
Figure 4. Model for the evolutionary origin of the insect DV patterning strategy.
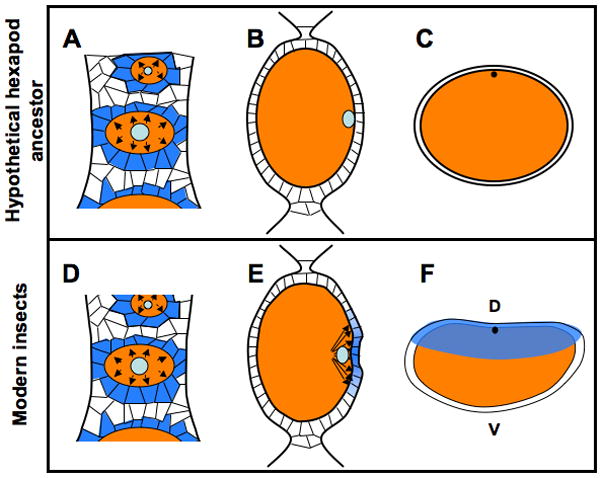
A–C: Schematic representation of features present in a hypothetical insect ancestor that predated the use of germline-soma signaling for DV axis establishment. Since the ovariole structure of ovaries is synapomorphic for insects, a mechanism for germline-soma communication in mediating the encapsulation of oocytes (orange ovals) by somatic cells was present in a common insect ancestor. Those somatic cells (marked in blue in A) receiving the EGF signal emanating from the oocyte (represented by outward pointing arrows) will give rise to the follicle cells, which later give rise to eggshell structures such as the vitelline membrane and chorion, but at this stage have no role in receiving or transmitting patterning information. In this hypothetical ancestor, the oocyte nucleus (light blue) moved to a cortical location (B), to allow the formation of the polar body (black spot) in the egg after laying, but originally had no role in embryonic patterning (C).
D–F: Features present in modern insects, based on results presented in this manuscript. The role in EGF signaling in recruiting follicle cells to the oocyte (A) is maintained (D). The oocyte nucleus still moves to the cortex (E), as this event is still critical for regulation of meiosis. In this ancestor, however, EGF signaling becomes strongly associated with the position of the oocyte nucleus, allowing asymmetric activation of this pathway (blue gradient in E) in the overlying follicle cells. This patterning information is stored in the follicle cells, where it is incorporated into the eggshell (transparent blue in F) and is then later transmitted to the embryo to provide a basis for patterning the now maternally established DV axis.