A, Maher LM, Heilman KM. Spatial characteristics of thematic role representation. Neuropsychologia. 1995;33:643–648. doi: 10.1016/0028-3932(94)00134-b. [DOI] [PubMed] [Google Scholar]
- 17.Chen L, West WC, Waters G, Caplan D. Determinants of BOLD signal correlates of processing object-extracted relative clauses. Cereb Cortex. 2006;42:591–604. doi: 10.1016/s0010-9452(08)70397-6. [DOI] [PubMed] [Google Scholar]
- 18.Christiansen MH, Chater N. Toward a connectionist model of recursion in human linguistic performance. Cognitive Science. 1999;23:157–205. [Google Scholar]
- 19.Clark CR, Egan GF, McFarlane AC, Morris P, Weber D, Sonkkilla C, Marcina J, Tochon-Danguy HJ. Updating working memory for words: a PET activation study. Hum Brain Mapp. 2000;9:42–54. doi: 10.1002/(SICI)1097-0193(2000)9:1<42::AID-HBM5>3.0.CO;2-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Cooke A, Zurif EB, DeVita C, Alsop D, Koenig P, Detre J, Gee J, Pinango M, Balogh J, Grossman M. Neural basis for sentence comprehension: grammatical and short-term memory components. Hum Brain Mapp. 2001;15:80–94. doi: 10.1002/hbm.10006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Copland D. The basal ganglia and semantic engagement: potential insights from semantic priming in individuals with subcortical vascular lesions, Parkinson’s disease, and cortical lesions. Journal of the International Neuropsychological Society. 2003;9:1041–1052. doi: 10.1017/S1355617703970081. [DOI] [PubMed] [Google Scholar]
- 22.Ferreira F, Clifton C., Jr The independence of syntactic processing. J Mem Lang. 1986;25:348–368. [Google Scholar]
- 23.Fiebach CJ, Schlesewsky M, Lohmann G, von Cramon DY, Friederici AD. Revisiting the role of Broca’s area in sentence processing: syntactic integration versus syntactic working memory. Hum Brain Mapp. 2005;24:79–91. doi: 10.1002/hbm.20070. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Fiebach C, Friederici AD, Muller K, von Cramon DY. fMRI evidence for dual routes to the mental lexicon in visual word recognition. Journal of Cognitive Neuroscience. 2002;14:11–23. doi: 10.1162/089892902317205285. [DOI] [PubMed] [Google Scholar]
- 25.Fischler I, Childers DG, Achariyapaopan T, Perry NW., Jr Brain potentials during sentence verification: automatic aspects of comprehension. Biological Psychology. 1985;21:83–106. doi: 10.1016/0301-0511(85)90008-0. [DOI] [PubMed] [Google Scholar]
- 26.Forman SD, Cohen JD, Fitzgerald M, Eddy WF, Mintun MA, Noll DC. Improved assessment of significant activation in functional magnetic resonance imaging (fMRI): Use of a cluster-size threshold. Magnetic Resonance in Medicine. 1995;33:636–647. doi: 10.1002/mrm.1910330508. [DOI] [PubMed] [Google Scholar]
- 27.Freedman SE, Forster KL. The psychological status of overgenerated sentences. Cognition. 1985;19:101–132. doi: 10.1016/0010-0277(85)90015-0. [DOI] [PubMed] [Google Scholar]
- 28.Friederici AD, Hahne A, Mecklinger A. Temporal structure of syntactic parsing: early and late event-related brain potential effects. J Exper Psychol, Learn, Mem, Cogn. 1996;22:1219–1248. doi: 10.1037//0278-7393.22.5.1219. [DOI] [PubMed] [Google Scholar]
- 29.Friederici AD, Ruschemeyer SA, Hahne A, Fiebach CJ. The Role of left inferior frontal and superior temporal cortex in sentence comprehension: localizing syntactic and semantic processes. Cereb Cortex. 2003;13:170–177. doi: 10.1093/cercor/13.2.170. [DOI] [PubMed] [Google Scholar]
- 30.Friederici AD, Fiebach CJ, Schlesewsky M, Bornkessel ID, von Cramon DY. Processing linguistic complexity and grammaticality in the left frontal cortex. Cereb Cortex. 2006;16:1709–1717. doi: 10.1093/cercor/bhj106. [DOI] [PubMed] [Google Scholar]
- 31.Frisch S, Kotz SA, von Cramon DY, Friederici AD. Why the P600 is not just a P300: The role of the basal ganglia. Clinical Neurophysiology. 2003;114:336–340. doi: 10.1016/s1388-2457(02)00366-8. [DOI] [PubMed] [Google Scholar]
- 32.Grewe T, Bornkessel I, Zysset S, Wiese R, von Vramon DY, Schlesewsky M. Linguistic prominence and Broca’s area: the influence of animacy as a linearization principle. NeuroImage. 2006;32:1395–1402. doi: 10.1016/j.neuroimage.2006.04.213. [DOI] [PubMed] [Google Scholar]
- 33.Grewe T, Bornkessel-Schlesewsky I, Zysset S, Wiese R, von Cramon DY, Schlesewsky M. The role of the posterior superior temporal sulcus in the processing of unmarked transitivity. Neuroimage. 2007;35:343–52. doi: 10.1016/j.neuroimage.2006.11.045. [DOI] [PubMed] [Google Scholar]
- 34.Grodzinsky Y, Friederici AD. Neuroimaging of syntax and syntactic processing. Current Opinion in Neurobiology. 2006;16:240–6. doi: 10.1016/j.conb.2006.03.007. [DOI] [PubMed] [Google Scholar]
- 35.Grossman M, Cooke A, DeVita C, Lee C, Alsop D, Detre J, Gee J, Chen W, Stern MB, Hurtig HI. Grammatical and resource compoents of sentence processing in Parkinson’s disease. Neurology. 2003;60:775–781. doi: 10.1212/01.wnl.0000044398.73241.13. [DOI] [PubMed] [Google Scholar]
- 36.Hagoort P, van Berkum JJA. Beyond the sentence given. Philosophical Transactions of the Royal Society B. 2007;362:801–11. doi: 10.1098/rstb.2007.2089. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Hagoort P, Brown C, Groothusen J. The syntactic positive shift (SPS) as an ERP measure of syntactic processing. In: Garnsey SM, editor. Language and Cognitive Processing Special Issue: Event-related Brain Potentials in the Study of Language. 4. Vol. 8. Lawrence Erlbaum Associates; Hove: 1993. pp. 439–483. [Google Scholar]
- 38.Hagoort P, Hald L, Bastiaansen M, Petersson KM. Integration of word meaning and world knowledge in language comprehension. Science. 2004;304:438–441. doi: 10.1126/science.1095455. [DOI] [PubMed] [Google Scholar]
- 39.Hoeks JCJ, Stowe LA, Doedens G. Seeing words in context: the interaction of lexical and sentence level information during reading. Cognitive Brain Research. 2004;19:59–73. doi: 10.1016/j.cogbrainres.2003.10.022. [DOI] [PubMed] [Google Scholar]
- 40.Jackendoff R. Foundations of language. Oxford, Great Britain: Oxford University Press; 2002. [Google Scholar]
- 41.Just MA, Carpenter PA, Keller TA, Eddy WF, Thulborn KR. Brain activation modulated by sentence comprehension. Science. 1996;274:114–116. doi: 10.1126/science.274.5284.114. [DOI] [PubMed] [Google Scholar]
- 42.Kable JW, Lease-Spellmeyer J, Chatterjee A. Neural substrates of action event knowledge. J Cogn Neurosci. 2002;14:795–805. doi: 10.1162/08989290260138681. [DOI] [PubMed] [Google Scholar]
- 43.Kamide Y, Altmann GTM, Haywood SL. The time-course of prediction in incremental sentence processing: Evidence from anticipatory eye movements. Journal of Memory and Language. 2003;49:133–156. [Google Scholar]
- 44.Keller TA, Carpenter PA, Just MA. The neural bases of sentence comprehension: A fMRI examination of syntactic and lexical processing. Cereb Cortex. 2001;11:223–237. doi: 10.1093/cercor/11.3.223. [DOI] [PubMed] [Google Scholar]
- 45.Kim A, Osterhout L. The independence of combinatory semantic processing: evidence from event-related potentials. Journal of Memory and Language. 2005;52:205–225. [Google Scholar]
- 46.King J, Just MA. Individual differences in syntactic processing: The role of working memory. Journal of Memory and Language. 1991;30:580–602. [Google Scholar]
- 47.Knutson KM, Wood JN, Grafman J. Brain Activation in Processing Temporal Sequence: an fMRI Study. NeuroImage. 2004;23(4):1299–1307. doi: 10.1016/j.neuroimage.2004.08.012. [DOI] [PubMed] [Google Scholar]
- 48.Knutson KM, Wood JN, Spampinato &, Grafman J. Politics on the Brain: An fMRI Investigation. Social Neuroscience. 2006;1(1):25–40. doi: 10.1080/17470910600670603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Kolk HHJ, Chwilla DJ, van Herten M, Oor PJ. Structure and limited capacity in verbal working memory: a study with event-related potentials. Brain and Language. 2003;85:1–36. doi: 10.1016/s0093-934x(02)00548-5. [DOI] [PubMed] [Google Scholar]
- 50.Konishi S, Nakajima K, Uchida I, Sekihara K, Miyashita Y. No-go dominant brain activity in human inferior prefrontal cortex revealed by functional magnetic resonance imaging. European Journal of Neuroscience. 1998;10:1209–1213. doi: 10.1046/j.1460-9568.1998.00167.x. [DOI] [PubMed] [Google Scholar]
- 51.Kruggel F, von Cramon DY. Modeling the hemodynamic response in single-trial functional MRI experiments. Magn Reson Med. 1999;42:787–797. doi: 10.1002/(sici)1522-2594(199910)42:4<787::aid-mrm22>3.0.co;2-v. [DOI] [PubMed] [Google Scholar]
- 52.Kuperberg GR, Kreher DA, Sitnikova T, et al. The role of animacy and thematic relationships in processing active English sentences: Evidence from event-related potentials. Brain and Language. 2007;100:223–237. doi: 10.1016/j.bandl.2005.12.006. [DOI] [PubMed] [Google Scholar]
- 53.Kuperberg GR, Caplan D, Sitnikova T, Eddy M, Holcomb P. Neural correlates of processing syntactic, semantic and thematic relationships in sentences. Lang Cogn Processes. 2006;21:489–530. [Google Scholar]
- 54.Kuperberg GR, Kreher DA, Blais K, et al. Semantic influences on syntactic processing: evidence from event-related potentials. Journal of Cognitive Neuroscience. 2005 Suppl Abstract. [Google Scholar]
- 55.Kuperberg GR, McGuire PK, Bullmore ET, Brammer MJ, Rabe-Hesketh S, Wright IC, Lythgoe DJ, Williams SCR, David AS. Common and distinct neural substrates for pragmatic, semantic, and syntactic processing of spoken sentences: An fMRI study. Journal of Cognitive Neuroscience. 2000;12:321–341. doi: 10.1162/089892900562138. [DOI] [PubMed] [Google Scholar]
- 56.Kutas M, Federmeier KD. Electrophysiology reveals semantic memory use in langauge comprehension. Trends in Cognitive Sciences. 2000;4:463–470. doi: 10.1016/s1364-6613(00)01560-6. [DOI] [PubMed] [Google Scholar]
- 57.Lee D, Marks B, Newman SD. A comparison of on-line and off-line processing during sentence comprehension. Presented at CUNY Sentence Processing Conference; Chapel Hill, NC. 2008. [Google Scholar]
- 58.Lee D, Newman SD. The effect of presentation paradigm on syntactic processing: An event-related fMRI study. Human Brain Mapping. 2010;31:65–79. doi: 10.1002/hbm.20845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Levelt WJM. Models of word production. Trends in Cognitive Sciences. 1999;3:223–233. doi: 10.1016/s1364-6613(99)01319-4. [DOI] [PubMed] [Google Scholar]
- 60.Lieberman P, Kako E, Friedman J, Tajchman G, Feldman LS, Jiminez EB. Speech production, syntax comprehension, and cognitive deficits in Parkinson’s disease. Brain and Language. 1992;43:169–189. doi: 10.1016/0093-934x(92)90127-z. [DOI] [PubMed] [Google Scholar]
- 61.Murray WS, Rowan M. Early, mandatory, pragmatic processing. Journal of Psycholinguistic Research. 1998;27:1–22. [Google Scholar]
- 62.Nadeau SE, Crosson B. Subcortical aphasia. Brain and Language. 1997;58:355–402. doi: 10.1006/brln.1997.1707. [DOI] [PubMed] [Google Scholar]
- 63.Newman SD, Just MA, Keller TA, Roth J, Carpenter PA. Differential effects of syntactic and semantic processing on the subregions of Broca’s area. Brain Res Cogn Brain Res. 2003;16:297–307. doi: 10.1016/s0926-6410(02)00285-9. [DOI] [PubMed] [Google Scholar]
- 64.Newman SD, Lee D, Ratliff K. How much does the comprehension probe interact with on-line syntactic processing? Human Brain Mapping. 2009;30:2499–2511. doi: 10.1002/hbm.20684. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Ni W, Constable RT, Mencl WE, Pugh KR, Fulbright RK, Shaywitz SE, Shaywitz BA, Gore JC, Shankweiler D. An event-related neuroimaging study distinguishing form and content in sentence processing. Journal of Cognitive Neuroscience. 2000;12:120–133. doi: 10.1162/08989290051137648. [DOI] [PubMed] [Google Scholar]
- 66.Poldrack RA, Wagner AD, Prull MW, Desmond JE, Glover GH, Gabrieli JDE. Functional specialization for semantic and phonological processing in the left inferior prefrontal cortex. NeuroImage. 1999;10:15–35. doi: 10.1006/nimg.1999.0441. [DOI] [PubMed] [Google Scholar]
- 67.Price CJ, Moore CJ, Humphreys GW, Wise RJS. Segregating semantic from phonological processes during reading. Journal of Cognitive Neuroscience. 1997;9:727–734. doi: 10.1162/jocn.1997.9.6.727. [DOI] [PubMed] [Google Scholar]
- 68.Smith EE, Jonides J. Storage and executive processes in the frontal lobes. Science. 1999;283:1657–1661. doi: 10.1126/science.283.5408.1657. [DOI] [PubMed] [Google Scholar]
- 69.Tenenhaus MK, Carlson GN, Trueswell JC. The role of thematic structures in interpretation and parsing. Language and Cognitive Processes. 1989;4:211–234. [Google Scholar]
- 70.Townsend DJ, Bever TG. Sentence Comprehension: The integration of habits and rules. MA: MIT Press; 2001. [Google Scholar]
- 71.Wagner AD, Koutstaal W, Maril A, Schacter DL, Buckner RL. Task-specific repetition priming in left inferior prefrontal cortex. Cerebral Cortex. 2000;10:1176–1184. doi: 10.1093/cercor/10.12.1176. [DOI] [PubMed] [Google Scholar]
- 72.Wallesch CW, Papagno C. Subcortical aphasia. In: Rose FC, Whurr R, Wyke MA, editors. Aphasia. London: Whurr Publishers; 1988. [Google Scholar]
- 73.Wood JN, Romero SG, Makale M, Grafman J. Category-specific representations of social and nonsocial knowledge in the human prefrontal cortex. Journal of Cognitive Neuroscience. 2003;15:236–248. doi: 10.1162/089892903321208178. [DOI] [PubMed] [Google Scholar]
- 74.Wu DH, Waller S, Chatterjee A. The functional neuroanatomy of thematic role and locative relational knowledge. J Cogn Neurosci. 2007;19:1542–1555. doi: 10.1162/jocn.2007.19.9.1542. [DOI] [PubMed] [Google Scholar]
- 75.Yekutieli D, Benjamini Y. Resampling-based false discovery rate controlling multiple test procedures for correlated test statistics. Journal of Statistical Planning and Inference. 1999;82:171–196. [Google Scholar]
- 76.Zarahn E, Aguirre G, D’Esposito M. A trial-based experimental design for fMRI. Neuroimage. 1997;6:122–138. doi: 10.1006/nimg.1997.0279. [DOI] [PubMed] [Google Scholar]
- 77.Zarahn E. Testing for neural responses during temporal components of trials with BOLD fMRI. Neuroimage. 2000;11:783–796. doi: 10.1006/nimg.2000.0560. [DOI] [PubMed] [Google Scholar]