

Abstract
Background
Asymptomatic Plasmodium spp. infections and anemia are highly prevalent conditions in tropical regions. We studied whether asymptomatic parasitemia induces hepcidin- and/or cytokine-mediated iron maldistribution and anemia.
Design and Methods
A group of 1197 Indonesian schoolchildren, aged 5–15 years, were screened by microscopy for the presence of parasitemia. Concentrations of hemoglobin, serum hepcidin and parameters of iron status and inflammation were determined at baseline and 4 weeks after antimalarial treatment.
Results
Asymptomatic P. falciparum and P. vivax parasitemia were detected in 73 (6.1%) and 18 (1.5%) children, respectively, of whom 84% and 83% had a C-reactive protein concentration below 5 mg/L. Children with P. falciparum or P. vivax parasitemia had significantly lower hemoglobin concentrations than 17 aparasitemic controls (12.6 and 12.2 g/dL versus 14.4 g/dL; P<0.01), together with significantly higher serum hepcidin concentrations (5.2 and 5.6 nM versus 3.1 nM; P<0.05). The latter was associated with signs of iron maldistribution with higher ferritin concentrations and lower values of serum iron concentration, transferrin saturation and erythrocyte mean cell volume. Concentrations of growth differentiation factor 15 were similar across groups. Antimalarial treatment partly reversed these abnormalities and led to a significant increase in hemoglobin concentration.
Conclusions
Asymptomatic malarial parasitemia is associated with increased hepcidin concentrations and anemia, in the absence of a manifest acute phase response. Prolonged iron maldistribution may be an underestimated cause of anemia. Screening for parasitemia should be performed before starting iron supplementation, as iron therapy may be less effective and even hazardous in these circumstances.
Keywords: asymptomatic parasitemia, hepcidin, Plasmodium spp., infection
Introduction
Infectious diseases induce alterations in the distribution of iron in the human body. Recent findings have shown that many of these alterations are attributable to the actions of the hormone hepcidin.1 Hepcidin inhibits the absorption of iron through enterocytes and impairs iron release by macrophages through degradation of ferroportin, the only iron exporter. Pro-inflammatory cytokines stimulate hepcidin expression, which leads to hypoferremia and increased iron storage in the reticuloendothelial system, both typical features of the anemia of chronic disease.2
Recent advances in the understanding of iron homeostasis and the development of reliable hepcidin assays offer new opportunities to improve iron supplementation practices in regions in which malaria is endemic. Iron deficiency is a common condition in malaria-endemic regions and its complications are not restricted to anemia, but include impaired growth, cognitive development and physical capacity.3 Iron supplementation and iron fortification of foods have, therefore, been considered important public health interventions,4 but there is currently growing concern about the safety of iron supplementation in people exposed to malaria, as iron may increase malaria-associated morbidity and mortality.5,6
So far, studies on iron homeostasis and malaria have largely been restricted to symptomatic P. falciparum malaria. However, asymptomatic P. falciparum parasitemia, i.e. the presence of parasitemia in the absence of fever or malaria-related symptoms, is also an extremely common and chronic condition among (semi-)immune persons in malaria areas, with prevalence rates exceeding 50% in some areas.7,8 Moreover, up to half of all malaria cases outside Africa are caused by P. vivax, which is characterized by a more indolent and chronic course than P. falciparum, although it may also ultimately result in severe anemia.9,10
Previously, our group studied hepcidin expression in anemic Tanzanian children with febrile P. falciparum malaria with high parasite densities and in malaria-naïve Dutch volunteers who were experimentally infected with P. falciparum.11,12 Hepcidin concentrations were increased in both groups and associated with profound disturbances in iron homeostasis, suggesting that iron supplements should be withheld during symptomatic P. falciparum malaria. To our knowledge, no data are yet available on the effects of asymptomatic P. falciparum or P. vivax parasitemia on hepcidin expression. Due to the chronic and relapsing nature of both conditions, disturbances in the uptake, recycling and utilization of iron could play an important and so far under-recognized role in the pathogenesis of anemia in tropical areas and reduce the hemoglobin response to iron supplements and iron fortified foods.
The aim of the present study was, therefore, to determine whether asymptomatic parasitemia induces disturbances in iron homeostasis. Concentrations of serum hepcidin and a range of hematologic, inflammatory and serum iron parameters were studied in schoolchildren with asymptomatic P. falciparum or P. vivax parasitemia on the Indonesian island Sumba, before and after antimalarial treatment. Serum concentrations of soluble transferrin receptor (sTfR) were used as a parameter of erythropoietic activity and functional iron status. The levels of a recently discovered negative regulator of hepcidin expression, growth differentiation factor 15 (GDF15), a molecule expressed by erythroid precursor cells, were also measured.13 Finally, we determined the influence of the inherited red cell disorder Southeast Asian ovalocytosis on iron homeostasis in our study population. Southeast Asian ovalocytosis is highly prevalent on Sumba (estimated prevalence 10%)14 and may affect iron homeostasis, as reported for other red cell disorders.15
Design and Methods
Study site
This study was conducted in the Kodi region of West Sumba District, East Nusa Tenggara Province, Indonesia, an area with perennial P. falciparum and P. vivax transmission, stable enough to induce clinical immunity.16
Study design
This study used archived material collected during in vivo drug sensitivity tests in the Southwest Sumba District, East Nusa Tenggara Province, Indonesia. Overall, 1197 children aged 5 through 15 years from seven elementary schools were screened for the presence of parasitemia by microscopic examination of thick and thin malaria smears. All malaria smears were read the same day. The study team returned the following morning and those children with asymptomatic parasitemia were enrolled after written informed consent from parents or guardians and consent from the child. Asymptomatic parasitemia was defined as asexual P. falciparum or P. vivax parasitemia in the absence of fever (temperature ≤ 37.9°C) and of clinical signs or symptoms suggestive of malaria or another infectious disease. Seventeen healthy children with negative malaria smears were enrolled as a control group. A history was taken and a short physical examination performed according to a predefined checklist before 3 mL of venous blood were drawn into a serum and EDTA tube. Smear-positive children were treated with a 3-day regimen of artesunate-amodiaquine (P. falciparum) or a combination regimen of 3 and 7 days of chloroquine and primaquine (P. vivax), according to guidelines from the Indonesian Ministry of Health. Finally, seven subjects with clinical P. falciparum (n=4) or P. vivax (n=3) malaria who presented to the local hospital were enrolled as a positive control group.
To evaluate the effect of treatment of asymptomatic parasitemia, a follow-up visit was conducted 4 weeks after antimalarial treatment. The same study procedures were performed as during enrollment. The study was carried out with ethical approval from the Eijkman Institute for Molecular Biology Research Ethics Committees (Jakarta, Indonesia).
Laboratory methods
Thick and thin blood smears were stained with Giemsa and examined by a certified microscopist using 1000× oil immersion light microscopy. Parasite density was quantified against 200 white blood cells and converted to parasites per microliter of blood using the measured leukocyte count. The blood smear was also used to assess the presence of ovalocytes, which strongly suggests the presence of Southeast Asian ovalocytosis, a disease that results from a 27-bp deletion in the band 3 gene.17 Presence of ovalocytes in blood smears from Sumbanese was highly predictive for the presence of the band 3 gene mutation (Dr. Syafruddin, unpublished data). A full blood count was determined by a standard hematology analyzer (Arcus, Diatron, Vienna, Austria).
Serum hepcidin was measured using a combination of weak cation exchange chromatography and time-of-flight mass spectrometry (TOF MS). An internal standard (synthetic hepcidin-24; Peptide International Inc.) was used for quantification.18,19 Peptide spectra were generated on a Microflex LT matrix-enhanced laser desorption/ionization TOF MS platform (Bruker Daltonics). Serum hepcidin-25 concentrations were expressed in nanomoles. The lower limit of detection of this method was 0.5 nM; average coefficients of variation were 2.7% (intra-run) and 6.5% (inter-run). The median reference level of serum hepcidin-25 concentrations in healthy Dutch adults is 4.2 nM, while the range is 0.5–13.9 nM.20
Serum concentrations of total iron, total iron-binding capacity (TIBC), ferritin, sTfR and C-reactive protein were determined as described previously.11,12 GDF15 concentrations were measured with DuoSet (R&D Systems, Minneapolis, MN, USA) enzyme-linked immunosorbent assay for human GDF15 according to the manufacturer’s protocol. Serum concentrations of interleukin-1 receptor antagonist (IL-1ra), interleukin (IL)-6, IL-10 and tumor necrosis factor-α (TNF-α) were analyzed in one batch using a multiplex assay (Luminex, Austin, TX, USA) with lower limits of detection of 7 pg/mL (IL-1ra, IL-6 and IL-10) and 20 pg/mL (TNF-α).
Statistical analysis
Differences in characteristics and laboratory parameters between subjects in the malaria and control groups and among subjects before and after antimalarial treatment were analyzed by Student’s t test and paired Student’s t test, respectively. A two-sided P value of less than 0.05 was considered to be statistically significant. All analyses were performed with the use of SPSS software.
Results
Characteristics
Of a total of 1197 screened schoolchildren, 73 (6.1%) had asymptomatic P. falciparum and 18 (1.5%) asymptomatic P. vivax parasitemia. Seventeen asymptomatic children with a negative malaria smear were enrolled as a control group (see Table 1 for the baseline characteristics). Only 16% and 17% of children with asymptomatic P. falciparum and P. vivax parasitemia, respectively, had an elevated (>5 mg/L) C-reactive protein concentration; the mean C-reactive protein concentration in those children was 13.7 mg/L (range, 6–39 mg/L). Serum concentrations of IL-1ra, IL-6, IL-10 and TNF-α were low with values above the lower detection limit in only 65%, 15%, 38% and 4% of baseline samples, respectively. Hemoglobin concentrations were significantly lower in children with either P. falciparum or P. vivax parasitemia than in controls (Figure 1). Antimalarial treatment resulted in a modest, though significant increase in hemoglobin concentration in both groups with mean concentration increasing from the baseline 12.6 g/dL to 13.4 g/dL (P<0.01) after treatment in the group with asymptomatic P. falciparum and from 12.2 g/dL to 13.7 g/dL (P<0.01) in the group with asymptomatic P. vivax.
Table 1.
Baseline characteristics of the study participants.

Figure 1.
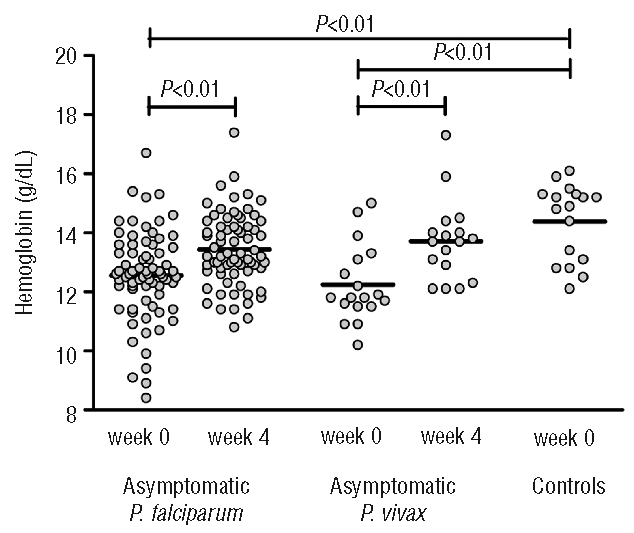
Hemoglobin concentrations in children with asymptomatic P. falciparum parasitemia (n=73), asymptomatic P. vivax parasitemia (n=18) and controls (n=17). Data depicted are before and 4 weeks after the start of antimalarial treatment; the line represents the mean. P values were determined using the Student’s t test and paired t test.
Serum hepcidin and parameters of iron status and inflammation
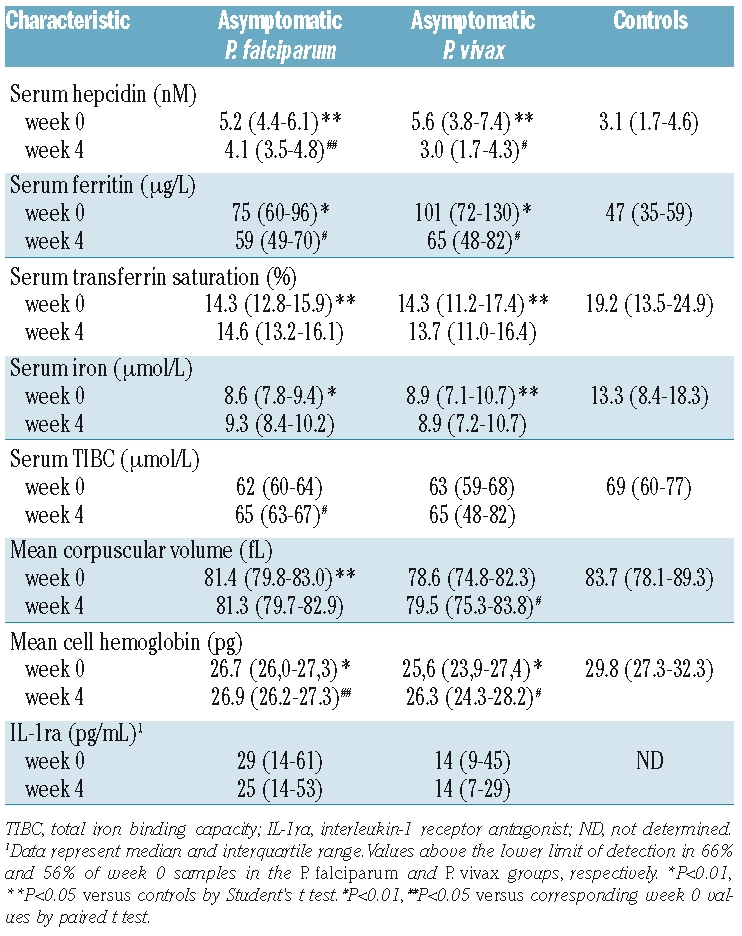
Baseline and week 4 serum concentrations of hepcidin and IL-1ra and values of the iron status parameters are presented in Figure 2 and Table 2. Children with asymptomatic P. falciparum or P. vivax parasitemia had significantly higher baseline hepcidin concentrations than those of the control children, together with higher serum ferritin concentrations and lower values of serum iron, transferrin saturation, TIBC, mean corpuscular volume and mean cell hemoglobin. Antimalarial treatment resulted in small but significant decreases in serum concentrations of hepcidin and ferritin in both groups with asymptomatic parasitemia. Hepcidin concentrations decreased by a median of 22% and 40% in the P. falciparum and P. vivax group, whereas ferritin concentrations decreased by a median of 14% and 30%, respectively. This decrease was associated with small increases in TIBC (P. falciparum group), mean corpuscular volume (P. vivax group) and mean cell hemoglobin (both groups), while the transferrin saturation and serum iron remained unchanged. There was no change in IL-1ra concentrations at week 4. However, C-reactive protein concentrations normalized to values below 5 mg/L in 12 of 15 children with elevated C-reactive protein concentrations before antimalarial treatment, while IL-10 concentrations became undetectable in 10 of 35 children with detectable IL-10 concentrations at baseline. The effect of antimalarial treatment on IL-6 and TNF-α concentrations could not be determined as serum concentrations of the baseline samples were below the lower detection limit in the vast majority of children.
Figure 2.
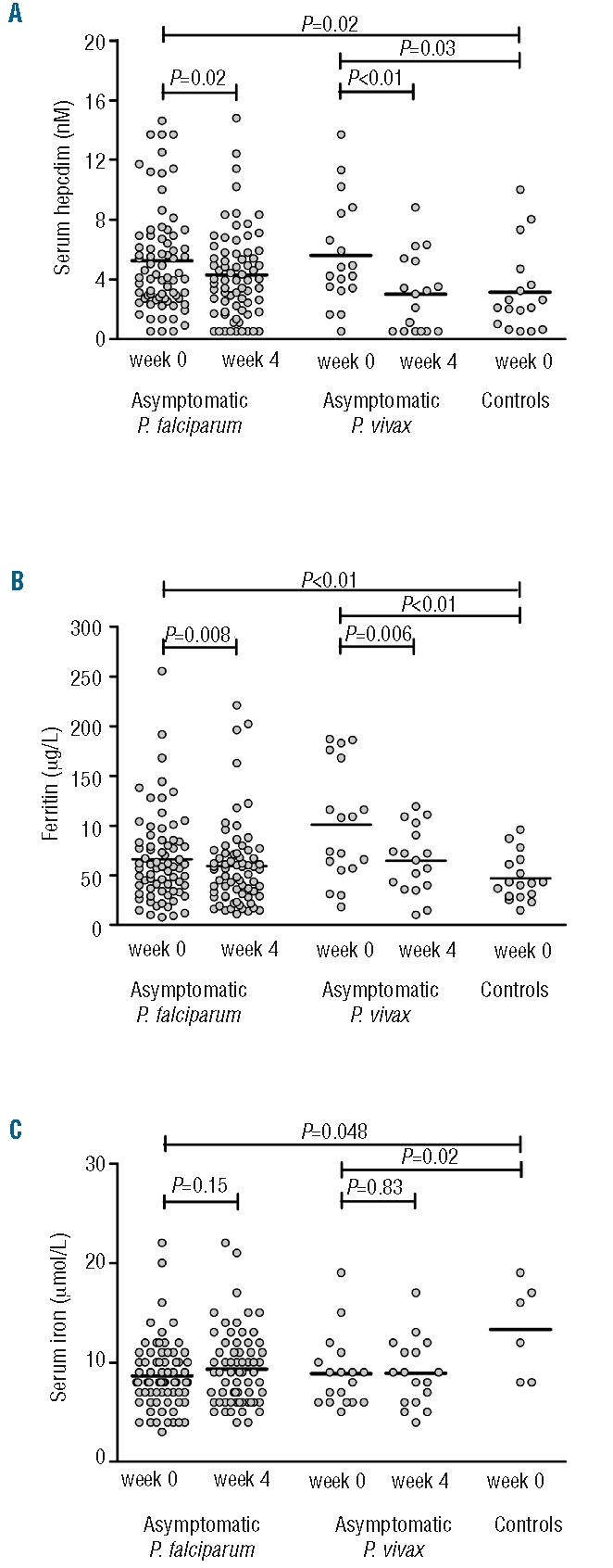
Serum concentrations of hepcidin (A), ferritin (B) and transferrin saturation (C) in children with asymptomatic P. falciparum parasitemia (n=73), asymptomatic P. vivax parasitemia (n=18) and controls (n=17). Data depicted are before and 4 weeks after the start of antimalarial treatment; the line represents the mean. P values were determined using the Student’s t test and paired t test.
Table 2.
Serum hepcidin and iron parameters (mean with 95% confidence interval) at baseline and 4 weeks after antimalarial treatment.
Soluble transferrin receptor and growth differentiation factor 15 concentrations
Erythropoietic activity was evaluated by assessment of serum concentrations of sTfR and GDF15. Both asymptomatic P. falciparum and P. vivax malaria were associated with markedly elevated sTfR concentrations, which remained elevated at week 4 (Figure 3A). In contrast, no differences in GDF15 concentrations were observed between the groups (Figure 3B). Erythropoietic activity is a negative regulator of hepcidin expression and serum concentrations of sTfR in the children with asymptomatic parasitemia were indeed inversely correlated with serum hepcidin concentrations (Pearson’s R=− 0.33; P=0.002).
Figure 3.
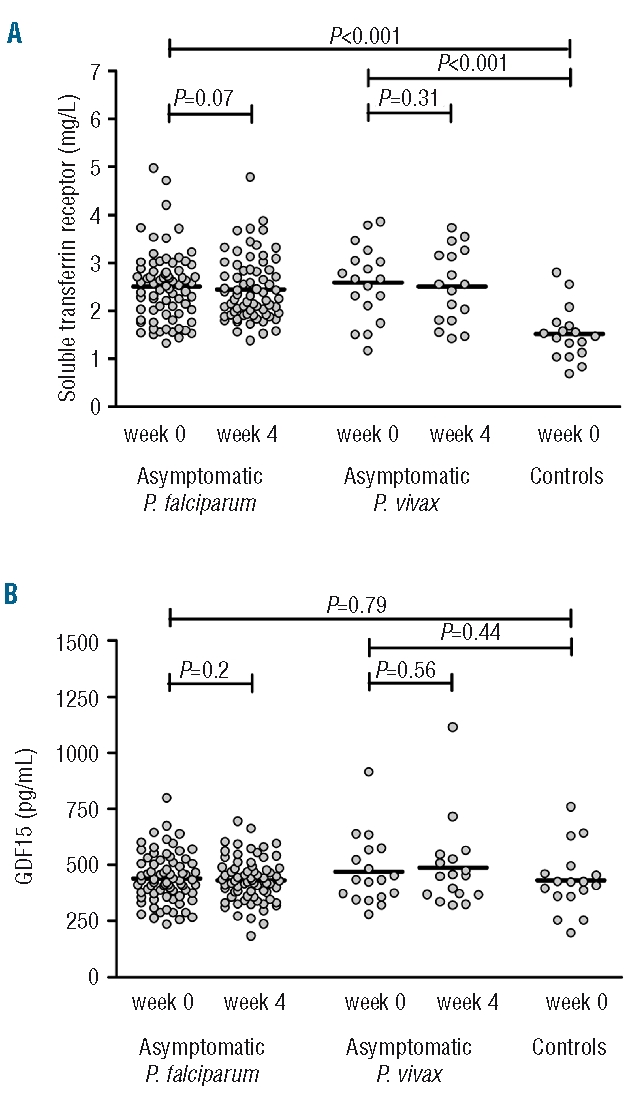
Serum concentrations of soluble transferrin receptor (A) and growth differentiation factor 15 (B) in children with asymptomatic P. falciparum parasitemia (n=73), asymptomatic P. vivax parasitemia (n=18) and controls (n=17). Data depicted are before and 4 weeks after the start of antimalarial treatment; the line represents the mean. P values determined using the Student’s t test and paired t test.
Since no data on GDF15 have previously been reported in malaria patients, we also determined concentrations of this protein in seven patients who presented with febrile, symptomatic P. falciparum (n=4) or P. vivax (n=3) malaria to the regional hospital. Symptomatic malaria was associated with lower sTfR concentrations (mean 1.3 mg/L; range, 1.0–2.2 mg/L) and higher GDF15 concentrations (mean 2467 pg/mL; range, 940–4500 pg/mL) when compared with asymptomatic parasitemia. Although GDF15 has been reported to down-regulate hepcidin expression, inflammation was dominant as serum hepcidin concentrations were markedly elevated (mean 24.3 nM; range 6.8–37.9 nM) in the presence of high serum concentrations of CRP (mean 83 mg/L; range 14–211 mg/L) and ferritin (mean 713 μg/L; range, 97–1387 μg/L) and low serum iron concentrations (mean 3.1 μmol/L; range, 2.0 – 6.0 μmol/L).
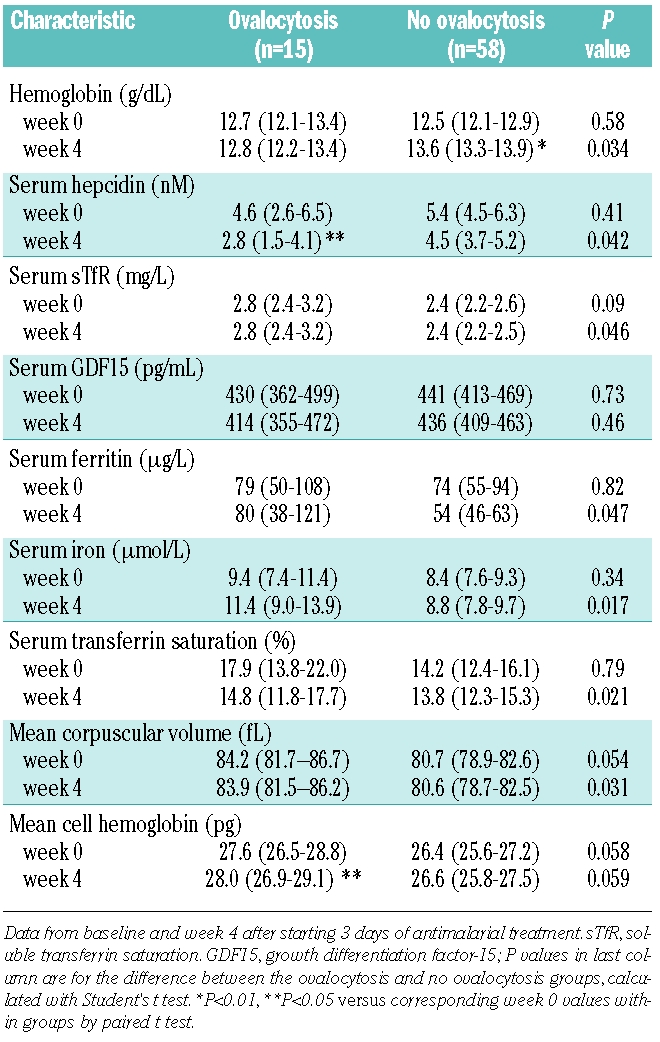
Ovalocytosis
Ovalocytes were seen in the blood smear in 15 of 73 (21%) children with asymptomatic P. falciparum parasitemia (Table 1), a finding suggestive of Southeast Asian ovalocytosis. There were no significant differences in demographic or clinical characteristics between children with and without ovalocytes (data not shown). No significant differences in iron parameters were observed between the two groups at baseline, except for higher value of mean corpuscular volume and mean cell hemoglobin (Table 3). By week 4, however, children with ovalocytosis had a slightly higher sTfR concentration, together with lower serum concentrations of hepcidin, serum iron, ferritin and transferrin saturation (Table 3). In addition, while antimalarial treatment induced a significant rise in hemoglobin concentration in the group without ovalocytosis, there was no change in hemoglobin or serum ferritin concentrations in the group with ovalocytosis.
Table 3.
Hematologic and iron parameters (mean with 95% confidence interval) in children with and without ovalocytosis.
Discussion
The present study shows that serum hepcidin concentrations are increased in children with asymptomatic P. falciparum and P. vivax parasitemia in the absence of a marked acute phase response. These higher hepcidin concentrations were accompanied by findings suggestive of iron maldistribution, i.e. higher serum ferritin concentrations and lower values of serum transferrin saturation, serum iron concentration and mean corpuscular volume. Antimalarial treatment partly reversed these abnormalities and resulted in a significant increase in hemoglobin concentration and significant decreases in serum concentrations of hepcidin and ferritin.
Are these mildly elevated hepcidin concentrations functionally important, i.e. do they reduce the availability of iron for erythropoiesis? We recently showed in an experimental human malaria model that even a mild increase in serum hepcidin, to concentrations in the range of those observed in the current study, induces hypoferremia and a steep fall in the hemoglobin content of reticulocytes, which is suggestive of impaired iron incorporation in hemoglobin.12 Moreover, the lower values of hemoglobin, mean corpuscular volume, serum iron concentration and transferrin saturation in children with asymptomatic parasitemia also support the presence of functional iron deficiency. Since asymptomatic P. falciparum and P. vivax parasitemia are chronic and relapsing conditions, a possible delocalization of iron with iron retention in monocytes/macrophages may theoretically increase the risk of anemia and of infections with intracellular bacteria (e.g. Salmonella and Mycobacteria spp.), which are extremely common in regions in which malaria is endemic. We, therefore, suggest screening individual children in malaria areas for the presence of malaria when iron therapy is considered and including malaria screening and treatment as an integral part of universal iron supplementation programs. In addition, as recently proposed by Theurl et al. and by our group, hepcidin is a promising new parameter for the selection of individuals who will benefit from iron therapy.11,12,21
The exact cause of the up-regulated hepcidin production in asymptomatic malaria remains speculative. Previous studies found only little inflammation in patients with asymptomatic parasitemia, which is in line with the low circulating concentrations of C-reactive protein and inflammatory cytokines in our present study.22 Therefore, even though an effect of mild inflammation cannot be fully excluded, other mechanisms should be considered. Recent work by Armitage et al. showed that P. falciparum-parasitized erythrocytes directly up-regulate hepcidin production by peripheral blood mononuclear cells via pattern recognition receptors.23 Our earlier observations, in children with febrile malaria, that IL-6 concentrations were independent of urine hepcidin concentrations in multivariate regression analysis are also consistent with the existence of such an IL-6-independent pathway in malaria.11
Erythropoietic activity is also increasingly recognized as a central regulator of hepcidin production.24 Although the precise pathways are yet to be elucidated, two novel erythroid regulators, TWSG1 (twisted gastrulation homolog 1) and GDF15, were recently shown to down-regulate hepcidin expression.13,25 GDF15 is a molecule expressed by erythroid precursors and increased GDF15 concentrations have been found in patients with thalassemia major and other conditions with ineffective erythropoiesis.13,26,27 Although earlier bone marrow studies in subjects with low density P. falciparum and with P. vivax malaria were suggestive of ineffective erythropoiesis, the normal GDF15 concentrations in our present study do not support this.28–30 Instead, as also previously reported by Verhoef et al.,31 the elevated sTfR concentrations, a parameter thought to correlate well with erythroid mass,32 suggests that erythropoiesis is effectively increased in asymptomatic parasitemia. Increased erythropoietic activity compensating for increased red cell destruction may negate some of the hepcidin-stimulatory effects of malaria antigen or inflammation. In contrast, erythropoiesis is usually suppressed in symptomatic malaria, as reflected by the low sTfR concentrations together with elevated GDF15 concentrations in the seven patients with febrile malaria in our study.33
Southeast Asian ovalocytosis is a common inheritable red blood cell disorder on the island of Sumba. It results from a 27-base pair deletion in the band 3 gene, which leads to rigid and oval-shaped red blood cells. In our study, the presence of ovalocytes in the blood smear, a finding highly suggestive of Southeast Asian ovalocytosis,17 was related to higher sTfR concentrations and normal GDF15 concentrations. A reduced life span of ovalocytes with a compensatory increase in erythropoiesis may account for this finding and may also explain why antimalarial treatment resulted in a greater decrease in serum hepcidin concentrations. By contrast, no changes in hemoglobin and ferritin concentrations were observed in the ovalocytosis group, a finding not readily explained though possibly related to ongoing low-grade hemolysis and erythrophagocytosis of the rigid ovalocytes. Finally, thalassemia may also influence iron homeostasis, but this hemoglobinopathy seemed rare in our study population because less than 7% of children had microcytosis (<70 fL), a finding that is considered typical of thalassemia. Moreover, none of these children with microcytosis also had strongly elevated concentrations of sTfR and GDF15, which are usually present in thalassemia intermedia or major.13
In conclusion, asymptomatic P. falciparum and P. vivax parasitemia were associated with lowered hemoglobin concentrations and increased serum hepcidin concentrations in the absence of marked inflammation. Antimalarial treatment resulted in a decrease in serum hepcidin concentrations and an increase in hemoglobin concentrations, although a causal relationship between these findings is unproven. We speculate that asymptomatic parasitemia with impaired iron handling may be an underestimated cause of anemia in tropical countries. We suggest that universal iron supplementation programs should not be implemented without proper screening for parasitemia and functional iron deficiency. Hepcidin may prove a useful screening tool for this purpose.
Acknowledgments
the authors would like to thank E. Wiegerinck, S. Klaver, S. Verweij and C. Jacobs for performing the measurements of hepcidin and markers of iron status and inflammation.
Footnotes
Funding: this work was supported by a junior research grant from the Radboud University Nijmegen Medical Centre (QdM) and NWO-WOTRO (WIZ93-465) through PRIOR (PBA).
Authorship and Disclosures
QdM and AJV designed the study, drafted the manuscript and analyzed the data. DS, PBA, SK, TOR, and OD recruited the study participants and collected the blood samples. DS supervised the laboratory work. All authors contributed to the interpretation and discussion of the results and approved the final manuscript.
The authors reported no potential conflicts of interest.
References
- 1.Andrews NC. Forging a field: the golden age of iron biology. Blood. 2008;112(2):219–30. doi: 10.1182/blood-2007-12-077388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Weiss G. Iron metabolism in the anemia of chronic disease. Biochim Biophys Acta. 2009;1790(7):682–93. doi: 10.1016/j.bbagen.2008.08.006. [DOI] [PubMed] [Google Scholar]
- 3.Conclusions and recommendations of the WHO Consultation on prevention and control of iron deficiency in infants and young children in malaria-endemic areas. Food Nutr Bull. 2007;28:S621–7. doi: 10.1177/15648265070284s414. [DOI] [PubMed] [Google Scholar]
- 4.UNICEF/UNU/WHO/MI. Preventing Iron Deficiency in Women and Children: Technical Consensus on Key Issues. Technical Workshop; October 7–9, 1998; Boston and Ottawa: International Nutrition Foundation and MI; [Google Scholar]
- 5.Prentice AM, Ghattas H, Doherty C, Cox SE. Iron metabolism and malaria. Food Nutr Bull. 2007;28(4 Suppl):S524–39. doi: 10.1177/15648265070284S406. [DOI] [PubMed] [Google Scholar]
- 6.Sazawal S, Black RE, Ramsan M, Chwaya HM, Stoltzfus RJ, Dutta A, et al. Effects of routine prophylactic supplementation with iron and folic acid on admission to hospital and mortality in preschool children in a high malaria transmission setting: community-based, randomised, placebo-controlled trial. Lancet. 2006;367 (9505):133–43. doi: 10.1016/S0140-6736(06)67962-2. [DOI] [PubMed] [Google Scholar]
- 7.Bottius E, Guanzirolli A, Trape JF, Rogier C, Konate L, Druilhe P. Malaria: even more chronic in nature than previously thought; evidence for subpatent parasitaemia detectable by the polymerase chain reaction. Trans R Soc Trop Med Hyg. 1996;90(1):15–9. doi: 10.1016/s0035-9203(96)90463-0. [DOI] [PubMed] [Google Scholar]
- 8.Camargo EP, Alves F, Pereira da Silva LH. Symptomless Plasmodium vivax infections in native Amazonians. Lancet. 1999;353 (9162):1415–6. doi: 10.1016/s0140-6736(99)00941-1. [DOI] [PubMed] [Google Scholar]
- 9.Genton B, D’Acremont V, Rare L, Baea K, Reeder JC, Alpers MP, et al. Plasmodium vivax and mixed infections are associated with severe malaria in children: a prospective cohort study from Papua New Guinea. PLoS Med. 2008;5(6):e127. doi: 10.1371/journal.pmed.0050127. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Tjitra E, Anstey NM, Sugiarto P, Warikar N, Kenangalem E, Karyana, et al. Multidrug-resistant Plasmodium vivax associated with severe and fatal malaria: a prospective study in Papua, Indonesia. PLoS Med. 2008;5(6):e128. doi: 10.1371/journal.pmed.0050128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.de Mast Q, Nadjm B, Reyburn H, Kemna EH, Amos B, Laarakkers C, et al. Assessment of urinary concentrations of hepcidin provides novel insight into disturbances in iron homeostasis during malarial infection. J Infect Dis. 2009;199(2):253–62. doi: 10.1086/595790. [DOI] [PubMed] [Google Scholar]
- 12.de Mast Q, van Dongen-Lases EC, Swinkels DW, Nieman AE, Roestenberg M, Druilhe P, et al. Mild increases in serum hepcidin and interleukin-6 concentrations impair iron incorporation in haemoglobin during an experimental human malaria infection. Br J Haematol. 2009;145(6):657–64. doi: 10.1111/j.1365-2141.2009.07664.x. [DOI] [PubMed] [Google Scholar]
- 13.Tanno T, Bhanu NV, Oneal PA, Goh SH, Staker P, Lee YT, et al. High levels of GDF15 in thalassemia suppress expression of the iron regulatory protein hepcidin. Nat Med. 2007;13(9):1096–101. doi: 10.1038/nm1629. [DOI] [PubMed] [Google Scholar]
- 14.Kimura M, Soemantri A, Ishida T. Malaria species and Southeast Asian ovalocytosis defined by a 27-bp deletion in the erythrocyte band 3 gene. Southeast Asian J Trop Med Public Health. 2002;33(1):4–6. [PubMed] [Google Scholar]
- 15.Kearney SL, Nemeth E, Neufeld EJ, Thapa D, Ganz T, Weinstein DA, et al. Urinary hepcidin in congenital chronic anemias. Pediatr Blood Cancer. 2007;48(1):57–63. doi: 10.1002/pbc.20616. [DOI] [PubMed] [Google Scholar]
- 16.Syafruddin D, Krisin K, Asih P, Sekartuti S, Dewi RM, Coutrier F, et al. Seasonal prevalence of malaria in West Sumba district, Indonesia. Malar J. 2009;8:8. doi: 10.1186/1475-2875-8-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.O’Donnell A, Allen SJ, Mgone CS, Martinson JJ, Clegg JB, Weatherall DJ. Red cell morphology and malaria anaemia in children with Southeast-Asian ovalocytosis band 3 in Papua New Guinea. Br J Haematol. 1998;101(3):407–12. doi: 10.1046/j.1365-2141.1998.00742.x. [DOI] [PubMed] [Google Scholar]
- 18. [Accessed on May 14th 2009]. www.hepcidinanalysis.com.
- 19.Swinkels DW, Girelli D, Laarakkers C, Kroot J, Campostrini N, Kemna EH, et al. Advances in quantitative hepcidin measurements by time-of-flight mass spectrometry. PLoS ONE. 2008;3(7):e2706. doi: 10.1371/journal.pone.0002706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Kroot JJ, Hendriks JC, Laarakkers CM, Klaver SM, Kemna EH, Tjalsma H, et al. (Pre)analytical imprecision, between-subject variability, and daily variations in serum and urine hepcidin: implications for clinical studies. Anal Biochem. 2009;389(2):124–9. doi: 10.1016/j.ab.2009.03.039. [DOI] [PubMed] [Google Scholar]
- 21.Theurl I, Aigner E, Theurl M, Nairz M, Seifert M, Schroll A, et al. Regulation of iron homeostasis in anemia of chronic disease and iron deficiency anemia: diagnostic and therapeutic implications. Blood. 2009;113(21):5277–86. doi: 10.1182/blood-2008-12-195651. [DOI] [PubMed] [Google Scholar]
- 22.Imrie H, Fowkes FJ, Michon P, Tavul L, Reeder JC, Day KP. Low prevalence of an acute phase response in asymptomatic children from a malaria-endemic area of Papua New Guinea. Am J Trop Med Hyg. 2007;76(2):280–4. [PubMed] [Google Scholar]
- 23.Armitage AE, Pinches R, Eddowes LA, Newbold CI, Drakesmith H. Plasmodium falciparum infected erythrocytes induce hepcidin (HAMP) mRNA synthesis by peripheral blood mononuclear cells. Br J Haematol. 2009;147(5):769–71. doi: 10.1111/j.1365-2141.2009.07880.x. [DOI] [PubMed] [Google Scholar]
- 24.Nemeth E. Iron regulation and erythropoiesis. Curr Opin Hematol. 2008;15(3):169–75. doi: 10.1097/MOH.0b013e3282f73335. [DOI] [PubMed] [Google Scholar]
- 25.Tanno T, Porayette P, Sripichai O, Noh SJ, Byrnes C, Bhupatiraju A, et al. Identification of TWSG1 as a second novel erythroid regulator of hepcidin expression in murine and human cells. Blood. 2009;114(1):181–6. doi: 10.1182/blood-2008-12-195503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ramirez JM, Schaad O, Durual S, Cossali D, Docquier M, Beris P, et al. Growth differentiation factor 15 production is necessary for normal erythroid differentiation and is increased in refractory anaemia with ring-sideroblasts. Br J Haematol. 2009;144(2):251–62. doi: 10.1111/j.1365-2141.2008.07441.x. [DOI] [PubMed] [Google Scholar]
- 27.Tamary H, Shalev H, Perez-Avraham G, Zoldan M, Levi I, Swinkels DW, et al. Elevated growth differentiation factor 15 expression in patients with congenital dyserythropoietic anemia type I. Blood. 2008;112(13):5241–4. doi: 10.1182/blood-2008-06-165738. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Abdalla SH. Hematopoiesis in human malaria. Blood Cells. 1990;16(2–3):401–16. [PubMed] [Google Scholar]
- 29.Wickramasinghe SN, Looareesuwan S, Nagachinta B, White NJ. Dyserythropoiesis and ineffective erythropoiesis in Plasmodium vivax malaria. Br J Haematol. 1989;72(1):91–9. doi: 10.1111/j.1365-2141.1989.tb07658.x. [DOI] [PubMed] [Google Scholar]
- 30.Wickramasinghe SN, Abdalla SH. Blood and bone marrow changes in malaria. Baillieres Best Pract Res Clin Haematol. 2000;13(2):277–99. doi: 10.1053/beha.1999.0072. [DOI] [PubMed] [Google Scholar]
- 31.Verhoef H, West CE, Ndeto P, Burema J, Beguin Y, Kok FJ. Serum transferrin receptor concentration indicates increased erythropoiesis in Kenyan children with asymptomatic malaria. Am J Clin Nutr. 2001;74(6):767–75. doi: 10.1093/ajcn/74.6.767. [DOI] [PubMed] [Google Scholar]
- 32.Beguin Y. Soluble transferrin receptor for the evaluation of erythropoiesis and iron status. Clin Chim Acta. 2003;329(1):9–22. doi: 10.1016/s0009-8981(03)00005-6. [DOI] [PubMed] [Google Scholar]
- 33.Lamikanra AA, Brown D, Potocnik A, Casals-Pascual C, Langhorne J, Roberts DJ. Malarial anemia: of mice and men. Blood. 2007;110(1):18–28. doi: 10.1182/blood-2006-09-018069. [DOI] [PubMed] [Google Scholar]