
Figure 5.
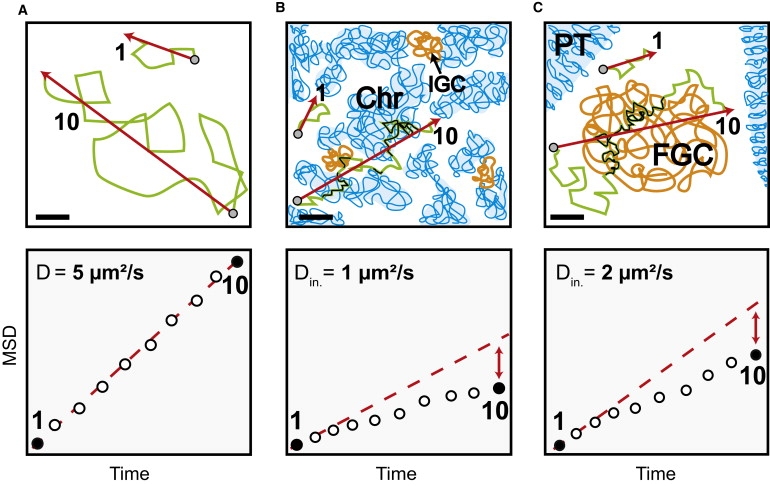
Impact of the local nuclear environment on the mobility of large particles and corresponding MSD-time plots (lower lane). (A) Determination of MSD in buffer. (Lower lane) The MSD is linear due to Brownian motion. (B) Mobility in mammalian cell nuclei. The unfolded chromatin (Chr) and interchromatin granule clusters reduce particle mobility on long timescales (t = 10), but not on short timescales (t = 1), when particles diffuse within interchromatin channels with an initial diffusion coefficient due to the effective nucleoplasmic viscosity. At long times the traveled distance is reduced due to frequent collisions with mobility barriers or unspecific and specific binding events (black sections of the trajectories) due to unfolded chromatin and interchromatin structures leading to a nonlinear MSD-time dependence (lower lane). (C) Mobility in salivary gland cell nuclei. The interphase chromatin is compacted into polytene chromosomes (PT) leaving vast regions devoid of chromatin. The diffusion on short timescales (t = 1) is due to the effective local viscosity as in mammalian cell nuclei. (Lower lane) The particle mobility at long times and large spatial scales (t = 10) shows a nonlinear MSD-time dependence. Because this cannot be due to chromatin it implies the presence of another extended nuclear structure influencing the mobility (black trajectory parts) of endogenous molecules (BR2.1 mRNPs) and inert tracer particles (500 kDa dextrans). Presumably, these structures correspond to FGCs (40). Initial diffusion coefficients for 500 kDa dextrans are shown in the lower lane. Bars, 2 μm.