

Abstract
Purpose of the review
Food allergy (FA), a growing clinical and public health problem in the U.S. and worldwide, is likely determined by multiple environmental and genetic factors. The purpose of this review is to summarize recent advances in FA genetic research.
Recent findings
There is compelling evidence that genetic factors may play a role in FA. However, the specific genetic loci that may modulate individual risk of FA remain to be identified. To date, only a limited number of candidate gene association studies of FA have been reported. Polymorphism(s) in 9 genes have been associated with the incidence of FA or FA severity in at least one study. But most of these findings remain to be replicated in independent populations. In contrast, there are considerable advances in genetics of other allergic diseases such as asthma and atopic dermatitis. While asthma and atopic dermatitis often co-exist with FA, the relevance of their candidate genes to FA remains to be evaluated.
Summary
Genetics in FA is a promising research area but is still in its infancy. More studies are needed to dissect susceptible genes of FA. A genome-wide association approach may serve as a powerful tool to identify novel genes related to FA. Furthermore, the role of gene-environment interaction, gene-gene interaction, and epigenetics in FA remains largely unexplored. Given the complex nature of FA, future studies need to integrate environment, genomics and epigenomics in order to better understand the multi-facet etiology and biological mechanisms of FA.
Keywords: Food allergy, genetics, Allergic diseases
Introduction
Food allergy (FA), defined as an immunoglobulin (Ig) E-mediated hypersensitivity reaction to food, is emerging as a major clinical and public health problem worldwide [1, 2]. It affects approximately 5-8% of children and 1-5% of adults [3-5]. Such prevalence has risen substantially over the past decade, in parallel to the rise in prevalence previously seen for other atopic conditions [1, 2, 6-10]. Despite this, our current understanding on the etiology and biological mechanisms of FA is still incomplete. It is generally believed that FA, like the other allergic diseases such as asthma and atopic dermatitis (AD), is determined by both environmental and genetic factors[11, 12]. The use of genomic information, accelerated by the sequencing of the human genome and the advent of new tools and technologies, raised widespread hope that FA genetics can significantly contribute to the prediction, prevention, and treatment of FA. The main purpose of this review is to summarize current state in FA genetic research and to offer future perspectives.
Diagnosis of Food Allergy in the Research Setting
There is a lack of well-established methods to define FA in large population studies. Currently, various methods have been used, ranging from self-report to double-blind, placebo-controlled food challenge (DBPCFC) [13]. DBPCFC has been promoted as the gold standard for establishing the diagnosis of FA. However, the procedure is laborious, time-consuming, and associated with uncertain risks such as anaphylaxis, and is therefore not routinely performed in the large-scale studies[14]. Other tests such as prick skin tests (PST) and in vitro measurements of food-specific IgE, are more commonly used. These methods, however, do have limitations, with positive predictive value (PPV) of PST being only 50-60%. Some investigators have proposed clinical cut-offs which obviate the need for oral challenges for establishing the diagnosis. Those include proposed values for the wheal size of skin tests[15-17] and for food-specific IgE levels [18-21]. More recently, studies have attempted to combine clinical history, in vivo and/or in vitro testing to predict the likelihood of clinical reactivity[17, 22-24]. To date, there is not a generally accepted definition of FA in large-scale genetic epidemiological research. In this review, we have included all the articles related to genetics of FA (regardless how it was defined) as well as the articles on the genetics of food-specific IgE or skin prick test (SPT).
Familial Aggregation in Food Allergy
Previous studies have documented that family history is a strong risk factor for FA development [25-28]. A survey of the 622 families with probands of peanut allergy in the UK noted an increased prevalence of peanut allergy in succeeding generations. A child has a 7-fold increase in the risk of peanut allergy if he or she has a parent or sibling with peanut allergy [26]. In another study carried out in the UK, the frequency of peanut allergy has been found to be significantly higher in relatives of peanut allergy patients than in the general populations (7% vs. 0.5%)[27]. In our ongoing study in a Chicago cohort, we demonstrated a strong familial aggregation of FA and food allergen sensitization among family members[28].
Heritability of Food Allergy
In a small twin study (58 twin pairs) on peanut allergy, Sicherer et al reported that the concordance rate of peanut allergy (64.3%) among identical twins was significantly higher than that among dizygotic twins (6.8%) [29] and that the estimated heritability of peanut allergy was 81.6%[29]. This finding was similar to results concerning other allergic diseases, such as asthma (87%), atopic dermatitis (74%) and allergic rhinitis (74-82%). In our Chicago family-based cohort, the estimated heritability ranged from 0.15 to 0.35 for different food-specific IgE, all of which were statistically significant [28]. In our recent large twin study in a Chinese population [30], we demonstrated that sensitization to common food allergens were influenced by both genetic and environmental factors. The estimated heritability to peanut sensitization and to shellfish sensitization was 0.51 and 0.54, respectively [30].
Candidate-Gene Studies of Food Allergy
Positional cloning and candidate gene approaches are the two general approaches used to identify candidate genes of complex diseases. To our knowledge, there is no report on the positional cloning of FA. Several small-scale candidate-gene studies of FA have been reported, but with inconsistent results. The reported genes that were associated with FA or FA severity in at least one study were summarized below.
Major Histocompatibility Complex (HLA) Gene Family
Significant associations with peanut allergy were reported previously for the HLA class II DR beta 1(HLA-DRB1), DQ beta 1(HLA-DQB1) and DP beta 1(HLA-DPB1) gene polymorphisms [31]. Significant association with apple allergy were reported for HLA-DRB1*07 allele [32]. Hand et al also observed that the frequency of HLA-Beta*07 and HLA-DRB1*11 were increased in the nut-allergic patients compared to the atopic controls (12.20% vs 3.66%), and the frequency of HLA-DRB*13 and DQB1*06 were both increased in the nut allergy patients compared to both the atopic and blood donor controls. However, none of those increased frequencies were statistically significant after adjustment of multiple testing [33]. Shreffler et al showed that the genetic polymorphisms in the HLA class II gene family had no associations with peanut allergy in 73 children and 75 of their siblings [34].
CD14 Gene
Another gene of interest for FA is CD14 gene, which is the receptor for lipopolysaccharides. In a study comprised of 175 asthmatic and 77 food-allergic patients with varying ages, the C-159T polymorphism in the promoter of CD14 was found to be associated with non-atopic asthma and FA subjects, particularly in white subjects[35]. However, another study reported an inconsistent result, which found no association between two polymorphisms (C-159T and C-550T) in the promoter of CD14 and FA [36].
FOXP3 Gene
The expression of Forkhead box P3 (FOXP3), a member of the forkhead/winged-helix family of transcriptional regulators, has been thought to be the best marker for naturally occurring regulatory T-cells. Torgerson et al reported that a 1300-base pair deletion in the non-coding region of the FOXP3 gene could lead to low FOXP3 mRNA expression levels and significantly decreased protein expression in peripheral blood lymphocytes [37]. They also observed that this gene variant could cause severe food allergy as a variant IPEX syndrome[37].
STAT6 Gene
Signal transducer and activator of transcription (STAT6) is a central molecule in the signal transduction pathway regulated by IL-4 and IL-13 in IgE isotype switching and production of TH2 cytokines[38, 39]. Amoli et al reported that the G allele in the STAT6 G2964A polymorphism was significantly increased in nut-allergic patients compared with controls under a recessive model [40]. They also found that this polymorphism is associated with severity in nut-allergic patients[40]. However, Negoro et al found no association between this SNP and the severity of food allergy in 220 Japanese allergic children [41].
SPINK5 Gene
Serine protease inhibitor Karzal type 5 (SPINK5) is a protease inhibitor protein. It has been reported that SPINK5 can be expressed in the thymus and its defects have been suggested to cause abnormal maturation of T lymphocytes and leading to Th2 responses such as increased IgE level and eosinophilia [42]. A recent report in Japanese children with atopic dermatitis (AD) showed that patients with the SPINK5 1258AA or 1258GG genotype displayed a significantly higher prevalence of FA[43].
Interleukin 10 (IL10) Gene
IL10 down-regulates the expression of Th1 cytokines, MHC class II Ags, and costimulatory molecules on macrophages. Two SNPs in this gene, A-1082G and C-627A, lie on the putative transcription factor-binding sites and are associated with the production of this cytokine [44, 45]. Negoro et al reported that IL10 -627AA polymorphism was significantly associated with the severity of both FA and AD in 220 Japanese allergic children. Recently, in another study of atopic Japanese children, the authors observed no association between IL10 -627AA and the prevalence of FA. However, they reported that Children carrying the IL10 -1082AA genotype were significantly associated with 2.5 times higher risk of FA [46].
Interleukin 13 (IL13) Gene
IL13 is an important immunoregulatory cytokine produced primarily by activated Th2 cells. IL13 gene polymorphisms have been linked to asthma by more than 25 studies [47]. A recent study in unrelated German children drawn from the multicenter atopic study also showed that the C-1055T polymorphism in the IL13 gene is associated with increased risk of food sensitization [48].
Candidate-Gene Studies of Other Allergic Diseases
FA is strongly associated with certain allergic diseases such as asthma and atopic dermatitis (AD) [49-51]. Our studies in Chinese twin cohorts have shown that allergen sensitizations (including both food allergens and aero-allergens) might be contributed by some common genetic factors, suggesting some common genes shared by FA and the other allergic phenotypes [30]. Among all of the allergic phenotypes, asthma is one of the most studied allergic phenotype in genetic research. Some excellent reviews have summarized current advances in asthma genetics [47, 52]. Briefly, more than 100 genes have been linked to asthma or asthma-related phenotypes in at least one population and 33 genes have been validated in more than five independent populations [47].
To our knowledge, most of the asthma candidate genes are involved in IgE synthesis, innate immunity, allergic inflammation, and/or hyperreactivity of the cells and organs, which are the common pathways shared by multiple allergic diseases including asthma, AD and FA. The current findings have supported that these genes may be also associated with some other allergic phenotypes. For example, five genes, including the mast cell chymase 1 gene (CMA1), IL13, the interleukin 4 receptor gene(IL4R), SPINK5, and the filaggrin gene(FLG), are not only among the most validated genes for asthma, but also have been linked to AD in at least two or more independent studies[53]. Meanwhile, the membrane-spanning 4-domains, subfamily A, member 2 gene (MS4A2) variants were significantly associated with asthma[54, 55], total IgE [54, 56, 57], specific IgE to grass pollen allergen [54], nasal allergy [57] and atopy [58]. Additionally, IL18 genetic variants were significantly associated with asthma[59], AD[60], total IgE [61], specific sensitization to common allergens, and seasonal allergic rhinitis[61]. N-acetyltransferase 2 (NAT2) genetic variants were significantly associated with asthma[62], total serum IgE[62], blood eosinophil counts[62], and allergy (defined as inhalational, food or mixed allergy)[63]. The cytotoxic T-lymphocyte antigen 4 gene (CTLA4) variants were significantly associated with asthma[64], total IgE[64], allergy[64], AD[65], and cord blood IgE[66]. Conceivably, asthma candidate genes, especially those validated in multiple populations, may also serve as candidate genes for food allergy. However, the relevance of these candidate genes to FA remains to be evaluated.
Genome-Wide Associations (GWA) of Allergic Diseases
GWA studies rely on searching for common genetic variants in dense sets of SNPs across the genome that are risk factors for diseases of interest [67]. As compared to candidate gene approach, the main strength of GWA studies lies in their ability to discover truly novel disease candidate genes (not relying on previous knowledge), especially those associated with moderate disease risks. [68]. Although no published GWA studies of FA are currently available, some progress has been made for other allergic phenotypes. Moffatt et al genotyped more than 317,000 SNPs in 994 patients with childhood asthma and 1243 subjects without asthma and identified that genetic variants regulating ORMDL3 expression are determinants of childhood asthma[69]. In a GWA study of 403 asthma families, Murphy et al identified two significant SNPs in chromosomes 1 (rs10863712) and 14 (rs1294497) for childhood asthma[70]. Esparza-Gordillo et al reported the first GWAs study of AD in 939 cases and 975 controls as well as 270 complete nuclear families [71]. Their study identified that subjects carrying AA genotypes in the rs7927894 had a 1.47 times higher risk of developing AD, comparing to those noncarriers[71]. Also, two large-scale GWA studies on allergic-related quantitative traits have been reported. One was conducted using 9,392 Icelanders using Illumina 317K SNP, in which 5 SNPs (including rs1420101, rs12619285, rs4857855, rs4143832 and rs3184504) were significantly associated with blood eosinophil counts[72]. The other was conducted in 1,530 population-based individuals testing 353,569 SNPs, which identified FCER1A as the novel susceptibility locus for total serum IgE levels. These GWA studies not only shed light on the further genetic studies of FA, but also underscore the promise of GWAS in dissecting genetics of FA.
Gene-Environment (G×E) and Gene-Gene (G×G) Interactions
Gene-environment (G×E) interactions, which reflect the complex interplay between environmental exposure and genetic predisposition, have received increasing attention. Studies in mice showed that G×E interactions explain a proportion of the phenotypic variance that may be higher than the main effect explained by either genetic or environmental effects considered separately[73]. Currently, research of G×E interactions on allergic diseases is limited, mainly focusing on three environmental factors, including smoking, air pollution and microbial exposures. Specifically, the GSTP1 Ile105Val polymorphism would modify the effect of air pollution on allergic sensitization to inhalant and/or food allergens[74]. The influence of the C-159T polymorphism on the CD14 gene seems to depend on environmental sources of microbial simulation[75-79]. Additionally, subjects carrying the CD14 TT genotype of the C-159T polymorphism was noted to increase the protective effect of dog exposure on AD[75] and to modify the effect of farm exposures on atopy[76].
Gene-gene interactions, which are the functional interplay between genetic variants, are also likely to contribute to the complexity of human diseases[47]. Current evidence in the genetics of allergic phenotypes and/or food allergy has strongly suggested significant G×G interactions among genes involving the Th2-cell differentiation and signaling pathways. For example, a large study in German children showed that combining polymorphisms in IL4, IL13, IL4RA, and STAT6 genes leads to a 10.8 fold increase in the risk for high serum IgE levels and a 16.8 fold increase in the risk for development of asthma, compared with the effect of any single SNP[80]. Liu et al reported that, among unrelated German children from a multicenter atopy study, the effect of IL13-1112TT genotype on food sensitization was modified by polymorphisms in the IL4RA gene[48]. Currently, the roles of G×G and G×E interactions in FA development remain largely unexplored, and represent a promising area of research.
Epigenetics
Epigenetics is broadly defined as changes in gene expression patterns that can be inherited and are independent of changes in DNA sequences. Epigenetic alterations are believed to occur not only prenatally or shortly after birth, but also during later developmental periods, influencing gene expression differentially throughout the lifespan. As such, epigenetic regulation provides an attractive mechanistic explanation for some molecular events linking early exposure to later disease development [81]. Particularly, DNA methylation and chromatin modifications are the two major mechanisms that are most studied in epigenetic research.
Previous data has suggested that epigenetics plays an important role in Th cell differentiation and cytokine gene expression[82-86], leading to the conclusion that both are important biological pathways of FA. For example, Jones et al reported that methylation on the -53 CpG site in the IFN-r promoter suppressed IFN-r transcription by inhibiting cAMP response element binding protein (CREB), and thus, was associated with Th2 polarization [84]. By using an animal model, Liu et al found that altering methylation of IL-4 and IFN-r promoter regions by inhaling environmental exposures was significantly correlated with changes in IgE levels [85]. In addition, Su et al found that inhibition of endogenous histone deacetylase (HDAC) activity shifted Th1:Th2 ratios by 3-fold to 8-fold, skewing recall responses toward a more Th2-like phenotype[86]. We anticipate that further studies on how epigenetic alternation affects the risk of FA and how these epigenetic alternations induced by environmental exposure hold great promise to clarify the mechanisms concerning how early exposure affects the development of FA and to identify early biomarkers to effectively predict and prevent the incidence of FA.
Conclusion
FA is likely the results of complex interplay of a large number of genetic and environmental factors. Given our limited understanding of the complex biological pathways and mediators involved in FA, GWA studies with adequate design and power to detect G×E and G×G interactions may represent the best available approach to discover a novel set of genes related to FA. One challenge but promising area in FA research is how to integrate environment, genomics and epigenomics to better understand the complex etiology and biological mechanisms of FA (Figure 1). Such study would require ascertainment of phenotypic, environmental, genomic, and epigenomic data in a same study population with sufficient sample size. It would also require strong interdisciplinary collaboration and adequate funding support. It is anticipated that the advances in FA genetics and epigenetics will help us better understand the molecular processes involved in various pathways of FA and, ultimately, lead to more effective prevention and treatment of FA.
Figure 1.
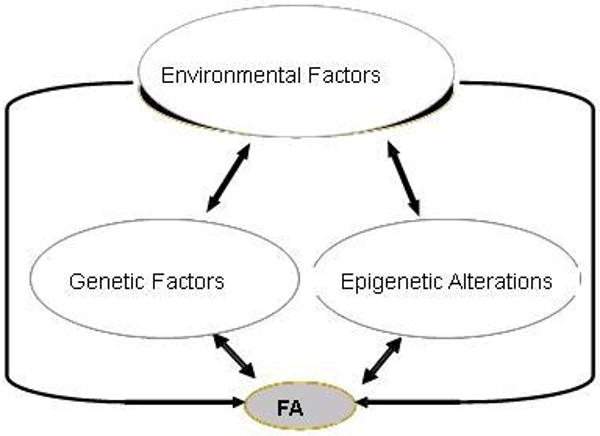
Interplay of environment, genetics and epigenetics in the development of food allergy
Acknowledgments
This work is supported in part by the Food Allergy Project/Food Allergy Initiative, The Chicago Community Trust (C2007-01166); and NIAID (R21AI079872; R56AI080627).
Footnotes
All the authors have no conflicts of interest.
References
- 1.Sampson HA. Update on Food Allergy. J Allergy Clin Immunol. 2004;113:805–819. doi: 10.1016/j.jaci.2004.03.014. quiz 820. [DOI] [PubMed] [Google Scholar]
- 2.Gupta R, Sheikh A, Strachan DP, Anderson HR. Time Trends in Allergic Disorders in the Uk. Thorax. 2007;62:91–96. doi: 10.1136/thx.2004.038844. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Bock SA. Prospective Appraisal of Complaints of Adverse Reactions to Foods in Children During the First 3 Years of Life. Pediatrics. 1987;79:683–688. [PubMed] [Google Scholar]
- 4.Sicherer SH, Sampson HA. 9. Food Allergy. J Allergy Clin Immunol. 2006;117:S470–475. doi: 10.1016/j.jaci.2005.05.048. [DOI] [PubMed] [Google Scholar]
- 5.Vierk KA, Koehler KM, Fein SB, Street DA. Prevalence of Self-Reported Food Allergy in American Adults and Use of Food Labels. J Allergy Clin Immunol. 2007;119:1504–1510. doi: 10.1016/j.jaci.2007.03.011. [DOI] [PubMed] [Google Scholar]
- 6.Ninan TK, Russell G. Respiratory Symptoms and Atopy in Aberdeen Schoolchildren: Evidence from Two Surveys 25 Years Apart. Bmj. 1992;304:873–875. doi: 10.1136/bmj.304.6831.873. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Peat JK, van den Berg RH, Green WF, et al. Changing Prevalence of Asthma in Australian Children. Bmj. 1994;308:1591–1596. doi: 10.1136/bmj.308.6944.1591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Sears MR, Holdaway MD, Flannery EM, et al. Parental and Neonatal Risk Factors for Atopy, Airway Hyper-Responsiveness, and Asthma. Arch Dis Child. 1996;75:392–398. doi: 10.1136/adc.75.5.392. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Sicherer SH. Clinical Update on Peanut Allergy. Ann Allergy Asthma Immunol. 2002;88:350–361. doi: 10.1016/S1081-1206(10)62363-0. quiz 361-352, 394. [DOI] [PubMed] [Google Scholar]
- 10.Grundy J, Matthews S, Bateman B, et al. Rising Prevalence of Allergy to Peanut in Children: Data from 2 Sequential Cohorts. J Allergy Clin Immunol. 2002;110:784–789. doi: 10.1067/mai.2002.128802. [DOI] [PubMed] [Google Scholar]
- 11.Bjorksten B. Genetic and Environmental Risk Factors for the Development of Food Allergy. Curr Opin Allergy Clin Immunol. 2005;5:249–253. doi: 10.1097/01.all.0000168790.82206.17. [DOI] [PubMed] [Google Scholar]
- 12*.Lack G. Epidemiologic Risks for Food Allergy. J Allergy Clin Immunol. 2008;121:1331–1336. doi: 10.1016/j.jaci.2008.04.032. [DOI] [PubMed] [Google Scholar]; It is an excellent review on the possible risk factors and theories for the development of food allergy.
- 13.Rona RJ, Keil T, Summers C, et al. The Prevalence of Food Allergy: A Meta-Analysis. J Allergy Clin Immunol. 2007;120:638–646. doi: 10.1016/j.jaci.2007.05.026. [DOI] [PubMed] [Google Scholar]
- 14.Martelli A, Bouygue GR, Isoardi P, et al. Oral Food Challenges in Children in Italy. Allergy. 2005;60:907–911. doi: 10.1111/j.1398-9995.2005.00777.x. [DOI] [PubMed] [Google Scholar]
- 15.Sporik R, Hill DJ, Hosking CS. Specificity of Allergen Skin Testing in Predicting Positive Open Food Challenges to Milk, Egg and Peanut in Children. Clin Exp Allergy. 2000;30:1540–1546. doi: 10.1046/j.1365-2222.2000.00928.x. [DOI] [PubMed] [Google Scholar]
- 16.Muraro A, Dreborg S, Halken S, et al. Dietary Prevention of Allergic Diseases in Infants and Small Children. Part Iii: Critical Review of Published Peer-Reviewed Observational and Interventional Studies and Final Recommendations. Pediatr Allergy Immunol. 2004;15:291–307. doi: 10.1111/j.1399-3038.2004.00127.x. [DOI] [PubMed] [Google Scholar]
- 17.Venter C, Pereira B, Grundy J, et al. Incidence of Parentally Reported and Clinically Diagnosed Food Hypersensitivity in the First Year of Life. J Allergy Clin Immunol. 2006;117:1118–1124. doi: 10.1016/j.jaci.2005.12.1352. [DOI] [PubMed] [Google Scholar]
- 18.Sampson HA. Utility of Food-Specific Ige Concentrations in Predicting Symptomatic Food Allergy. J Allergy Clin Immunol. 2001;107:891–896. doi: 10.1067/mai.2001.114708. [DOI] [PubMed] [Google Scholar]
- 19.Boyano Martinez T, Garcia-Ara C, Diaz-Pena JM, et al. Validity of Specific Ige Antibodies in Children with Egg Allergy. Clin Exp Allergy. 2001;31:1464–1469. doi: 10.1046/j.1365-2222.2001.01175.x. [DOI] [PubMed] [Google Scholar]
- 20.Garcia-Ara C, Boyano-Martinez T, Diaz-Pena JM, et al. Specific Ige Levels in the Diagnosis of Immediate Hypersensitivity to Cows' Milk Protein in the Infant. J Allergy Clin Immunol. 2001;107:185–190. doi: 10.1067/mai.2001.111592. [DOI] [PubMed] [Google Scholar]
- 21.Clark AT, Ewan PW. Interpretation of Tests for Nut Allergy in One Thousand Patients, in Relation to Allergy or Tolerance. Clin Exp Allergy. 2003;33:1041–1045. doi: 10.1046/j.1365-2745.2003.01624.x. [DOI] [PubMed] [Google Scholar]
- 22.Perry TT, Matsui EC, Kay Conover-Walker M, Wood RA. The Relationship of Allergen-Specific Ige Levels and Oral Food Challenge Outcome. J Allergy Clin Immunol. 2004;114:144–149. doi: 10.1016/j.jaci.2004.04.009. [DOI] [PubMed] [Google Scholar]
- 23.Verstege A, Mehl A, Rolinck-Werninghaus C, et al. The Predictive Value of the Skin Prick Test Weal Size for the Outcome of Oral Food Challenges. Clin Exp Allergy. 2005;35:1220–1226. doi: 10.1111/j.1365-2222.2005.2324.x. [DOI] [PubMed] [Google Scholar]
- 24.Ridout S, Matthews S, Gant C, et al. The Diagnosis of Brazil Nut Allergy Using History, Skin Prick Tests, Serum-Specific Immunoglobulin E and Food Challenges. Clin Exp Allergy. 2006;36:226–232. doi: 10.1111/j.1365-2222.2006.02426.x. [DOI] [PubMed] [Google Scholar]
- 25.Hourihane JO, Warner JO. Allergy to Peanut. Lancet. 1996;348:1523. doi: 10.1016/s0140-6736(05)65947-8. [DOI] [PubMed] [Google Scholar]
- 26.Hourihane JO, Dean TP, Warner JO. Peanut Allergy in Relation to Heredity, Maternal Diet, and Other Atopic Diseases: Results of a Questionnaire Survey, Skin Prick Testing, and Food Challenges. Bmj. 1996;313:518–521. doi: 10.1136/bmj.313.7056.518. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Crespo JF, James JM, Fernandez-Rodriguez C, Rodriguez J. Food Allergy: Nuts and Tree Nuts. Br J Nutr. 2006;96 2:S95–102. doi: 10.1017/bjn20061869. [DOI] [PubMed] [Google Scholar]
- 28*.Tsai HJ, Kumar R, Pongracic J, et al. Familial Aggregation of Food Allergy and Sensitization to Food Allergens: A Family-Based Study. Clin Exp Allergy. 2009;39:101–109. doi: 10.1111/j.1365-2222.2008.03111.x. [DOI] [PMC free article] [PubMed] [Google Scholar]; The article demonstrated strong familial aggregation and indicated genetic susceptibility of food allergy and sensitization in US population.
- 29.Sicherer SH, Furlong TJ, Maes HH, et al. Genetics of Peanut Allergy: A Twin Study. J Allergy Clin Immunol. 2000;106:53–56. doi: 10.1067/mai.2000.108105. [DOI] [PubMed] [Google Scholar]
- 30**.Liu X, Zhang SC, Tsai HJ, et al. Genetic and Environmental Contributions to Allergen Sensitization in a Chinese Twin Study. Clinical and Experimental Allergy. doi: 10.1111/j.1365-2222.2009.03228.x. in press. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study suggested that sensitization to common food and aeroallergens were influenced by both genetic and environmental factors and that paired allergen sensitizations were contributed by both shared genetic and environmental factor, which underscoring the need to further delineate unique and/or pleiotropic genetic and environmental factors for allergen sensitization.
- 31.Howell WM, Turner SJ, Hourihane JO, et al. Hla Class Ii Drb1, Dqb1 and Dpb1 Genotypic Associations with Peanut Allergy: Evidence from a Family-Based and Case-Control Study. Clin Exp Allergy. 1998;28:156–162. doi: 10.1046/j.1365-2222.1998.00224.x. [DOI] [PubMed] [Google Scholar]
- 32.Senechal H, Geny S, Desvaux FX, et al. Genetics and Specific Immune Response in Allergy to Birch Pollen and Food: Evidence of a Strong, Positive Association between Atopy and the Hla Class Ii Allele Hla-Dr7. J Allergy Clin Immunol. 1999;104:395–401. doi: 10.1016/s0091-6749(99)70384-2. [DOI] [PubMed] [Google Scholar]
- 33.Hand S, Darke C, Thompson J, et al. Human Leucocyte Antigen Polymorphisms in Nut-Allergic Patients in South Wales. Clin Exp Allergy. 2004;34:720–724. doi: 10.1111/j.1365-2222.2004.1932.x. [DOI] [PubMed] [Google Scholar]
- 34.Shreffler WG, Charlop-Powers Z, Sicherer SH. Lack of Association of Hla Class Ii Alleles with Peanut Allergy. Ann Allergy Asthma Immunol. 2006;96:865–869. doi: 10.1016/S1081-1206(10)61351-8. [DOI] [PubMed] [Google Scholar]
- 35.Woo JG, Assa'ad A, Heizer AB, et al. The -159 C-->T Polymorphism of Cd14 Is Associated with Nonatopic Asthma and Food Allergy. J Allergy Clin Immunol. 2003;112:438–444. doi: 10.1067/mai.2003.1634. [DOI] [PubMed] [Google Scholar]
- 36.Campos E, Shimojo N, Inoue Y, et al. No Association of Polymorphisms in the 5′ Region of the Cd14 Gene and Food Allergy in a Japanese Population. Allergol Int. 2007;56:23–27. doi: 10.2332/allergolint.O-06-432. [DOI] [PubMed] [Google Scholar]
- 37.Torgerson TR, Linane A, Moes N, et al. Severe Food Allergy as a Variant of Ipex Syndrome Caused by a Deletion in a Noncoding Region of the Foxp3 Gene. Gastroenterology. 2007;132:1705–1717. doi: 10.1053/j.gastro.2007.02.044. [DOI] [PubMed] [Google Scholar]
- 38.Takeda K, Tanaka T, Shi W, et al. Essential Role of Stat6 in Il-4 Signalling. Nature. 1996;380:627–630. doi: 10.1038/380627a0. [DOI] [PubMed] [Google Scholar]
- 39.Takeda K, Kamanaka M, Tanaka T, et al. Impaired Il-13-Mediated Functions of Macrophages in Stat6-Deficient Mice. J Immunol. 1996;157:3220–3222. [PubMed] [Google Scholar]
- 40.Amoli MM, Hand S, Hajeer AH, et al. Polymorphism in the Stat6 Gene Encodes Risk for Nut Allergy. Genes Immun. 2002;3:220–224. doi: 10.1038/sj.gene.6363872. [DOI] [PubMed] [Google Scholar]
- 41.Negoro T, Orihara K, Irahara T, et al. Influence of Snps in Cytokine-Related Genes on the Severity of Food Allergy and Atopic Eczema in Children. Pediatr Allergy Immunol. 2006;17:583–590. doi: 10.1111/j.1399-3038.2006.00463.x. [DOI] [PubMed] [Google Scholar]
- 42.Chavanas S, Bodemer C, Rochat A, et al. Mutations in Spink5, Encoding a Serine Protease Inhibitor, Cause Netherton Syndrome. Nat Genet. 2000;25:141–142. doi: 10.1038/75977. [DOI] [PubMed] [Google Scholar]
- 43.Kusunoki T, Okafuji I, Yoshioka T, et al. Spink5 Polymorphism Is Associated with Disease Severity and Food Allergy in Children with Atopic Dermatitis. J Allergy Clin Immunol. 2005;115:636–638. doi: 10.1016/j.jaci.2004.12.1114. [DOI] [PubMed] [Google Scholar]
- 44.Yilmaz V, Yentur SP, Saruhan-Direskeneli G. Il-12 and Il-10 Polymorphisms and Their Effects on Cytokine Production. Cytokine. 2005;30:188–194. doi: 10.1016/j.cyto.2005.01.006. [DOI] [PubMed] [Google Scholar]
- 45.Turner DM, Williams DM, Sankaran D, et al. An Investigation of Polymorphism in the Interleukin-10 Gene Promoter. Eur J Immunogenet. 1997;24:1–8. doi: 10.1111/j.1365-2370.1997.tb00001.x. [DOI] [PubMed] [Google Scholar]
- 46*.Alberto EJ, Shimojo N, Suzuki Y, et al. Il-10 Gene Polymorphism, but Not Tgf-Beta1 Gene Polymorphisms, Is Associated with Food Allergy in a Japanese Population. Pediatr Allergy Immunol. 2008;19:716–721. doi: 10.1111/j.1399-3038.2007.00709.x. [DOI] [PubMed] [Google Scholar]; It provided the first evidence that IL10 A-1082 allele is associated with food allergy in an Asian population.
- 47.Vercelli D. Discovering Susceptibility Genes for Asthma and Allergy. Nat Rev Immunol. 2008;8:169–182. doi: 10.1038/nri2257. [DOI] [PubMed] [Google Scholar]
- 48.Liu X, Beaty TH, Deindl P, et al. Associations between Specific Serum Ige Response and 6 Variants within the Genes Il4, Il13, and Il4ra in German Children: The German Multicenter Atopy Study. J Allergy Clin Immunol. 2004;113:489–495. doi: 10.1016/j.jaci.2003.12.037. [DOI] [PubMed] [Google Scholar]
- 49.Rhodes HL, Sporik R, Thomas P, et al. Early Life Risk Factors for Adult Asthma: A Birth Cohort Study of Subjects at Risk. J Allergy Clin Immunol. 2001;108:720–725. doi: 10.1067/mai.2001.119151. [DOI] [PubMed] [Google Scholar]
- 50.Rhodes HL, Thomas P, Sporik R, et al. A Birth Cohort Study of Subjects at Risk of Atopy: Twenty-Two-Year Follow-up of Wheeze and Atopic Status. Am J Respir Crit Care Med. 2002;165:176–180. doi: 10.1164/ajrccm.165.2.2104032. [DOI] [PubMed] [Google Scholar]
- 51.Gustafsson D, Sjoberg O, Foucard T. Development of Allergies and Asthma in Infants and Young Children with Atopic Dermatitis--a Prospective Follow-up to 7 Years of Age. Allergy. 2000;55:240–245. doi: 10.1034/j.1398-9995.2000.00391.x. [DOI] [PubMed] [Google Scholar]
- 52.Bosse Y, Hudson TJ. Toward a Comprehensive Set of Asthma Susceptibility Genes. Annu Rev Med. 2007;58:171–184. doi: 10.1146/annurev.med.58.071105.111738. [DOI] [PubMed] [Google Scholar]
- 53.Kiyohara C, Tanaka K, Miyake Y. Genetic Susceptibility to Atopic Dermatitis. Allergol Int. 2008;57:39–56. doi: 10.2332/allergolint.R-07-150. [DOI] [PubMed] [Google Scholar]
- 54.Li A, Hopkin JM. Atopy Phenotype in Subjects with Variants of the Beta Subunit of the High Affinity Ige Receptor. Thorax. 1997;52:654–655. doi: 10.1136/thx.52.7.654. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hizawa N, Maeda Y, Konno S, et al. Genetic Polymorphisms at Fcer1b and Pai-1 and Asthma Susceptibility. Clin Exp Allergy. 2006;36:872–876. doi: 10.1111/j.1365-2222.2006.02413.x. [DOI] [PubMed] [Google Scholar]
- 56.Shirakawa T, Li A, Dubowitz M, et al. Association between Atopy and Variants of the Beta Subunit of the High-Affinity Immunoglobulin E Receptor. Nat Genet. 1994;7:125–129. doi: 10.1038/ng0694-125. [DOI] [PubMed] [Google Scholar]
- 57.Nagata H, Mutoh H, Kumahara K, et al. Association between Nasal Allergy and a Coding Variant of the Fc Epsilon Ri Beta Gene Glu237gly in a Japanese Population. Hum Genet. 2001;109:262–266. doi: 10.1007/s004390100561. [DOI] [PubMed] [Google Scholar]
- 58.Hill MR, James AL, Faux JA, et al. Fc Epsilon Ri-Beta Polymorphism and Risk of Atopy in a General Population Sample. Bmj. 1995;311:776–779. doi: 10.1136/bmj.311.7008.776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Imboden M, Nicod L, Nieters A, et al. The Common G-Allele of Interleukin-18 Single-Nucleotide Polymorphism Is a Genetic Risk Factor for Atopic Asthma. The Sapaldia Cohort Study. Clin Exp Allergy. 2006;36:211–218. doi: 10.1111/j.1365-2222.2006.02424.x. [DOI] [PubMed] [Google Scholar]
- 60.Kim E, Lee JE, Namkung JH, et al. Association of the Single-Nucleotide Polymorphism and Haplotype of the Interleukin 18 Gene with Atopic Dermatitis in Koreans. Clin Exp Allergy. 2007;37:865–871. doi: 10.1111/j.1365-2222.2007.02717.x. [DOI] [PubMed] [Google Scholar]
- 61.Kruse S, Kuehr J, Moseler M, et al. Polymorphisms in the Il 18 Gene Are Associated with Specific Sensitization to Common Allergens and Allergic Rhinitis. J Allergy Clin Immunol. 2003;111:117–122. doi: 10.1067/mai.2003.43. [DOI] [PubMed] [Google Scholar]
- 62.Batra J, Sharma SK, Ghosh B. Arylamine N-Acetyltransferase Gene Polymorphisms: Markers for Atopic Asthma, Serum Ige and Blood Eosinophil Counts. Pharmacogenomics. 2006;7:673–682. doi: 10.2217/14622416.7.5.673. [DOI] [PubMed] [Google Scholar]
- 63.Zielinska E, Niewiarowski W, Bodalski J, et al. Arylamine N-Acetyltransferase (Nat2) Gene Mutations in Children with Allergic Diseases. Clin Pharmacol Ther. 1997;62:635–642. doi: 10.1016/S0009-9236(97)90083-7. [DOI] [PubMed] [Google Scholar]
- 64.Howard TD, Postma DS, Hawkins GA, et al. Fine Mapping of an Ige-Controlling Gene on Chromosome 2q: Analysis of Ctla4 and Cd28. J Allergy Clin Immunol. 2002;110:743–751. doi: 10.1067/mai.2002.128723. [DOI] [PubMed] [Google Scholar]
- 65.Jones G, Wu S, Jang N, et al. Polymorphisms within the Ctla4 Gene Are Associated with Infant Atopic Dermatitis. Br J Dermatol. 2006;154:467–471. doi: 10.1111/j.1365-2133.2005.07080.x. [DOI] [PubMed] [Google Scholar]
- 66.Chang JC, Liu CA, Chuang H, et al. Gender-Limited Association of Cytotoxic T-Lymphocyte Antigen-4 (Ctla-4) Polymorphism with Cord Blood Ige Levels. Pediatr Allergy Immunol. 2004;15:506–512. doi: 10.1111/j.1399-3038.2004.00161.x. [DOI] [PubMed] [Google Scholar]
- 67.Hirschhorn JN, Daly MJ. Genome-Wide Association Studies for Common Diseases and Complex Traits. Nat Rev Genet. 2005;6:95–108. doi: 10.1038/nrg1521. [DOI] [PubMed] [Google Scholar]
- 68**.Weidinger S, Gieger C, Rodriguez E, et al. Genome-Wide Scan on Total Serum Ige Levels Identifies Fcer1a as Novel Susceptibility Locus. PLoS Genet. 2008;4:e1000166. doi: 10.1371/journal.pgen.1000166. [DOI] [PMC free article] [PubMed] [Google Scholar]; It is the first GWA study on total serum IgE levels, in which, FCER1A is identified and replicated as a new susceptibility locus influencing serum IgE levels.
- 69.Moffatt MF, Kabesch M, Liang L, et al. Genetic Variants Regulating Ormdl3 Expression Contribute to the Risk of Childhood Asthma. Nature. 2007;448:470–473. doi: 10.1038/nature06014. [DOI] [PubMed] [Google Scholar]
- 70.Murphy A, Weiss ST, Lange C. Screening and Replication Using the Same Data Set: Testing Strategies for Family-Based Studies in Which All Probands Are Affected. PLoS Genet. 2008;4:e1000197. doi: 10.1371/journal.pgen.1000197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71**.Esparza-Gordillo J, Weidinger S, Folster-Holst R, et al. A Common Variant on Chromosome 11q13 Is Associated with Atopic Dermatitis. Nat Genet. 2009;41:596–601. doi: 10.1038/ng.347. [DOI] [PubMed] [Google Scholar]; It is the first GWA study on atopic dermatitis, which demonstrates a highly significant association with A-allele of rs7927894 in all four study groups, including more than 4,100 atopic dermatitis cases. A list of additional candidate genes are also provided in this article.
- 72**.Gudbjartsson DF, Bjornsdottir US, Halapi E, et al. Sequence Variants Affecting Eosinophil Numbers Associate with Asthma and Myocardial Infarction. Nat Genet. 2009;41:342–347. doi: 10.1038/ng.323. [DOI] [PubMed] [Google Scholar]; This GWA study in 9,392 Icelanders has discovered several sequence variants for blood eosinophil counts and three susceptibility loci related to asthma.
- 73.Valdar W, Solberg LC, Gauguier D, et al. Genetic and Environmental Effects on Complex Traits in Mice. Genetics. 2006;174:959–984. doi: 10.1534/genetics.106.060004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74*.Melen E, Nyberg F, Lindgren CM, et al. Interactions between Glutathione S-Transferase P1, Tumor Necrosis Factor, and Traffic-Related Air Pollution for Development of Childhood Allergic Disease. Environ Health Perspect. 2008;116:1077–1084. doi: 10.1289/ehp.11117. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study demonstrated a significant gene-environment interaction between a natural antioxidant enzyme, GSTP1, and a long-term exposure to ambient air pollution from traffic with respect to sensitization to common allergen in Children.
- 75.Gern JE, Reardon CL, Hoffjan S, et al. Effects of Dog Ownership and Genotype on Immune Development and Atopy in Infancy. J Allergy Clin Immunol. 2004;113:307–314. doi: 10.1016/j.jaci.2003.11.017. [DOI] [PubMed] [Google Scholar]
- 76.Leynaert B, Guilloud-Bataille M, Soussan D, et al. Association between Farm Exposure and Atopy, According to the Cd14 C-159t Polymorphism. J Allergy Clin Immunol. 2006;118:658–665. doi: 10.1016/j.jaci.2006.06.015. [DOI] [PubMed] [Google Scholar]
- 77.Eder W, Klimecki W, Yu L, et al. Opposite Effects of Cd 14/-260 on Serum Ige Levels in Children Raised in Different Environments. J Allergy Clin Immunol. 2005;116:601–607. doi: 10.1016/j.jaci.2005.05.003. [DOI] [PubMed] [Google Scholar]
- 78.Ege MJ, Bieli C, Frei R, et al. Prenatal Farm Exposure Is Related to the Expression of Receptors of the Innate Immunity and to Atopic Sensitization in School-Age Children. J Allergy Clin Immunol. 2006;117:817–823. doi: 10.1016/j.jaci.2005.12.1307. [DOI] [PubMed] [Google Scholar]
- 79.Simpson A, John SL, Jury F, et al. Endotoxin Exposure, Cd14, and Allergic Disease: An Interaction between Genes and the Environment. Am J Respir Crit Care Med. 2006;174:386–392. doi: 10.1164/rccm.200509-1380OC. [DOI] [PubMed] [Google Scholar]
- 80.Kabesch M, Schedel M, Carr D, et al. Il-4/Il-13 Pathway Genetics Strongly Influence Serum Ige Levels and Childhood Asthma. J Allergy Clin Immunol. 2006;117:269–274. doi: 10.1016/j.jaci.2005.10.024. [DOI] [PubMed] [Google Scholar]
- 81**.Miller RL, Ho SM. Environmental Epigenetics and Asthma: Current Concepts and Call for Studies. Am J Respir Crit Care Med. 2008;177:567–573. doi: 10.1164/rccm.200710-1511PP. [DOI] [PMC free article] [PubMed] [Google Scholar]; This article has reviewed evidence supporting the hypothesis that asthma risk may be influenced by environmental factors inducing epigenetic changes, summarized the proposed epigenetic mechanisms in the development of asthma, and suggested that the study of environmental epigenetics held a great promise to help us understand a theoretically preventable environmental disease.
- 82.Agarwal S, Rao A. Modulation of Chromatin Structure Regulates Cytokine Gene Expression During T Cell Differentiation. Immunity. 1998;9:765–775. doi: 10.1016/s1074-7613(00)80642-1. [DOI] [PubMed] [Google Scholar]
- 83.Lee DU, Agarwal S, Rao A. Th2 Lineage Commitment and Efficient Il-4 Production Involves Extended Demethylation of the Il-4 Gene. Immunity. 2002;16:649–660. doi: 10.1016/s1074-7613(02)00314-x. [DOI] [PubMed] [Google Scholar]
- 84.Jones B, Chen J. Inhibition of Ifn-Gamma Transcription by Site-Specific Methylation During T Helper Cell Development. Embo J. 2006;25:2443–2452. doi: 10.1038/sj.emboj.7601148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85*.Liu J, Ballaney M, Al-alem U, et al. Combined Inhaled Diesel Exhaust Particles and Allergen Exposure Alter Methylation of T Helper Genes and Ige Production in Vivo. Toxicol Sci. 2008;102:76–81. doi: 10.1093/toxsci/kfm290. [DOI] [PMC free article] [PubMed] [Google Scholar]; This study is the first to demonstrate that inhaled environmental exposures influence methylation of TH genes in vivo, which supports a new paradigm in the development of asthma, and possibly, the other allergic diseases.
- 86**.Su RC, Becker AB, Kozyrskyj AL, Hayglass KT. Epigenetic Regulation of Established Human Type 1 Versus Type 2 Cytokine Responses. J Allergy Clin Immunol. 2008;121:57–63 e53. doi: 10.1016/j.jaci.2007.09.004. [DOI] [PubMed] [Google Scholar]; This study demonstrated that endogenous histone acetylation plays a key regulatory role in maintaining both the intensity and balance of ongoing Th1-associated vs. Th2-assocatied immune response in human beings, which offer an epigenetic regulatory mechanism explaining how environment may influence the maintenance of Th1/Th2 immunity.