

Summary
Background
It is widely believed that the hippocampus plays a temporary role in the retrieval of episodic and contextual memories. Initial research indicated that damage to this structure produced amnesia for newly acquired memories but did not affect those formed in the distant past. A number of recent studies, however, have found that the hippocampus is required for the retrieval of episodic and contextual memories regardless of their age. These findings are currently the subject of intense debate and a satisfying resolution has yet to be identified.
Results
The current experiments address this issue by demonstrating that detailed memories always require the hippocampus while memories that lose precision become independent of this structure. First, we show that the dorsal hippocampus is preferentially activated by the retrieval of detailed context fear memories. We then establish a causal relationship between the hippocampus and detailed memories using a context generalization procedure. Mice that exhibit high levels of generalization to a novel environment show no memory loss when the hippocampus is subsequently inactivated. In contrast, mice that discriminate between contexts are significantly impaired by hippocampus inactivation.
Conclusions
Our data suggest that detailed context memories always require the hippocampus (independent of their age) while memories that lose precision can be retrieved without this structure. These findings account for inconsistencies in the literature - memories of our distant past can be either lost or retained after hippocampus damage depending on their quality – and provide a new framework for understanding memory consolidation.
Highlights
The retrieval of new context fear memories increases immediate early gene expression in the hippocampus.
Immediate early gene expression decreases as memories age and become less precise.
If memory is tested at a single retention interval, hippocampus inactivation selectively impairs the retrieval of precise context memories.
Introduction
The hippocampus is essential for the formation and retrieval of episodic and contextual memory in humans and animals. It is well established that cell loss or dysfunction in this area produces significant amnesia for newly acquired information 1-5. In contrast, the fate of old memories is less clear. Initial work indicated that the hippocampus was not involved in the retrieval of episodic or contextual memories formed in the distant past. These findings gave rise to the idea that the hippocampal system plays a temporary role in the formation and retrieval of new memories as they are being permanently stored in regions of the neocortex (i.e. consolidation theory) 6-8. Recent work has presented several challenges to this theory. First, functional imaging studies (fMRI) commonly observe hippocampus activation during the retrieval of both new and old episodic memories 9-11. Consistent with these findings, damage to the hippocampal system in humans and animals often impairs the retrieval of both recent and remote memories 2, 12-18. Results like these have contributed to the development of multiple trace theory (MTT), which argues that episodic and contextual memories are permanently stored in the hippocampus 17. These findings are currently the subject of intense debate in the field and a satisfying resolution has yet to be identified.
The current paper sought to address these discrepancies by examining the relationship between memory quality and hippocampus-dependency. In previous experiments, we demonstrated that context memories become less specific and more general during the consolidation period 19. Mice trained to fear a specific context are initially able to discriminate between it and a similar environment. However, as the time between training and testing is increased, memory becomes less accurate and animals are unable to distinguish between these contexts. Based on these results we suggested that memory quality may be a critical factor that determines whether or not the hippocampus is essential for retrieval. As memories become less detailed and more general across time they gradually become independent of this structure. However, when the details of memories are retained then the hippocampus continues to be necessary for their retrieval. This idea is consistent with previous theories that argue the function of consolidation is to gradually integrate memories of specific events with general knowledge that is stored in the neocortex 2, 20, 21.
Results
The retrieval of recent but not remote memory increases immediate early gene expression in the hippocampus
To test the idea just described we used contextual fear conditioning in mice and two different complementary approaches. Our first approach was to monitor hippocampus activity during memory retrieval using immediate early gene (IEG) expression as measured by qRT-PCR. This technique has several advantages over immunohistochemistry and in situ hybridization (ISH), which are commonly used to index IEG activity. First, very small changes in mRNA expression can be amplified and detected via PCR. Second, the observed changes can be quickly and reliably quantified without the use of stereology or image thresholding. Therefore, we first determined if qRT-PCR was capable of detecting increased gene expression in the dorsal hippocampus following contextual fear conditioning. To optimize IEG expression mice were trained with a robust context conditioning protocol (five, 0.75 mA shocks). Arc, C-fos and Zif268 expression were analyzed in mice sacrificed immediately, 15 minutes, 30 minutes or 60 minutes after training (Figure 1A). We found an increase in the expression of all three IEGs (significant effect of time, F (4, 36) = 3.71, p < .05, no effect of gene F (2, 9) =3.27, p > .05, no time × gene interaction, F < 1) that was significantly different from homecage controls (HC) at 15 minutes (Fisher's PLSD, p < .05) (Figure 1B). We next determined if qRT-PCR was sensitive enough to detect activity changes that occur in the hippocampus during the consolidation period. To do this, mice were trained with 5 shocks and tested 1 or 28 days later (Figure 1C). Consistent with previous results, we found reduced expression of Arc, C-fos and Zif268 in the dorsal hippocampus following the retrieval of remote memories (significant effect of test day, F (1,6) = 25.35, p < .05), no effect of gene, F < 1, no test day × gene interaction, F < 1) 22-24 (Figure 1E). This effect was not due to differences in performance as freezing was equivalent during the recent and remote memory tests (no effect of test day, F (1,6) = 1.37, p > .05) (Figure 1D).
Figure 1. Immediate early gene expression is increased in the dorsal hippocampus following the retrieval of recent context fear memories.
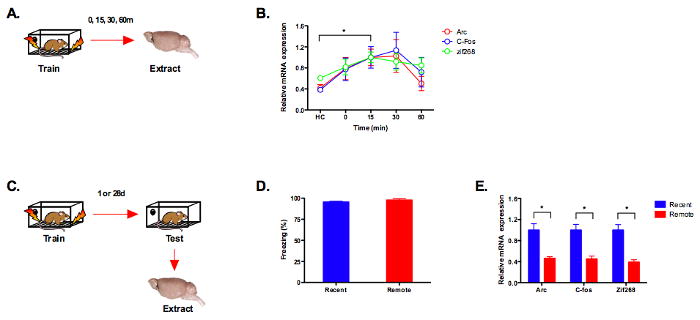
(a) Experimental design. (b) Immediate early gene expression (Arc, C-fos, Zif268) (n = 4 per group) was significantly increased 15 minutes after training relative to homecage controls (p < .05). (c) Experimental design. (d) Freezing levels during 1d (recent, n =4) and 28d (remote, n = 4) context fear tests were equivalent (p > .05). (e) Immediate early gene expression (Arc, c-fos, zif268) was reduced in the dorsal hippocampus during the retrieval of remote memory (p < .05). Note: mRNA expression is shown relative to the recent memory test.
Temporary inactivation of the hippocampus impairs the retrieval of recent but not remote memory
In the next experiment, we demonstrated that hippocampus inactivation selectively impairs the retrieval of recently formed context fear memories. Mice were trained with a single shock and tested 1 or 28 days later (Figure 2). The hippocampus was then transiently inactivated with the AMPA receptor antagonist CNQX. Temporary inactivation provides several advantages over traditional lesion methodology including an absence of damage to distal structures such as the amygdala and neocortex25. Similar to previous studies, inactivation of the dorsal hippocampus with CNQX impaired the retrieval of recent but not remote memories (significant test day × infusion interaction (F (1, 68) = 5.95, p < .05) (Figure 2B) 3, 8. After testing, we extracted the dorsal hippocampus from saline infused animals and analyzed expression of Arc using qRT-PCR. Consistent with our previous experiment, Arc expression was elevated following the 1 day test relative to HC controls (p < .05) and decreased after the 28 day test (p < .05) (Figure 2C).
Figure 2. The retrieval of remote context fear memories does not require the dorsal hippocampus.
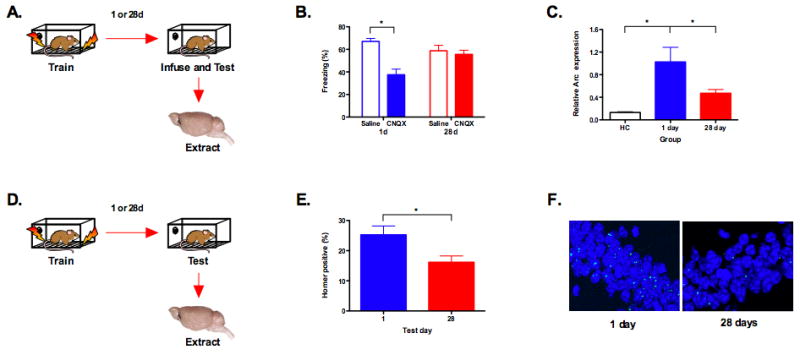
(a) Experimental design. (b) Inactivation of the dorsal hippocampus with CNQX impaired memory retrieval at day 1 (saline, n = 16, CNQX, n = 16) but not day 28 (saline, n = 20, CNQX, n = 20) (p < .05). (c) Arc expression in the saline animals increased during the day 1 test compared to homecage controls (HC) (n =3) (p < .05) and decreased during the day 28 test (p < .05). Note: mRNA expression is shown relative to the 1 day test. (d) Experimental design. (e) Fewer cells expressed Homer 1A in the CA1 region of the hippocampus during the day 28 test (n = 4) compared to the day 1 test (n = 4) (p < .05). (f) Representative samples of Homer1A expression following retrieval at 1 day and 28 days.
Since the mice froze slightly less during the 28 day memory test we conducted a regression analysis to determine if there was a relationship between freezing and Arc expression. We found no relationship between these variables at either 1 day (slope not significantly different from zero, F < 1, r2 = .002) or 28 days (slope not significantly different from zero, F < 1, r2 = .00012). This suggests that reduced Arc expression at 28 days is not due to reduced memory strength.
To determine if reduced IEG expression reflects less gene expression per cell or the activation of fewer neurons during memory retrieval we performed fluorescent in situ hybridization (FISH) for Homer 1A. Homer is an IEG that reliably indexes cellular activity in the hippocampus and neocortex. The correspondence between Arc and Homer expression in activated CA1 neurons is 95% 26, 27. We examined expression of Homer 1A in the CA1 region of the dorsal hippocampus during memory retrieval 1 or 28 days after training (Figure 2D). During these tests, we found a significant reduction in the percentage of neurons that expressed Homer at 28 days compared to 1 day (significant effect of test day, F (1, 6) = 6.41, p < .05) (Figure 2E and 2F). This suggests that the reduction in IEG expression during remote memory retrieval reflects the activation of fewer cells in the hippocampus.
Remote memories are less precise than recently formed memories
Next, we examined whether hippocampus activation was correlated with memory quality (Figure 3A). To do this we trained four separate groups of mice in context A and then tested them in a similar, yet novel environment (context B) at 1, 7, 14 or 28 days. We previously showed that mice exhibit increased generalization to this environment over time 19. Based on this finding we, and others, have argued that context memories lose many of their details with the passage of time 28-30. As a control, we tested an additional four groups of mice in a context that was entirely distinct from the training environment (context C). Mice showed significantly more generalization to the similar context than the distinct environment (significant effect of context, F (1, 49) = 169.2, p < .05). In addition, generalization was significantly higher at 28 days compared to 1 day in the similar context (significant effect of test day, F (1, 12) = 4.81, p < .05) but not in the distinct environment (F < 1) (Figure 3B). The fact that generalization to context B increased over time suggests that fear memories become less precise after training. These changes are not likely due to non-specific increases in fear (i.e. sensitization) as they were not observed in context C. Moreover, fear to context A did not increase over time in either the current experiments (Figures 1D and 2B) or in our previous study, suggesting that generalization to context B is not the result of increased memory strength (i.e. incubation) 19. Therefore, increases in generalization result from an inability to discriminate between the training context and a similar environment.
Figure 3. The dorsal hippocampus is less activated during the retrieval of generalized context fear memories.
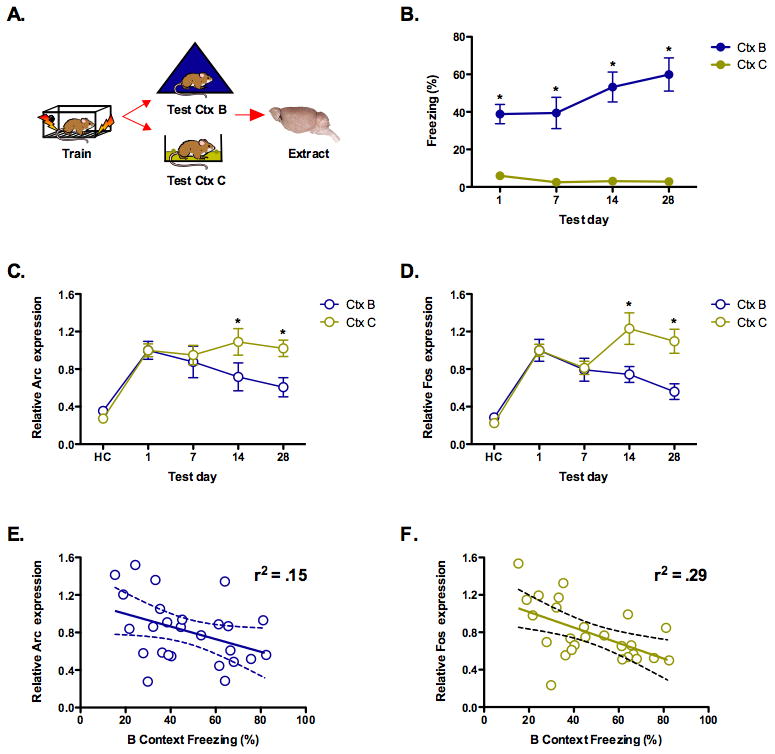
(a) Experimental design. (b) Mice showed more generalized fear to a context that was similar to the training environment (context B) than a distinct context (context C) (p < .05). Fear generalization to the similar environment increased over time (days 1, 7, 14, and 28, n = 8, 6, 6, 6) (p < .05) whereas fear of the distinct context did not (n = 8, 8, 8, 7) (p > .05). (c) Arc activation in the dorsal hippocampus decreased as generalization to context B increased (p < .05). Activation in context C did not change over time (p > .05). (d) C-fos activation in the dorsal hippocampus decreased as generalization to context B increased (p < .05). Activation in context C did not change (p > .05). (e) Linear regression analysis found a significant relationship between freezing in context B and Arc expression in dorsal hippocampus (p < .05). (f) Linear regression analysis found a significant relationship between freezing in context B and dorsal hippocampal c-Fos expression (p < .05). Note: mRNA expression is shown relative to the 1-day test.
We then examined IEG expression in the same animals and found that generalization was correlated with reduced activation of the dorsal hippocampus. As generalization to context B increased, the hippocampus expressed less Arc and C-fos while it remained activated by context C at all test intervals (significant context × test day interaction (F (3, 49) = 2.86, p < .05) (Figure 3C and 3D). Post hoc tests (Fisher's PLSD) revealed that IEG expression was significantly reduced in context B compared to context C at 14 and 28 days (p < .05). These data suggest that the hippocampus is less activated by the retrieval of generalized context fear memories. Previous work has shown that the excitability of hippocampal neurons is increased for several days after new learning 31, 32. Consequently, an alternative interpretation of the current data is that decreased IEG expression simply reflects a reduction in hippocampal activity after conditioning. This does not appear to be the case, however, as exposure to context C continued to produce robust IEG expression at all test intervals (likely driven by new learning). Therefore, the reduced hippocampal activation that occurs over time is contingent on memory retrieval.
A regression analysis conducted on the freezing scores in context B and IEG expression also revealed that there was a significant relationship between these factors for both Arc (slope significantly different from zero, F (1, 24) = 4.32, p < .05, r2 = .15) and C-Fos (slope significantly different from zero, F (1, 24) = 9.95, p < .05, r2 = .29) (Figures 3E and 3F). The more that mice generalized to context B the less IEG expression was observed in the dorsal hippocampus.
Memory precision can be predicted using context generalization
The previous experiments demonstrate that the hippocampus is less activated by remote context fear memories that have lost details. Consequently, reduced activity could be due to the age of a memory or its content. To test between these possibilities we first examined context fear generalization in more depth (Figure 4). We trained a large group of mice (n = 185) and tested them in context A and context B 1 or 14 days after training. Consistent with our previous results, context discrimination was significantly better at 1 day than 14 days (significant test day × context interaction, F (1, 183) = 28.878, p < .05) (Figure 4A)19. We then calculated a discrimination index (A/A+B) for each animal and constructed a frequency distribution (Figure 4B). As expected, the mean discrimination ratio was significantly higher at day 1 (x̄ = 0.67) than day 14 (x̄ = 0.58) (F (1, 183) = 20.964, p < .05). However, even though the majority of mice could discriminate at 1 day, a few animals were unable to do so (discrimination ratio < .5). In addition, a large number of mice tested at day 14 froze more in context A than context B (discrimination ratio > .5). If the hippocampus is necessary for the retrieval of detailed memories then inactivating it should selectively impair freezing in animals that discriminate, independent of the test interval. In the next section we developed a method to test this prediction.
Figure 4. Fear generalization is an excellent predictor of context discrimination.
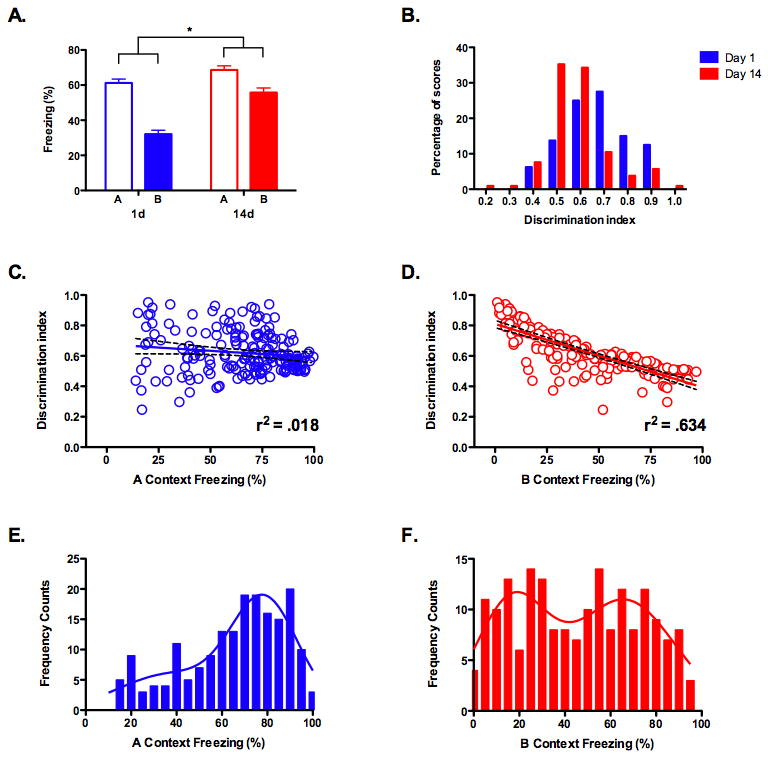
(a) A large group of mice were tested in context A and B, 1 day (n = 80) or 14 days (n = 105) after training. Discrimination was significantly better at day 1 compared to day 14 (p < .05). (b) Frequency distributions of the discrimination index (A/A + B) for mice tested at day 1 or day 14. The mean discrimination index was significantly lower at day 1 compared to day 14 (p < .05). (c) Linear regression analysis found no relationship between freezing in context A and the discrimination index (p > .05). (d) Linear regression analysis found a significant relationship between freezing in context B and the discrimination index (p < .05). (e) Non-linear regression analysis of freezing scores in context A found a single distribution with a mean of 77.72. (f) Non-linear regression analysis of freezing scores in context B found a bimodal distribution with means of 16.7 and 66.1, respectively. The intersection of these two distributions occurred at a freezing value of 42.
A regression analysis conducted on the freezing scores in context A and the discrimination index revealed that there was no relationship between these factors (slope not significantly different from zero, F (1, 183) = 3.47, p < .05, r2 = .018) (Figure 4C). In contrast, there was a robust relationship between freezing in context B and the ability to discriminate (slope significantly different from zero, F (1, 183) = 318, p < .05, r2 = .634) (Figure 4C). The less that mice generalized to context B the better they were able to discriminate. This was not simply a performance artifact (e.g. more/less freezing makes it harder/easier to discriminate) because the same relationship was not observed in context A. This suggests that discrimination is largely driven by the amount of freezing in context B. Therefore, using freezing scores in context B, we should be able to predict which mice are able to discriminate between contexts. If our hypothesis is correct, then hippocampus inactivation should selectively impair memory retrieval in these animals.
To categorize animals prior to hippocampus inactivation we analyzed the frequency distribution for freezing scores in context A and context B. A single large peak was observed in context A (Figure 4E), while multiple peaks were observed in context B (Figure 4F). The mean of the first peak in context B was 16.7, which correlates with a discrimination index of 0.74. The mean of the second peak was 66.1, which correlates with a discrimination index of 0.53. Therefore, mice from the first distribution should be able to discriminate between contexts while those in the second distribution should not. Accordingly, we chose the intersection of these curves (42% freezing) as our threshold to distinguish between mice belonging to each of these two groups. Mice that froze less than this value in context B were categorized as discriminators and those that froze more were categorized as generalizers.
The retrieval of precise context memories requires the hippocampus
In the next experiment, mice were implanted with guide cannule that targeted the dorsal hippocampus. They were then trained in context A and tested in context B 14 days later (Figure 5A). Using the threshold from our previous analysis, we divided the mice into two groups: discriminators and generalizers (Figure 5B). The next day all of the animals were tested in the original training environment. Half of the animals in each group received an infusion of saline into the dorsal hippocampus before this test while the others received CNQX. We found that hippocampus inactivation with CNQX selectively impaired memory retrieval in mice that could discriminate between contexts, while those that generalized were not affected (significant group × drug interaction (F (1, 33) = 4.768, p < .05) (Figure 5C). These results demonstrate that the hippocampus plays a selective role in the retrieval of detailed context memories. This effect was independent of memory age as all animals were tested at 14 days.
Figure 5. The hippocampus is required for the retrieval of detailed but not generalized context memories.
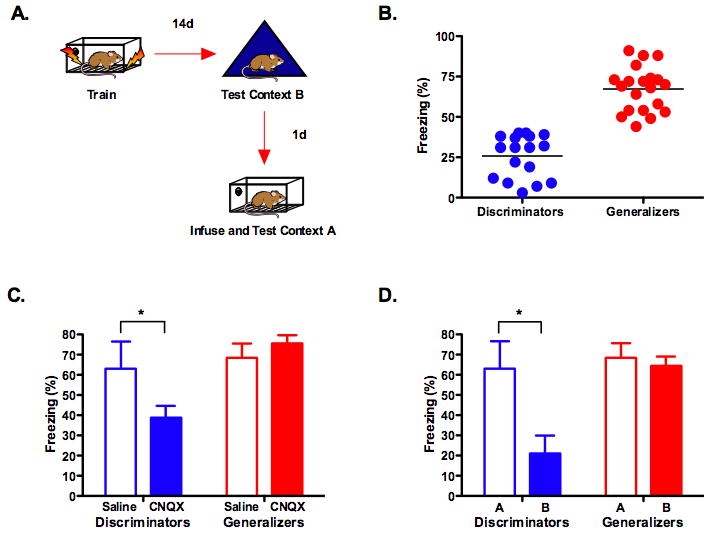
(a) Experimental design. (b) Fourteen days after training mice were tested in context B. Following this test, animals were divided into two groups: discriminators and generalizers. (c) Inactivation of the dorsal hippocampus with CNQX impaired memory retrieval in the discriminators (saline n =4, CNQX n = 13) (p < .05) but had no effect in the generalizers (saline n =10, CNQX n = 10) (p > .05). (d) An analyses of freezing scores in mice that received saline confirmed that discriminators were able to distinguish between context A and context B (p < .05) while generalizers could not (p >.05).
To confirm that our threshold for categorization was valid we compared freezing scores in context A and context B for mice that received saline infusions (Figure 5D). As predicted, the discriminators froze substantially more in the training context than in the novel environment while the generalizers froze the same amount in both contexts (significant group × context interaction (F (1, 12) = 7.762, p < .05). This difference could not be attributed to disparate levels of learning or performance as freezing in the training context was identical in both groups (F < 1).
To further validate these results we analyzed the same data set using regression analysis. Similar to our previous experiment, we found no relationship between freezing in context A and the discrimination index (slope not significantly different from zero, F (1, 35) = 2.238, p > .05, r2 = .06) (Figure 6A) but a strong relationship between freezing in context B and discrimination (slope significantly different from zero, F (1, 35) = 20.54, p < .05, r2 = .36) (Figure 6B). Next, we examined the relationship between freezing in context A and context B in mice that received infusions of saline or CNQX. In the saline mice, no relationship was observed between these variables (slope not significantly different from zero, F (1, 12) = 1.251, p > .05, r2 = .09) (Figure 6C). In contrast, there was a significant relationship between freezing in context A and context B in mice that received infusions of CNQX (slope significantly different from zero, F (1, 21) = 15.12, p < .05, r2 = .41) (Figure 6D). The less animals froze in context B the more their freezing scores were reduced by CNQX when tested in context A. This relationship is not simply a performance artifact (more/less freezing in context B leads to more/less freezing in context A) as is it was not observed in mice that received infusions of saline. This analysis strengthens our conclusion that the hippocampus plays a selective role in the retrieval of detailed context fear memories.
Figure 6. Generalization predicts which mice will be affected by hippocampus inactivation.
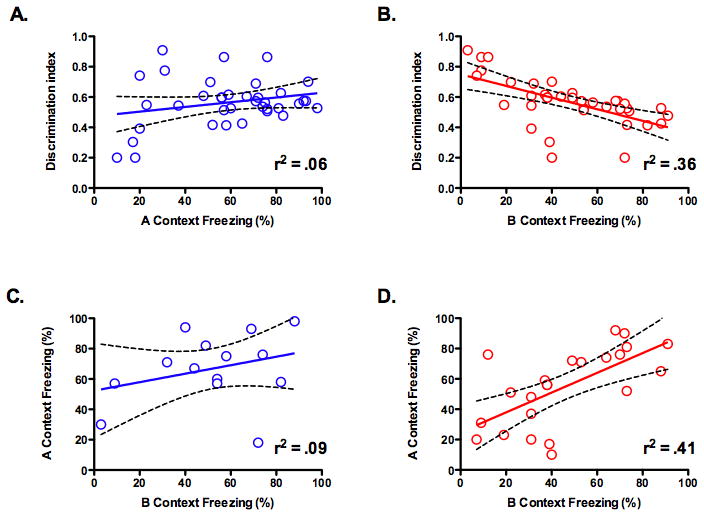
(a) Linear regression analysis found no relationship between freezing in context A and the discrimination index (p > .05). (b) Linear regression analysis found a significant relationship between freezing in context B and the discrimination index (p < .05). (c) Linear regression analysis found no relationship between freezing in context B and freezing in context A for mice that received saline infusions (p > .05). (d) Linear regression analysis found a significant relationship between freezing in context B and freezing in context A for mice that received CNQX infusions (p > .05).
An alternative account of the data presented in Figures 5 and 6 is that inactivation of the hippocampus reduces freezing by changing the internal state of the animal and causing a context-shift. Mice that are sensitive to a shift in the external context (i.e. discriminators) may also be more sensitive to an internal shift caused by drug infusion. While plausible, the inability of this account to explain the results of our other experiments or the findings of previous studies makes it unlikely. For example, in Figure 4 we show that IEG expression is reduced in the hippocampus as generalization to context B increases. This finding is consistent with our hypothesis that generalized context fear memories are not retrieved by the hippocampus. In contrast, a context-shift account makes no predictions about hippocampus activity during memory retrieval nor does it explain the decrease in IEG expression that accompanies generalization. In addition, previous work from our lab has shown that context discrimination depends on the hippocampus (Frankland et al., 1998). In the absence of this structure, mice show significantly increased generalization to a non-shocked environment. Consistent with this finding, fluorescent in situ hybridization (FISH) studies have shown that exposure to two distinct contexts activates non-overlapping groups of neurons in the hippocampus (Guzowski et al., 1999). Together, these results demonstrate that a) cells in the hippocampus are able to detect external context-shifts and b) these cells are essential for an animal to utilize contextual information and discriminate between environments. The implication is that the hippocampus plays an important role in detecting and encoding unique environments, a fact that is consistent with our hypothesis. The data are inconsistent with the idea that hippocampus inactivation impairs freezing because it produces a large state change. Instead, this structure plays a crucial role in the encoding of unique environments, which is in turn necessary to detect shifts in context.
Discussion
In summary, our data help to resolve the current debate about the role of the hippocampus in memory retrieval. First, we demonstrate that the dorsal hippocampus plays a permanent role in the retrieval of detailed context fear memories. Second, we show that context memories tend to lose details with the passage of time, and when they do so they can be retrieved without this structure. Memories that retain details continue to be retrieved by the hippocampus. Therefore, study designs that promote the retention of detailed memories should find that the hippocampus plays a permanent role in their retrieval. In contrast, conditions that result in memory generalization across time will lead to a temporal gradient of hippocampal involvement; this structure will be important shortly after training but not later. Both of these findings have been observed in the animal literature, although the quality of memory was not examined 3, 8, 13.
The current results can also explain why memory does not become independent of the hippocampus in tasks like the Morris watermaze 37. Unlike contextual fear conditioning, successful performance in the watermaze requires the retrieval of a precise memory about the spatial location of the platform. An inability to remember this specific information will impair searching during the probe test 38. Consequently, if hippocampus lesions impair the retrieval of detailed spatial information then both recent and remote memory will be disrupted in this task. In contrast, the current experiments demonstrate that retrieval of an imprecise context memory is sufficient to support freezing. As a result, context fear conditioning can be expressed without the hippocampus 5, 39.
The results of our study are consistent with a transformation account of memory consolidation, which argues that episodic memories lose details and become more schematic as they are permanently stored in regions of the neocortex 29, 30, 40. In contrast, the standard model of consolidation states that detailed features of episodic memories are retained over time 40, 41. This distinction has proven difficult to resolve even though the content of remote episodic memory has been studied in some detail in humans. Several studies have observed a decrease in the number of details recalled by subjects with hippocampus damage while others have found no change 17, 41, 42. However, the number of details recalled may not reflect memory accuracy. As autobiographical memories get older, subjects are more likely to remember and believe false details about these events 43, 44. Given this fact, future studies that examine memory accuracy rather than the number of generated details will be informative. It may be the case (with the use of more sensitive measuring devices) that the accurate recall of detailed information activates the hippocampus more than detailed, but inaccurate, information.
A potential interpretation of the current results is that generalized context fear memories are similar to semantic memories. Semantic memories, however, are thought to represent the extraction of general features from multiple learning events and our mice were trained with only a single conditioning trial 20. Although semantic memories are often formed across many trials, the studies described above demonstrate that specific episodes can become more schematic and “semantic-like” over time 40, 45. This is thought to occur as regions of the neocortex extract the general features of the original episodic memory as it is repeatedly retrieved and replayed over time 40, 45. A similar replay process occurs for hippocampus-dependent memories in rodents and could produce a related effect; the extraction of general contextual features that occur during multiple replay events 21, 46, 47.
Although the current results demonstrate that hippocampus-independence is related to memory quality, other important factors have also been identified. For example, studies with rats have shown that the incorporation of new information into existing frameworks or schemas allows it to become independent of the hippocampus within twenty-four hours 48. This suggests that the rate of memory consolidation depends on the prior experience of the subjects. Interestingly, we found that mice undergo the shift from precise to general memories at different rates. In a small number of animals, memory for the training context appeared to be generalized when tested as early as 24-hours after training, while in others it remained precise even after a month. Distinguishing between discriminators and generalizers allowed us to test hippocampal dependency in each of these groups. These results suggest that future studies will be able to identify specific factors, like environmental experience, that influence the rate of memory consolidation.
A recent paper found that with extended training, mice with hippocampus damage are able to discriminate between contexts at remote time points, although freezing performance is significantly reduced 16. Similarly, when animals are given extensive experience in a complex environment (e.g. 3 months) they are able to form and retain spatial memories following hippocampus lesions 49, 50. These data suggest that structures outside the hippocampus can be recruited to support the retrieval of precise memories under some conditions 5, 39.
Experimental Procedures
Subjects
F1 hybrids were generated for all experiments by breeding C57BL/6 (Taconic) males and 129SvE (Jackson) females. Mice ranged from 3 to 6 months of age, were group housed with free access to food and tap water and maintained on a 12:12 h light:dark cycle in the Herbert L. Washington Vivarium in the Department of Psychology at UCLA. All experiments were performed during the light phase of the cycle.
Fear conditioning
The general apparatus and procedures used in these experiments have been described previously19. In all experiments, the mice were allowed to explore the training environment (context A) for 2 minutes before shock was delivered. Thirty seconds after the last shock, the mice were removed from the training context and returned to their home cage. All animals received a 5-minute context test after training and freezing behavior was scored as previously described19. Mice received five footshocks (2 sec, 0.75 mA) in Figure 1 and a single footshock (2 sec, 0.5 mA) in Figures 2-5. Testing was conducted in context A, context B (described previously19) or context C. Context C consisted of a small plastic cage (27.5 cm × 17 cm × 12.5 cm) with new bedding, located in a novel, dimly lit room down the hall from the training environment.
RT-PCR
Brains were extracted and flash frozen on dry ice, and sliced at 2 mm sections in a brain block kept at -20 °C and the dorsal hippocampus was microdissected. Total RNA from the dorsal hippocampus was extracted by RNeasy Mini Kit (Qiagen, Valencia, CA, USA) and treated with DNase (Qiagen). Total RNA was reverse-transcribed into cDNA using oligo (dT) 20 primers and Superscipt III First-Strand Synthesis System (Invitrogen, Carlsbad, CA, USA). Expression of Arc and c-fos were determined by real-time PCR. Arc and c-fos primers were designed by Primer Express 2.0 (Applied Biosystems) and the following primer sequences were used: Arc, 5′ TATTCAGGCTGGGTCCTGTC 3′ (forward) and 5′ TGGAGCAGCTTATCCAGAGG 3′ (reverse); C-fos, 5′ TCACCCTGCCCCTTCTCA 3′ (forward) and 5′ CACGTTGCTGATGCTCTTGAC 3′ (reverse) Zif-268 5′TTGCCGATGGCTTGACATG3′ (forward) and 5′TAAGGCTAAGGTGAGCGTGTCC3′ (reverse). Real-Time PCR was performed in an ABI PRISM 7900HT Sequence Detection System (Applied Biosystems, Foster City, CA, USA). Cycling parameters were as follows: Initial denaturation at 95°C for 3 min followed by 40 cycles (95°C for 30 sec and 60°C for 1 min). The data were analyzed by the 2(x002C6)CT method 51 and mRNA expression is shown relative to that observed in homecage controls. The expression of 36B4 and HPRT genes were measured and used as housekeeping controls for all samples.
FISH
Fluorescent-in situ hybridization was performed as previously described27. Mice were trained, tested and then immediately sacrificed with isoflurane. Brains were extracted, flash frozen on dry ice and stored at -80° C until sectioning. 20 μM sections were taken on a cryostat (-20° C) and mounted on slides. Fluorescein-labeled riboprobes were made with commercial transcription kits (MaxiScript; Ambion, Austin, TX) and RNA labeling mixes (Roche Products, Hertforshire, UK). The H1a antisense riboprobe was generated using an H1a cDNA clone (generous gift from Paul Worley) and was directed to the 4.4 kb 3′ untranslated region (UTR) of the H1a mRNA27. Single-label FISH was performed as previously described27, 52, 53. Fluorescein-labeled H1a probe was detected with anti-fluorescein HRP (Roche Products) and a cyanine-5 substrate kit (CY5 DirectFISH; PerkinElmer Life Sciences). Nuclei were counterstained with 4′,6′-diamidino-2-phenylindole (DAPI; Vector Laboratories, Burlingame, CA).
Slides were imaged with an Olympus FluoView 1000 Laser Scanning Confocal Microscope (LSCM). Laser line (633 nm) from HeNe was used for imaging Cy5 and 700 nm femtosecond pulses from Ti:Saphire (Mai Tai, Spectra Physics) was used for two photon excitation of DAPI. Olympus 60×/1.2 oil-immersion objective lens was used for imaging the CA1 region of the dorsal hippocampus. Laser intensity, pinhole size, PMT dynode voltage, amplifier gain and offset were kept constant for imaging of all slides. Slices from the CA1 region of hippocampal were Z sectioned in 0.48 μM optical sections from top to bottom. Twenty sections from the middle of the slice (approximately 9.6um) were used for analysis. The software ImageJ (ImageJ 1.42g, NIH, USA) was used to obtain the distribution of homer puncta and set a threshold that separated homer signal from background noise. Only cells completely localized in the middle 20 optical sections and with homer particles above threshold in max projection were counted as homer positive cells. Total cells numbers were counted with DAPI staining. Only whole cells were counted. An average of 831 cells in the dorsal CA1 were counted for each mouse. The results are shown as the percentage of homer positive cells per total counted cells. All the image analysis was done blind.
Intra-hippocampal infusions
Mice were anesthetized with sodium pentobarbital (90 mg/kg) and mounted in a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA). The scalp of each animal was incised and retracted, and the skull was adjusted to place bregma and lambda in the same horizontal plane. Small burr holes were drilled at the appropriate injection sites. Plastic guide cannule (22 gauge; Plastics One, Roanoke, VA) were inserted bilaterally at the following positions relative to bregma (mm): AP: -2, ML: ±1.5, DV: -1 and affixed with dental cement (Harry J. Bosworth Company, Skokie, IL). Dummy cannule (28 gauge) were inserted into the guide cannule following surgery. Mice were allowed to recover for one week prior to behavioral testing. Twenty minutes prior to testing, the dummy cannule were removed and replaced with injection cannule (28 gauge) that projected an additional 0.5 mm from the tip of the guide cannule. CNQX (Sigma) (.83 mg/ml) or saline (0.9%) was infused into the hippocampus (.5 ul/side; 0.1 ul/minute). The injectors were left in place for 2 minutes after the end of the infusion to allow for diffusion. The mice were then returned to their homecage until testing.
Histology
Histological verification of the cannula locations was performed at the end of behavioral testing. Mice were perfused transcardially with 0.9% saline, followed by 4% PFA. After extraction from the skull, the brains were postfixed in 4% PFA and then transferred to a 30% sucrose solution until sectioning. Coronal sections (40 μm thick, taken every 120 μm) were cut on a cryostat (−16°C) and mounted on glass microscope slides. After drying, the sections were stained with cresyl violet to identify neuronal cell bodies. Cannula tips were verified by visual inspection of the stained sections reconstructed on the mouse Allen Reference Atlas 54. We were unable to analyze the brains of four mice from each CNQX group because of damaged sustained during the extraction and/or slicing process.
Supplementary Material
Supplemental Figure 1: Cannula placement in the dorsal hippocampus. (a) Cannula tip locations (anterior/posterior coordinates (mm) relative to bregma) for mice receiving intra-hippocampal infusions of CNQX. (b) Representative cannula placement in the dorsal hippocampus.
Acknowledgments
We would like to thank Yijun Cui, Jaejoon Won, Yong-Seok Lee, John Guzowski, Teiko Miyashita, Paul Worley, Connie Smith-Hicks, and Sheena Josselyn for their assistance. This work was funded by grants from the National Institute of Health (NIH) to AJS (R37-AG013622 and P50-MH0779720) and BJW (NRSA AG023403).
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
References
- 1.Squire LR, Stark CE, Clark RE. The medial temporal lobe. Annu Rev Neurosci. 2004;27:279–306. doi: 10.1146/annurev.neuro.27.070203.144130. [DOI] [PubMed] [Google Scholar]
- 2.Moscovitch M, Nadel L, Winocur G, Gilboa A, Rosenbaum RS. The cognitive neuroscience of remote episodic, semantic and spatial memory. Curr Opin Neurobiol. 2006;16:179–190. doi: 10.1016/j.conb.2006.03.013. [DOI] [PubMed] [Google Scholar]
- 3.Anagnostaras SG, Maren S, Fanselow MS. Temporally graded retrograde amnesia of contextual fear after hippocampal damage in rats: within-subjects examination. J Neurosci. 1999;19:1106–1114. doi: 10.1523/JNEUROSCI.19-03-01106.1999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Frankland PW, Bontempi B. The organization of recent and remote memories. Nat Rev Neurosci. 2005;6:119–130. doi: 10.1038/nrn1607. [DOI] [PubMed] [Google Scholar]
- 5.Wiltgen BJ, Sanders MJ, Anagnostaras SG, Sage JR, Fanselow MS. Context fear learning in the absence of the hippocampus. J Neurosci. 2006;26:5484–5491. doi: 10.1523/JNEUROSCI.2685-05.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Squire LR. Memory and the hippocampus: a synthesis from findings with rats, monkeys, and humans. Psychol Rev. 1992;99:195–231. doi: 10.1037/0033-295x.99.2.195. [DOI] [PubMed] [Google Scholar]
- 7.Squire LR, Zola-Morgan S. The medial temporal lobe memory system. Science. 1991;253:1380–1386. doi: 10.1126/science.1896849. [DOI] [PubMed] [Google Scholar]
- 8.Kim JJ, Fanselow MS. Modality-specific retrograde amnesia of fear. Science. 1992;256:675–677. doi: 10.1126/science.1585183. [DOI] [PubMed] [Google Scholar]
- 9.Rekkas PV, Constable RT. Evidence that autobiographic memory retrieval does not become independent of the hippocampus: an fMRI study contrasting very recent with remote events. J Cogn Neurosci. 2005;17:1950–1961. doi: 10.1162/089892905775008652. [DOI] [PubMed] [Google Scholar]
- 10.Steinvorth S, Corkin S, Halgren E. Ecphory of autobiographical memories: an fMRI study of recent and remote memory retrieval. Neuroimage. 2006;30:285–298. doi: 10.1016/j.neuroimage.2005.09.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lein ES, et al. Genome-wide atlas of gene expression in the adult mouse brain. Nature. 2007;445:168–176. doi: 10.1038/nature05453. [DOI] [PubMed] [Google Scholar]
- 12.Lehmann H, Lacanilao S, Sutherland RJ. Complete or partial hippocampal damage produces equivalent retrograde amnesia for remote contextual fear memories. Eur J Neurosci. 2007;25:1278–1286. doi: 10.1111/j.1460-9568.2007.05374.x. [DOI] [PubMed] [Google Scholar]
- 13.Sutherland RJ, O'Brien J, Lehmann H. Absence of systems consolidation of fear memories after dorsal, ventral, or complete hippocampal damage. Hippocampus. 2008;18:710–718. doi: 10.1002/hipo.20431. [DOI] [PubMed] [Google Scholar]
- 14.Sutherland RJ, et al. Retrograde amnesia after hippocampal damage: recent vs. remote memories in two tasks. Hippocampus. 2001;11:27–42. doi: 10.1002/1098-1063(2001)11:1<27::AID-HIPO1017>3.0.CO;2-4. [DOI] [PubMed] [Google Scholar]
- 15.Maren S, Aharonov G, Fanselow MS. Neurotoxic lesions of the dorsal hippocampus and Pavlovian fear conditioning in rats. Behav Brain Res. 1997;88:261–274. doi: 10.1016/s0166-4328(97)00088-0. [DOI] [PubMed] [Google Scholar]
- 16.Wang SH, Teixeira CM, Wheeler AL, Frankland PW. The precision of remote context memories does not require the hippocampus. Nat Neurosci. 2009;12:253–255. doi: 10.1038/nn.2263. [DOI] [PubMed] [Google Scholar]
- 17.Moscovitch M, et al. Functional neuroanatomy of remote episodic, semantic and spatial memory: a unified account based on multiple trace theory. J Anat. 2005;207:35–66. doi: 10.1111/j.1469-7580.2005.00421.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Steinvorth S, Levine B, Corkin S. Medial temporal lobe structures are needed to re-experience remote autobiographical memories: evidence from H.M. and W.R. Neuropsychologia. 2005;43:479–496. doi: 10.1016/j.neuropsychologia.2005.01.001. [DOI] [PubMed] [Google Scholar]
- 19.Wiltgen BJ, Silva AJ. Memory for context becomes less specific with time. Learn Mem. 2007;14:313–317. doi: 10.1101/lm.430907. [DOI] [PubMed] [Google Scholar]
- 20.McClelland JL, McNaughton BL, O'Reilly RC. Why there are complementary learning systems in the hippocampus and neocortex: insights from the successes and failures of connectionist models of learning and memory. Psychol Rev. 1995;102:419–457. doi: 10.1037/0033-295X.102.3.419. [DOI] [PubMed] [Google Scholar]
- 21.O'Reilly RC, Rudy JW. Conjunctive representations in learning and memory: principles of cortical and hippocampal function. Psychol Rev. 2001;108:311–345. doi: 10.1037/0033-295x.108.2.311. [DOI] [PubMed] [Google Scholar]
- 22.Frankland PW, Bontempi B, Talton LE, Kaczmarek L, Silva AJ. The involvement of the anterior cingulate cortex in remote contextual fear memory. Science. 2004;304:881–883. doi: 10.1126/science.1094804. [DOI] [PubMed] [Google Scholar]
- 23.Maviel T, Durkin TP, Menzaghi F, Bontempi B. Sites of neocortical reorganization critical for remote spatial memory. Science. 2004;305:96–99. doi: 10.1126/science.1098180. [DOI] [PubMed] [Google Scholar]
- 24.Bontempi B, Laurent-Demir C, Destrade C, Jaffard R. Time-dependent reorganization of brain circuitry underlying long-term memory storage. Nature. 1999;400:671–675. doi: 10.1038/23270. [DOI] [PubMed] [Google Scholar]
- 25.Anagnostaras SG, Gale GD, Fanselow MS. Hippocampus and contextual fear conditioning: recent controversies and advances. Hippocampus. 2001;11:8–17. doi: 10.1002/1098-1063(2001)11:1<8::AID-HIPO1015>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- 26.Vazdarjanova A, Guzowski JF. Differences in hippocampal neuronal population responses to modifications of an environmental context: evidence for distinct, yet complementary, functions of CA3 and CA1 ensembles. J Neurosci. 2004;24:6489–6496. doi: 10.1523/JNEUROSCI.0350-04.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Vazdarjanova A, McNaughton BL, Barnes CA, Worley PF, Guzowski JF. Experience-dependent coincident expression of the effector immediate-early genes arc and Homer 1a in hippocampal and neocortical neuronal networks. J Neurosci. 2002;22:10067–10071. doi: 10.1523/JNEUROSCI.22-23-10067.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Biedenkapp JC, Rudy JW. Context preexposure prevents forgetting of a contextual fear memory: implication for regional changes in brain activation patterns associated with recent and remote memory tests. Learn Mem. 2007;14:200–203. doi: 10.1101/lm.499407. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Winocur G, Frankland PW, Sekeres M, Fogel S, Moscovitch M. Changes in context-specificity during memory reconsolidation: selective effects of hippocampal lesions. Learn Mem. 2009;16:722–729. doi: 10.1101/lm.1447209. [DOI] [PubMed] [Google Scholar]
- 30.Winocur G, Moscovitch M, Sekeres M. Memory consolidation or transformation: context manipulation and hippocampal representations of memory. Nat Neurosci. 2007;10:555–557. doi: 10.1038/nn1880. [DOI] [PubMed] [Google Scholar]
- 31.Moyer JR, Jr, Thompson LT, Disterhoft JF. Trace eyeblink conditioning increases CA1 excitability in a transient learning-specific manner. J Neurosci. 1996;16:5536–5546. doi: 10.1523/JNEUROSCI.16-17-05536.1996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.McKay BM, Matthews EA, Oliveira FA, Disterhoft JF. Intrinsic neuronal excitability is reversibly altered by a single experience in fear conditioning. J Neurophysiol. 2009;102:2763–2770. doi: 10.1152/jn.00347.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Rampon C, et al. Enrichment induces structural changes and recovery from nonspatial memory deficits in CA1 NMDAR1-knockout mice. Nat Neurosci. 2000;3:238–244. doi: 10.1038/72945. [DOI] [PubMed] [Google Scholar]
- 34.Bourtchuladze R, et al. Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell. 1994;79:59–68. doi: 10.1016/0092-8674(94)90400-6. [DOI] [PubMed] [Google Scholar]
- 35.Chen C, Rainnie DG, Greene RW, Tonegawa S. Abnormal fear response and aggressive behavior in mutant mice deficient for alpha-calcium-calmodulin kinase II. Science. 1994;266:291–294. doi: 10.1126/science.7939668. [DOI] [PubMed] [Google Scholar]
- 36.Barrientos RM, O'Reilly RC, Rudy JW. Memory for context is impaired by injecting anisomycin into dorsal hippocampus following context exploration. Behav Brain Res. 2002;134:299–306. doi: 10.1016/s0166-4328(02)00045-1. [DOI] [PubMed] [Google Scholar]
- 37.Clark RE, Broadbent NJ, Squire LR. Hippocampus and remote spatial memory in rats. Hippocampus. 2005;15:260–272. doi: 10.1002/hipo.20056. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Morris RG. Spatial Localization Does Not Require the Presence of Local Cues. Learning and Motivation. 1981;12:239–260. [Google Scholar]
- 39.Frankland PW, Cestari V, Filipkowski RK, McDonald RJ, Silva AJ. The dorsal hippocampus is essential for context discrimination but not for contextual conditioning. Behav Neurosci. 1998;112:863–874. doi: 10.1037//0735-7044.112.4.863. [DOI] [PubMed] [Google Scholar]
- 40.Winocur G, Moscovitch M, Bontempi B. Memory formation and long-term retention in humans and animals: Convergence towards a transformation account of hippocampal-neocortical interactions. Neuropsychologia. 2010 doi: 10.1016/j.neuropsychologia.2010.04.016. [DOI] [PubMed] [Google Scholar]
- 41.Kirwan CB, Bayley PJ, Galvan VV, Squire LR. Detailed recollection of remote autobiographical memory after damage to the medial temporal lobe. Proc Natl Acad Sci U S A. 2008;105:2676–2680. doi: 10.1073/pnas.0712155105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Gilboa A, Winocur G, Grady CL, Hevenor SJ, Moscovitch M. Remembering our past: functional neuroanatomy of recollection of recent and very remote personal events. Cereb Cortex. 2004;14:1214–1225. doi: 10.1093/cercor/bhh082. [DOI] [PubMed] [Google Scholar]
- 43.Mendelsohn A, Furman O, Navon I, Dudai Y. Subjective vs. documented reality: a case study of long-term real-life autobiographical memory. Learn Mem. 2009;16:142–146. doi: 10.1101/lm.1157709. [DOI] [PubMed] [Google Scholar]
- 44.Barclay CR, Wellman HM. Accuracies and inaccuracies in autobiographical memories. Journal of Memory and Language. 1986;25:93–103. [Google Scholar]
- 45.Nadel L, Winocur G, Ryan L, Moscovitch M. Systems consolidation and hippocampus: two views. Debates in Neuroscience. 2007;1:55–66. [Google Scholar]
- 46.Foster DJ, Wilson MA. Reverse replay of behavioural sequences in hippocampal place cells during the awake state. Nature. 2006;440:680–683. doi: 10.1038/nature04587. [DOI] [PubMed] [Google Scholar]
- 47.Louie K, Wilson MA. Temporally structured replay of awake hippocampal ensemble activity during rapid eye movement sleep. Neuron. 2001;29:145–156. doi: 10.1016/s0896-6273(01)00186-6. [DOI] [PubMed] [Google Scholar]
- 48.Tse D, et al. Schemas and memory consolidation. Science. 2007;316:76–82. doi: 10.1126/science.1135935. [DOI] [PubMed] [Google Scholar]
- 49.Winocur G, Moscovitch M, Rosenbaum RS, Sekeres M. An investigation of the effects of hippocampal lesions in rats on pre- and postoperatively acquired spatial memory in a complex environment. Hippocampus. 2010 doi: 10.1002/hipo.20721. [DOI] [PubMed] [Google Scholar]
- 50.Winocur G, Moscovitch M, Fogel S, Rosenbaum RS, Sekeres M. Preserved spatial memory after hippocampal lesions: effects of extensive experience in a complex environment. Nat Neurosci. 2005;8:273–275. doi: 10.1038/nn1401. [DOI] [PubMed] [Google Scholar]
- 51.Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3:1101–1108. doi: 10.1038/nprot.2008.73. [DOI] [PubMed] [Google Scholar]
- 52.Guzowski JF, McNaughton BL, Barnes CA, Worley PF. Environment-specific expression of the immediate-early gene Arc in hippocampal neuronal ensembles. Nat Neurosci. 1999;2:1120–1124. doi: 10.1038/16046. [DOI] [PubMed] [Google Scholar]
- 53.Guzowski JF, Worley PF. Cellular compartment analysis of temporal activity by fluorescence in situ hybridization (catFISH) Curr Protoc Neurosci. 2001;Chapter 1(Unit 1 8) doi: 10.1002/0471142301.ns0108s15. [DOI] [PubMed] [Google Scholar]
- 54.Dong HW. The Allen Reference Atlas. Wiley; 2009. [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplemental Figure 1: Cannula placement in the dorsal hippocampus. (a) Cannula tip locations (anterior/posterior coordinates (mm) relative to bregma) for mice receiving intra-hippocampal infusions of CNQX. (b) Representative cannula placement in the dorsal hippocampus.