

Abstract
Isotopic studies of wild primates have used a wide range of tissues to infer diet and model the foraging ecologies of extinct species. The use of mismatched tissues for such comparisons can be problematic because differences in amino acid compositions can lead to small isotopic differences between tissues. Additionally, physiological and dietary differences among primate species could lead to variable offsets between apatite carbonate and collagen. To improve our understanding of the isotopic chemistry of primates, we explored the apparent enrichment (ε*) between bone collagen and muscle, collagen and fur or hair keratin, muscle and keratin, and collagen and bone carbonate across the primate order. We found that the mean ε* values of proteinaceous tissues were small (≤1‰), and uncorrelated with body size or phylogenetic relatedness. Additionally, ε* values did not vary by habitat, sex, age, or manner of death. The mean ε* value between bone carbonate and collagen (5.6 ± 1.2‰) was consistent with values reported for omnivorous mammals consuming monoisotopic diets. These primate-specific apparent enrichment values will be a valuable tool for cross-species comparisons. Additionally, they will facilitate dietary comparisons between living and fossil primates.
Electronic supplementary material
The online version of this article (doi:10.1007/s00442-010-1701-6) contains supplementary material, which is available to authorized users.
Keywords: Stable isotope, Keratin, Muscle, Collagen, Apatite, Carbonate
Introduction
Stable isotope ratios in animal tissues vary with diet, habitat, and environmental conditions, and are often used to assess the foraging ecology and habitat preferences of living and extinct species (West et al. 2006). These studies have varied methodologically, using a range of tissues. For instance, the diets of wild primates have been assessed using isotope values from hair (e.g., Schoeninger et al. 1997, 2006), tooth enamel (e.g., Codron et al. 2005; Fourie et al. 2008; Smith et al. 2010), bone (e.g., Ambrose and DeNiro 1986; Thackeray et al. 1996; Smith et al. 2010), and feces (e.g., Codron et al. 2006). These data, in turn, have been used to inform paleo-ecological models of extinct species, including early human ancestors (e.g., Thackeray et al. 1996; Codron et al. 2005; Sponheimer et al. 2006, 2010). Due to tissue preservation issues, these studies have frequently had to use different tissues in their modern and ancient comparisons.
Although the availability and state of preservation of specimens are practical constraints, it can be problematic to compare the isotopic composition of different tissues for two reasons. First, carbon and nitrogen isotope values of proteinaceous tissues can differ within an animal because each tissue has a unique amino acid (AA) composition, and the AAs themselves vary isotopically (e.g., Hare et al. 1991; Styring et al. 2010). Second, when studying fossils, researchers generally use the carbonate fraction of biological apatite. The isotopic difference [hereafter termed apparent enrichment, ε*(defined below)] between carbon in organic tissues, such as collagen in bone or dentin, and carbon in the carbonate in bone or tooth apatite varies with both digestive physiology and dietary macromolecular composition (reviewed in Hedges 2003).
A wide range of dietary and gut physiological adaptations among primates could lead to differences in ε* values for both carbon and nitrogen that could in turn confound ecological or paleoecological interpretations. Many experiments have been conducted on rodents and pigs, but most were focused on carbon isotope differences between carbonate and collagen. Scarcely any work has examined the differences between proteinaceous tissues, let alone unconventional taxa. For instance, only one published study has focused on nonhuman primates (O’Regan et al. 2008) (Table 1). Accordingly, we compared the carbon and nitrogen isotope values in keratin, muscle protein, bone collagen, and bone carbonate (carbon only) for a diverse group of primate species (Table 2). We expected that differences in AA composition would drive ε* variation among proteinaceous tissues for both carbon and nitrogen, but that these differences would be small and consistent across individuals and species. We also expected that ε* values for carbonate versus collagen carbon would vary among species as a function of both diet and digestive physiology, and factors that correlate with these variables (body size, habitat, etc.).
Table 1.
Tissue-tissue carbon and nitrogen fractionation values from previous research on mammals
| Taxon | Dieta | n | Δ13C | Range | Δ15N | Range | References | |
|---|---|---|---|---|---|---|---|---|
| Collagen–keratin | ||||||||
| Mouse | Captive | Mixed | 72 | 2.9 ± 2.2 | 1.1, 7.1b | 1 | ||
| Mouse | Captive | Uniform | 24 | 2.5 ± 0.3 | 2.3, 2.7b | 1 | ||
| Wolf | Captive | Uniform | 18 | 0.4 ± 0.8 | −0.9, 2.3 | 0.3 ± 0.7 | −1.0, 1.7 | 2 |
| Human | Modern | Mixed | 8 | 1.5 ± 0.5 | 0.8, 2.2 | 0.9 ± 0.2 | 0.7, 1.1 | 3 |
| Macaque | Wild | Mixedc | 13 | 0.1 ± 1.1 | −1.5, 1.0b | 0.4 ± 0.3 | −0.3, 1.1 | 5 |
| Collagen–muscle | ||||||||
| Mouse | Captive | Mixed | 72 | 2.4 ± 0.4 | 1.7, 2.9b | 1 | ||
| Moused | Captive | Mixed | 2 | 3.7 ± 0.1 | 3.6, 3.8 | 1 | ||
| Mouse | Captive | Uniform | 24 | 2.2 ± 0.8 | 1.6, 2.7b | 1 | ||
| Moused | Captive | Uniform | 6 | 2.4 ± 0.6 | 1.8, 3.5 | 1 | ||
| Sheep | Domestic | Mixed | 2 | 4.1 ± 0.1 | 4.0, 4.1 | 6 | ||
| Pig | Domestic | Uniform | 20 | 1.1 ± 1.6 | −0.1, 2.2b | 0.9 ± 0.6 | 0.5, 1.3 | 7 |
| Wolf | Captive | Uniform | 18 | 1.5 ± 0.7 | 0.4, 3.1 | −0.5 ± 0.8 | −1.8, 1.8 | 2 |
| Gemsbok | Wild | Mixed | 1 | 0.7 | 6 | |||
| Hartabeest | Wild | Mixed | 1 | 1.9 | 6 | |||
| Impala | Wild | Mixed | 1 | 1.7 | 6 | |||
| Kudu | Wild | Uniform | 2 | 2.4 ± 0.6 | 1.9, 2.8 | 6 | ||
| Springbok | Wild | Mixed | 3 | 2.4 ± 0.6 | 1.8, 3.0 | 6 | ||
| Warthog | Wild | Mixed | 1 | 1.7 | 6 | |||
| Muscle–keratin | ||||||||
| Gerbil | Captive | Mixed | 37 | −2.3 ± 0.7 | −3.6, −1.3b | 8 | ||
| Mouse | Captive | Mixed | 72 | 0.5 ± 2.1 | −1.3, 4.5b | 1 | ||
| Mouse | Captive | Uniform | 24 | 0.4 ± 0.5 | 0.0, 0.7b | 1 | ||
| Mouse | Captive | All | −2.9e | 9 | ||||
| Mouse | Captive | All | 18 | 0.3e | 10 | |||
| Pig | Domestic | Uniform | 5 | 1.8 | −0.1 | 11 | ||
| Fox | Captive | Mixed | 20 | −1.5f | 0.2 ± 0.1 | 0.1, 0.2b | 12 | |
| Wolf | Captive | Uniform | 19 | −1.2 ± 0.4 | −2.2, 0.5 | 0.8 ± 0.5 | −0.2, 1.5 | 2 |
| Carbonate-collagen | ||||||||
| Mouse | Captive | Mixed | 72 | 4.7 ± 3.0 | 1.3, 8.7b | 1 | ||
| Rat | Captive | Mixed | 20 | 4.2 ± 4.4 | −0.8, 11.1b | 13 | ||
| Rat | Captive | Mixed | 18−60 | 7.2 ± 4.6 | 1.3, 11.3b | 14 | ||
| Mouse | Captive | Uniform | 24 | 5.9 ± 1.1 | 5.1, 6.7b | 1 | ||
| Rat | Captive | Uniform | 8 | 5.0 ± 0.6 | 4.5, 6.0 | 13 | ||
| Rat | Captive | Uniform | 3−10 | 5.7 | 14 | |||
| Pig | Domestic | Mixed | 5 | 7.5 ± 1.0 | 6.4, 9.1 | 15 | ||
| Pig | Domestic | Uniform | 1 | 6.0 | 15 | |||
| Herbivore | Wild | All | 6.8 | 16 | ||||
| Giraffe | Wild | Uniform | 4 | 6.9 ± 0.3 | 6.7, 7.4 | 18, 19 | ||
| Hartabeest | Wild | Mixed | 1 | 8.4 | 18 | |||
| Topi | Wild | Uniform | 1 | 10.3 | 19 | |||
| Deer | Wild | Mixed | 1 | 6.8 | 18 | |||
| Reindeer | Wild | Uniform | 8 | 8.5 ± 0.8 | 7.0, 9.5 | 20 | ||
| Llama | Wild | Uniform | 6 | 7.1 ± 0.3 | 6.6, 7.3 | 17 | ||
| Hippo | Wild | Mixed | 4 | 6.6 ± 0.7 | 5.8, 7.5 | 18, 19 | ||
| Zebra | Wild | Mixed | 2 | 9.0 ± 1.1 | 8.2, 9.7 | 18, 19 | ||
| Omnivore | Wild | All | 5.2 | 16 | ||||
| Macaque | Wild | Mixed | 11 | 5.7 ± 0.5 | 5.0, 6.1 | 5 | ||
| Carnivore | Wild | All | 4.3 | 16 | ||||
| Fur Seal | Wild | Uniform | 2 | 2.2 ± 0.8 | 1.6, 2.7 | 16 | ||
| Harbor Seal | Wild | Uniform | 4 | 2.4 ± 1.1 | 1.6, 4.1 | 20 | ||
| Harp Seal | Wild | Uniform | 4 | 3.6 ± 1.1 | 2.2, 4.5 | 20 | ||
References: (1) Tieszen and Fagre (1993); (2) Fox-Dobbs et al. (2007); (3) O’Connell et al. 2001; (4) O’Regan et al. (2008); (5) Vogel (1978); (6) Hare et al. (1991); (7) Tieszen et al. (1983); (8) DeNiro and Epstein (1978); (9) DeNiro and Epstein (1981); (10) Nardoto et al. (2006); (11) Roth and Hobson (2000); (12) Jim et al. (2004); (13) Ambrose and Norr (1993); (14) Howland et al. (2003); (15) Lee-Thorp et al. (1989); (16) Schoeninger and DeNiro (1982); (17) Sullivan and Krueger (1981); (18) Kellner and Schoeninger (2007); (19) Nelson et al. (1986)
aWhenever possible, animal diets were divided into “uniform” (consumed all C3 or all C4) or “mixed” (consumed a combination of C3, C4 or marine). Otherwise, we use the category “All”. Diets for wild animals, which were inferred by the primary authors from each study, were considered mixed if the primary diet source (e.g., C3 or C4) was ≤90%
bStandard deviations and ranges were calculated for captive groups fed similar diets, or wild groups living in different regions
cDietary information is not available for these animals. The authors argue that collagen δ13C values suggest that some individuals may have consumed some C4 resources. However, apatite δ13C values do not support C4 consumption. Because no comparative plant data are available from the respective habitats, it is not possible to validate or refute C4 consumption
dNursing mothers (n = 2) and suckling babies (n = 6)
eMean Δ values estimated using Datathief 12.0
fStandard deviation and range were not presented
Table 2.
Species, body mass of males and females, and provenience of specimens included in this study
| Family and species | Body mass (kg)a | Type | Provenanceb | |
|---|---|---|---|---|
| Male | Female | |||
| Lorisoidea | ||||
| Galago senegalensis mohili | 0.2 | 0.2 | Captive | 1 |
| Lemuroidea | ||||
| Avahi laniger | 1.0 | 1.3 | Wild | 2 |
| Cheirogaleus major | 0.4 | 0.4 | Wild | 2 |
| D. madagascariensis | 2.6 | 2.5 | Captive | 1 |
| Eulemur fulvus albifrons | 2.0 | 2.2 | Captive | 1 |
| E. fulvus rufus | 2.2 | 2.3 | Wild | 4 |
| E. macaco flavifrons | 2.4 | 2.5 | Captive | 1 |
| E. mongoz | 1.6 | 1.6 | Captive | 1 |
| L. catta | 3.6 | 3.5 | Captive | 1 |
| Indri indri | 5.6 | 6.3 | Wild | 4 |
| Microcebus griseorufus | 0.05 | 0.06 | Wild | 3 |
| M. murinus | 0.1 | 0.1 | Captive | 1 |
| M. rufus | 0.1 | 0.1 | Wild | 2 |
| Propithecus coquereli verreauxi | 3.7 | 4.3 | Captive | 1 |
| P. diadema | 5.9 | 6.3 | Wild | |
| P. verreauxi | 3.3 | 3.0 | Wild | 3 |
| V. variagata | 3.5 | 3.5 | Captive | 1 |
| Ceboidea | ||||
| Alouatta paliatta | 6.5 | 4.2 | Wild | 6–8 |
| A. geoffroyi | 7.8 | 7.3 | Wild | 6,7 |
| C. capucinus | 3.7 | 2.5 | Wild | 6,7 |
| Cercopithecoidea | ||||
| Cercopithecus ascanius | 3.7 | 2.9 | Wild | 9 |
| Chlorocebus aethiops | 5.0 | 3.5 | Captive | 10 |
| Lophocebus albigena | 8.3 | 6.0 | Wild | 9 |
| M. mulatta | 11 | 8.8 | Wild | 11 |
| Papio anubis | 25.1 | 13.3 | Wild | 9 |
| P. badius | 8.4 | 8.2 | Wild | 9 |
| S. entellus | 19.2 | 14.8 | Captive | 10 |
| Hominoidea | ||||
| Gorilla gorilla | 170.4 | 71.5 | Captive | 10 |
| Homo sapiens | 72.1 | 62.1 | Captive | 12 |
| Hylobates moloch | 6.6 | 6.3 | Captive | 10 |
| Pan paniscus | 42.7 | 33.7 | Captive | 10 |
| P. troglodytes | 59.7 | 45.8 | Wild | 9 |
| P. troglodytes | 59.7 | 45.8 | Captive | 10 |
| Pongo pygmaeus | 78.5 | 35.8 | Captive | 10 |
aMass estimates are based on Smith and Jungers 1997 (H. sapiens = Danish values), excepting M. griseorufus (Génin 2008)
bSources: (1) Duke Lemur Center; (2) S.M. Karpanty, Ranomafana National Park, Madagascar, samples collected from raptor nests; (3) Beza Mahafaly Special Reserve; Madagascar, (4) L.R. Godfrey; (5) P.C. Wright; (6) Santa Rosa National Park, Costa Rica, (7) El Zota Research Station, Costa Rica, (8) K.E. Glander; (9) M.E. Carter; (10) Department of Anthropology, UC Santa Cruz; (11) O’Regan et al. 2008; (12) O’Connell et al. 2001
Background on variation in ε* values
Variation among proteinaceous tissues
The isotopic values of proteinaceous tissues within an individual could vary because (1) the concentration of different AAs varies among tissues, and (2) the isotopic composition of individual amino acids (AAs) shows considerable variation (19.9‰ average range for C, 24.4‰ average range for N; Fig. 1). This variability relates to isotopic differences among ingested AAs, differences in mammalian biosynthetic pathways for non-essential AAs, and the extent to which a mammal either synthesizes or incorporates a particular AA from its diet. This is a complex subject, but a few patterns have emerged. For carbon, glycine and metabolically-related AAs (serine, cysteine) are often 13C-enriched relative to other non-essential AAs, whereas essential AAs track variation in ingested AAs (Hare et al. 1991; Fogel and Tuross 2003; Jim et al. 2006). For nitrogen, there are a suite of AAs that are 15N-enriched with each trophic step (e.g., glutamate, asparate, alanine, isoleucine, valine, proline) and others that do not enrich (e.g., phenylalanine, lysine, glycine) (McClelland and Montoya 2002; Popp et al. 2007). Muscle (myosin) in humans, and most likely other primates, is dominated by 15N-enriched glutamate and alanine (Bergström et al. 1974). Collagen is mainly composed of 13C-enriched glycine (33%) and 15N-enriched aspartate (5%), glutamate (7%), and proline and hydroxyproline (33%). Primate keratin is dominated by 13C-enriched cysteine (~12–17%) and serine (~10%), and 15N-enriched glutamate (~17%) (Hrdy and Baden 1973; O’Connell et al. 2001).
Fig. 1.

Fractionation between individual amino acids and bulk collagen and tendon (Bos only) for carbon (a) and nitrogen (b) in mammals. The number of individuals for each taxon is provided in parentheses. Carbon data sources: Bos (1) (Hare and Estep 1983), Rattus (C3 = 2, C4 = 2) (Jim et al. 2006), Sus (C3 = 4, C4 = 4) (Hare et al. 1991), archaeological Bison (1), Oidocoleus(1), and Sylvilagus (1) (Fogel and Tuross 2003), and archaeological Procavia (2), Syncerus (2), Ovis (3), Raphicerus (2), Arctocephalus (2), whale (2), and humans (32) (Fogel and Tuross 2003; Corr et al. 2005). Nitrogen data sources: Bos (1) (Hare and Estep 1983), Sus (C3 = 4, C4 = 4) (Hare et al. 1991), archaeological Syncerus (2), Ovis (2), Raphicerus (2), Arctocephalus (2), whale (1), and humans (11) (Styring et al. 2010)
Variation in carbonate-apatite ε* values
The measured mean carbon isotope difference between carbon in carbonate and collagen (ε13*carbonate–collagen) is ~7‰ or greater in wild large-bodied herbivores and ~3‰ in faunivorous animals (Table 1). There are two potential explanations for this difference that are not mutually exclusive. First, it could result from differences in dietary macromolecular composition (i.e., protein, lipid, and carbohydrate), which affect both diet-to-protein and diet-to-carbonate ε* values due to differing δ13C values among macromolecules, and differential routing of macromolecules to particular tissues. Second, it could result from differences in how animals digest plant and animal matter, which only affect diet-to-carbonate ε* values (Hedges 2003).
Apatite carbonate, which likely forms in isotopic equilibrium with blood bicarbonate, reflects carbon in bulk diet (i.e., a proportional mixture of carbon from all assimilated macronutrients) (Ambrose and Norr 1993; Passey et al. 2005). The isotopic composition of consumer proteins reflects that of dietary proteins (Ambrose and Norr 1993; Tieszen and Fagre 1993; Ambrose et al. 1997; Howland et al. 2003; Jim et al. 2004, 2006). Essential AAs must be routed directly from the diet, but depending on dietary protein concentration, non-essential AAs can also be routed into consumer tissues or synthesized using carbon from dietary carbohydrates, lipids and proteins. Theoretically, animals on high protein diets (e.g., faunivores) should route more carbon from dietary protein to tissue protein, whereas animals on low protein diets (e.g., many herbivores) should synthesize more non-essential amino acids de novo, incorporating carbon from lipid and carbohydrate as well as protein into their tissue protein (Fogel and Tuross 2003; Hedges 2003; Martínez del Rio and Wolf 2005). Additionally, because assimilation of 13C-depleted lipids could lower apatite δ13C values without affecting body protein δ13C values (due to routing), faunivores with fat-rich diets (such as seals) should have even smaller ε13*carbonate–collagen values (Krueger and Sullivan 1984; Lee-Thorp et al. 1989; Hedges 2003). Provided that these animals consume monoisotopic diets (e.g., only C3-derived foods), this should result in larger and smaller ε13*carbonate–collagen values in herbivores and carnivores, respectively. Whereas, all primates consume a dominantly vegetarian diet (Milton 1987), some genera such as Cebus, Daubentonia, Galago, and Microcebus can consume considerable amounts of animal matter (Milton and May 1976). Based on these dietary differences, we might anticipate that these taxa should have lower ε13*carbonate–collagen values than more herbivorous species. Importantly, controlled diet studies demonstrate that animals fed a mixture of C3, C4 and marine-derived macronutrients exhibit substantial variation in ε13*carbonate–collagen values (Table 1). Mixed diets are unlikely in the majority of wild primate species. However, this could be important for captive primates if they consume manufactured pellets containing a mix of C3 and C4 foods.
The isotopic composition of carbonate in bone apatite is also predicted to vary with the extent to which complex carbohydrates are fermented in the gut (Hedges 2003). During fermentation, bacteria break down structural carbohydrates, releasing appreciable amounts of hydrogen, CO2, and volatile fatty acids (VFA) (Jensen 1996). Some of the released CO2 can be reduced to form CH4. This process discriminates heavily against 13C, leaving the remaining CO2 13C enriched (Metges et al. 1990; Schulze et al. 1997). If even a small amount of this 13C-enriched CO2 enters the blood bicarbonate pool, it could increase the δ13C value of apatite carbonate which forms from this pool, thus increasing ε13*carbonate–diet and ε13*carbonate–collagen values (Passey et al. 2005). The δ13C value of collagen is not affected by methane production (e.g., Metges et al. 1990).
Ruminants have been shown to produce copious amounts of methane and large Δcarbonate–collagen values (e.g., Crutzen et al. 1986; Metges et al. 1990; Table 1). Although some large, non-ruminant herbivores such as camelids and horses also exhibit high levels of methane production and elevated Δcarbonate–collagen values (Crutzen et al. 1986; Langer 1987; Table 1), methane production in most simple-stomached species is trivial, despite the presence of methanogenic bacteria (Crutzen et al. 1986; Jensen 1996). Acidic conditions in the stomachs and small intestines of simple-stomached animals may prevent methane production, but neutral conditions in the posterior portions of the colon may be more amenable (Jensen 1996). Nevertheless, because gases formed near the end of the gastro-intestinal tract do not likely have time to diffuse into the blood stream, 13C-depleted methane produced in the posterior portions of the colon have a negligible effect on apatite δ13C values. Little is known about methane production in nonhuman primates. For the most part, it is doubtful that nonhuman primates would differ substantially from other simple-stomached animals. However, colobine monkeys could provide a possible exception. This subfamily of Old World Primates, has been likened to ruminants because they have large sacculated stomachs to facilitate microbial fermentation of leaves (Kay and Davies 1994). Primates with adaptations for caeco-colic fermentation, such as Alouatta palliata (Lambert 1998), may also have increased levels of methane production. This possibility is strengthened by the observation that horses, which are also caeco-colic fermenters, have Δcarbonate–collagen values (Sullivan and Krueger 1981; Kellner and Schoeninger 2007, Table 1).
Isotopic terminology
Isotope ratios are typically presented using δ notation, where
 |
1 |
and R is the heavy-to-light isotope ratio in element X. It is expressed in parts per thousand (i.e., per mil, ‰). Carbon isotope values are reported relative to the V-PDB standard (a marine carbonate); nitrogen isotope values are relative to AIR. The offset, or fractionation, between two substances (a and b) is often expressed using Δ notation (Martínez del Rio et al. 2009), where
 |
2 |
δ values are trivial to calculate and accurate so long as the differences in δ values among tissues are small. However, Δ values become less accurate as the differences in δ values among tissues increase. We choose to use alternative expressions, the fractionation factor (α) and isotope enrichment values (ε), which provide exact solutions and are not limited by the isotopic scale on which they are calculated (e.g., PDB vs. SMOW). Δ and ε values are nearly identical when isotopic differences among tissues are <1–2‰, but the two increasingly differ with increasing isotopic differences among tissues. When tissues are ≥10‰, Δ and ε values can differ by as much as 0.5‰ (Cerling and Harris 1999). To calculate ε, we first calculate α.
 |
3 |
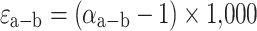 |
4 |
In animals, the observed α value between two tissues, or between diet and a tissue, is the net result of a large range of biochemical and transport phenomena, not the simple equilibrium and kinetic reactions for which isotopic fractionation factors are typically measured. We recognize the complexity of these physiological systems by denoting these as apparent fractionation factors (α*) and apparent enrichment values (ε*). When referring to values for a particular element, we will use ε13* for carbon and ε15* for nitrogen. Note that the sign of enrichment is dependent on which substance is in the numerator in Eq. 3. Hence ε* (and α* and Δ) values must always be reported with subscripts or explicitly defined.
Materials and methods
Sample acquisition
Tissues from captive and wild primates were acquired from cadaveric and osteologic collections in museums, universities and research field stations (Table 2). With a few exceptions, the animals were in good health at the time of death. The main manner of death for captive animals was electrocution, drowning, or short-term illness. However, a few individuals endured chronic illness, and some died at an advanced age. The manner of death for wild animals was largely unknown, but we were able to attribute the deaths of several individuals to predation or automobile impact (Electronic supplementary material, ESM, Table S1). The acquisition and analysis of tissues was approved by the Chancellor’s Animal Research Committee, University of California, Santa Cruz (approval nos. DOMIN 07.01 and ZIHL 97.12), and the Institutional Animal Care and Use Committee, Stony Brook University (approval no. 20001142). We combined our data with data from three preexisting datasets (Kibale primates: Carter, 2001; modern humans: O’Connell et al. 2001; Macaca mulatta: O’Regan et al. 2008).
Sample preparation and analysis
For each specimen, soft tissues were separated and lyophylized. Bone was defleshed; 20 mg were ground for the analysis of carbonate in bone apatite and 50 mg were crushed coarsely for extraction of collagen. For protein analysis, bone samples were treated with 5 ml of 0.5 N HCl for 72 h to remove the mineral fraction. Samples were rinsed 5× with water and dried. Lipids were removed from all proteinaceous tissues by repeated rinsing and sonication in 5 ml aliquots of petroleum ether for 15 min intervals until all visible lipids were removed. Samples were then rinsed 5× with ultrapure water and lyophilized.
With the exception of keratin—which was cleaned, cut to 1 mm lengths, and homogenized—all soft tissue samples were powdered using a mortar and pestle. Approximately 700 μg of ground soft tissue, homogenized keratin, or bone collagen were then sealed into tin boats and analyzed for δ13C and δ15N values on a ThermoElectron (Finnigan) Delta + XP continuous flow system coupled to an elemental analyzer (EA) at the University of California, Santa Cruz (UCSC) Stable Isotope Laboratory. Analytical precision (±1SD) based on 33 replicates of IAEA Acetanilide was −29.6 ± 0.1‰ for carbon and 1.1 ± 0.1‰ for nitrogen. We ran replicate samples for a subset of our specimens to determine sample precision. The average difference between the absolute value of 14 duplicate tissue samples was 0.2 ± 0.2‰ for carbon and 0.2 ± 0.2‰ for nitrogen. The average difference between the absolute value of five triplicate samples was 0.3 ± 0.1‰ and 0.3 ± 0.3‰ for carbon and nitrogen, respectively.
Bone carbonate samples were prepared using a modified technique from Koch et al. (1997). To oxidize organic materials, 1 ml of 30% laboratory-grade hydrogen peroxide (H2O2) was added to 20 mg of powdered sample and left for 48 h, then rinsed 5× with ultrapure water. To remove non-lattice bound carbonate, samples were reacted for 24 h with 0.5 ml of 1 M acetic acid (buffered to pH 5.0 with calcium acetate). Samples were again rinsed 5× with ultrapure water and lyophylized. For carbonate samples, 1.5 mg of powdered bone were put into steel cups and dried at 65°C for 1 h under vacuum. The samples were then analyzed on a Micromass Optima gas source mass spectrometer integrated with an Isocarb automated carbonate device. Samples were dissolved in 100% H3PO4 at 90°C, with concurrent cryogenic distillation of CO2 and H2O and automated CO2 admittance to the mass spectrometer for analysis. Reaction time was set at 740 s and blanks were run between samples. Accuracy and precision (±1SD) based on the international NBS 19 standard analyzed with samples was δ13C = 2.1 ± 0.1‰ (n = 18), very close to the known value of 2.0‰. The average difference between the absolute value of 10 duplicate samples was 0.3 ± 0.2‰.
Data analysis
We were not able to assess dietary composition or digestive physiology carefully for primates included in this study. Although it is tempting to divide primates into broad groups such as folivore, frugivore, or trophic omnivore, these dietary categories would likely be inaccurate for four reasons. First, the majority of primates are generalist primary consumers rather than strict folivores or frugivores. For example, the “frugivorous” lemur Varecia variagata can eat substantial amounts of leaves and fungus (A. Baden, personal communication). Conversely, the diet of Piliocolobus badius, a “folivorous” monkey, frequently contains fruit and flowers (Chapman et al. 2002a). Second, all primates have omnivorous tendencies (Fleagle 1999). In particular, many “frugivorous” primate species supplement their predominantly herbivorous diets either intentionally or inadvertently with insects or vertebrates. For example, among the “frugivorous” species, Hylobates lar and Lemur catta spend a substantial amount of time feeding on insects in addition to vegetation (Rowe 1996; Yamashita 2002), and Pan troglodytes consumes termites and red colobus monkeys (Boesch and Boesch-Achermann 2000). Third, primate diets can differ substantially between years and between localities (e.g., Chapman et al.2002a, b; González-Zamora et al. 2009). For example diets ranging from 49 to 87% leaves, and 13–49% fruits have been reported for Mexican populations of A. palliata (Cristóbal-Azkarate and Arroyo-Rodríguez 2007). Finally, we know little about the diets of most of our captive individuals, including the degree to which they were provisioned with chow.
Instead, we used one-way analysis of variance (ANOVA) and Tukey post-hoc tests of honestly significant differences (HSD) to detect differences in ε* values among habitats (e.g., captive, dry, or moist habitat) that may correlate with diet quality. Diet and digestive physiology may covary with two other variables that we were able to assess: body size and phylogenetic relatedness. In general, diet quality decreases with increasing body size (e.g., Kleiber 1961). More folivorous primates have longer and more complex guts than frugivorous or insectivorous primates (Chivers and Hladik 1980), and primates that are more closely related should have more similar digestive physiology. We used Pearson correlation coefficients to determine if ε* values correlate with body mass, and we tested for the potential confounding effects of phylogenetic relatedness by using the primate phylogeny of Bininda-Emonds et al. (2007) and the PDAP module of Mesquite version 2.5 (Maddison and Maddison 2008) to calculate phylogenetic independent contrasts.
Additionally, we used one-way ANOVA and Tukey HSD to detect differences in ε* values among manners of death (grouped into abrupt, short-term illness, long-term illness, and unknown). We used independent sample t tests to detect differences in ε* values between sexes. Detailed age information for strepsirrhines from the Duke Lemur Center allowed us to calculate percent lifespan lived. We grouped these individuals into five equal age classes, and used one-way ANOVA and Tukey HSD to detect differences in ε* values among age classes. Although there are no theoretical expectations for ε* differences among sexes, age classes, or manners of death, we sought to verify that metabolic or dietary differences between these different groups do not affect ε* values. Such comparisons are often missing from tissue fractionation and enrichment studies. Analyses were performed using JMP version 5.0.1a for Macintosh with the significance of all tests set at α ≤ 0.05.
Results
Mean and standard deviations for each species are presented in Table 3, and raw δ13C, δ15N, and ε* values are available in ESM Table S1. Patterns of apparent enrichment varied little within the Strepsirrhini (Fig. 2) and Haplorrhini (Fig. 3). Across primates, the ε13*collagen–keratin, ε13*collagen–muscle, ε13*muscle–keratin and ε13*carbonate–collagen values did not differ (p > 0.05). Whereas, the ε15*collagen–keratin and ε15*muscle–keratin values also did not differ (p > 0.05), ε15*collagen–muscle values did (t = −2.42, df = 18, p = 0.027); however, this result was driven by two Eulemur and Microcebus individuals. The removal of these two individuals resulted in no overall difference among species for ε15*collagen–muscle (p > 0.05).
Table 3.
Mean carbon and nitrogen apparent enrichment (ε*) values ± one standard deviation for all species and genera
| Superfamily and species | Habitat classesa | ε*bcollagen–keratin | ε*collagen–muscle | ε*muscle–keratin | ε*carbonate–collagen | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | Carbon | Nitrogen | n | Carbon | Nitrogen | n | Carbon | Nitrogen | n | Carbon | ||
| Lorisoidea | ||||||||||||
| Galago senegalensis mohili | C | 5 | 2.3 ± 1.0 | 0.6 ± 1.3 | 3 | 2.1 ± 0.2 | 0.5 ± 0.9 | 3 | 0.5 ± 0.9 | 0.8 ± 0.5 | 5 | 4.9 ± 0.5 |
| Lemuroidea | ||||||||||||
| Avahi laniger | M | 1 | 0.0 | −0.4 | 5 | 0.8 ± 1.0 | −0.7 ± 0.7 | 1 | −1.9 | 0.7 | 9 | 5.1 ± 0.9 |
| Cheirogaleus major | M | 4 | 1.5 ± 0.6 | −0.6 ± 1.7 | 4 | 1.4 ± 1.0 | −1.2 ± 1.2 | 2 | 0.1 ± 1.4 | 1.3 ± 1.7 | 9 | 5.5 ± 0.9 |
| Daubentonia madagascariensis | C | 2 | 2.0 ± 0.2 | 0.3 ± 0.5 | 2 | 2.6 ± 0.6 | −0.6 ± 1.6 | 2 | −0.6 ± 0.3 | 0.9 ± 1.2 | 1 | 3.8 |
| Eulemur fulvus albifrons | C | 3 | 1.0 ± 0.9 | 1.0 ± 0.5 | 3 | 0.7 ± 1.4 | 0.5 ± 0.7 | 3 | 0.3 ± 1.5 | 0.5 ± 0.3 | 3 | 6.1 ± 0.8 |
| E. fulvus rufus | M | 1 | 5.8 | |||||||||
| E. macaco flavifrons | C | 2 | 1.9 ± 0.6 | 2.3 ± 1.8 | 2 | 1.6 ± 0.5 | 1.4 ± 0.3 | 2 | 0.2 ± 1.0 | 1.1 ± 1.3 | 2 | 5.9 ± 0.3 |
| E. mongoz | C | 1 | 0.9 | 2.1 | 1 | 6.2 | ||||||
| Eulemur mean | 6 | 1.3 ± 0.8 | 1.6 ± 1.1 | 5 | 1.1 ± 1.1 | 0.8 ± 0.7 | 5 | 0.2 ± 1.2 | 0.8 ± 0.8 | 7 | 6.0 ± 0.5 | |
| Lemur catta | C | 2 | 1.3 ± 0.3 | 1.1 ± 0.0 | 2 | 0.6 ± 1.2 | 0.0 ± 0.1 | 2 | 0.7 ± 1.5 | 0.9 ± 0.3 | 2 | 6.7 ± 0.0 |
| Indri indri | M | 1 | 0.0 | −0.9 | 1 | 4.5 | ||||||
| Microcebus griseorufus | D | 2 | 5.7 ± 1.7 | |||||||||
| M. murinus | C | 3 | 1.6 ± 0.8 | 0.5 ± 0.3 | 3 | 3.0 ± 0.4 | 0.3 ± 0.3 | 3 | −1.0 ± 0.2 | 0.2 ± 0.3 | 3 | 5.4 ± 0.8 |
| M. rufus | M | 1 | 0.1 | 1.4 | 1 | 1 | −0.2 | 1 | −0.9 | 1.5 | 1 | 6 |
| Microcebus mean | 4 | 1.2 ± 1.0 | 0.7 ± 0.5 | 4 | 2.2 ± 1.0 | 0.2 ± 0.4 | 4 | −1.0 ± 0.1 | 0.5 ± 0.7 | 6 | 5.6 ± 0.9 | |
| Propithecus coquereli verreauxi | C | 3 | 0.1 ± 1.2 | 2.0 ± 1.2 | 3 | −0.2 ± 0.6 | 0.6 ± 0.6 | 3 | 0.3 ± 1.7 | 1.4 ± 1.1 | 2 | 6.1 ± 0.2 |
| P. diadema | M | 1 | 1.4 | 0.6 | 1 | 0.2 | −0.9 | 2 | 4.7 ± 0.5 | |||
| P. verreauxi | D | 4 | 1.1 ± 0.6 | 1.0 ± 0.7 | 2 | 0.2 ± 0.4 | 1.1 ± 1.3 | 2 | 1.3 ± 0.0 | 1.3 ± 0.7 | 6 | 4.8 ± 1.0 |
| Propithecus mean | 8 | 0.8 ± 0.9 | 1.3 ± 1.0 | 6 | 0.0 ± 0.4 | 0.5 ± 1.0 | 5 | 0.7 ± 1.3 | 1.4 ± 0.9 | 10 | 5.0 ± 1.0 | |
| Varecia variagata | C | 1 | 1.2 | 1.3 | 1 | −0.6 | 0.7 | 1 | −0.8 | 0.7 | 1 | 4.8 |
| Ceboidea | ||||||||||||
| Alouatta palliata | M | 1 | 1.1 | 0.6 | 4 | 7.6 ± 1.3 | ||||||
| A. palliata | D | 7 | 1.0 ± 0.6 | 0.4 ± 0.4 | 4 | 0.7 ± 0.5 | −0.6 ± 0.5 | 3 | 0.0 ± 0.9 | 1.1 ± 0.7 | 8 | 5.7 ± 0.4 |
| Alouatta mean | 8 | 1.0 ± 0.5 | 0.4 ± 0.4 | 4 | 0.7 ± 0.5 | −0.6 ± 0.5 | 3 | 0.0 ± 0.9 | 1.1 ± 0.7 | 22 | 6.1 ± 1.0 | |
| Ateles geoffroyi | M | 2 | 8.4 ± 0.1 | |||||||||
| A. geoffroyi | D | 2 | 1.3 ± 0.7 | 0.3 ± 0.4 | 2 | 5.3 ± 0.2 | ||||||
| Ateles mean | 2 | 1.3 ± 0.7 | 0.3 ± 0.4 | 4 | 6.8 ± 1.8 | |||||||
| Cebus capucinus | M | 1 | 5 | |||||||||
| C. capucinus | D | 2 | −0.5 ± 0.5 | 1.3 ± 0.2 | 2 | −0.3 ± 0.5 | −0.1 ± 0.4 | 1 | −0.2 | 1.6 | 1 | 5.8 |
| Cebus mean | 2 | −0.5 ± 0.5 | 1.3 ± 0.2 | 2 | −0.3 ± 0.5 | −0.1 ± 0.4 | 1 | −0.2 | 1.6 | 2 | 5.4 ± 0.6 | |
| Cercopithecoidea | ||||||||||||
| Cercopithecus ascanius c | M | 1 | 2 | 1.9 | 4 | 4.3 ± 0.4 | ||||||
| Chlorocebus aethiops | C | 1 | −0.6 | 1.8 | 1 | −0.4 | 1 | 1 | −0.2 | 0.9 | 1 | 5.9 |
| Lophocebus albigena c | M | 1 | 5.9 | |||||||||
| Macaca mulatta d | M | 13 | −0.3 ± 1.2 | 0.4 ± 0.4 | 11 | 5.9 ± 0.7 | ||||||
| Papio anubis c | M | 6 | 5.3 ± 0.8 | |||||||||
| Piliocolobus badius c | M | 3 | 1.2 ± 0.4 | 0.7 ± 0.5 | 15 | 4.6 ± 1.0 | ||||||
| Semnopithecus entellus | C | 3 | 0.8 ± 0.9 | 0.2 ± 0.1 | 3 | 5.1 ± 0.7 | ||||||
| Hominoidea | ||||||||||||
| Gorilla gorilla | C | 5 | 0.5 ± 0.2 | 1.4 ± 0.4 | 3 | 0.8 ± 0.5 | 0.05 ± 1.3 | 4 | 0.7 ± 2.2 | 1.2 ± 1.1 | 5 | 6.2 ± 0.8 |
| Homo sapiens e | C | 8 | 1.4 ± 0.5 | 0.9 ± 0.2 | ||||||||
| Hylobates moloch | C | 2 | 0.6 ± 0.2 | 1.7 ± 0.6 | 1 | 1.9 | 0.9 | 2 | −1.3 ± 0.7 | 0.0 ± 0.6 | 2 | 6.3 ± 0.5 |
| Pan paniscus | C | 2 | 0.6 ± 2.4 | 0.1 ± 0.6 | 1 | 0.4 | −1.7 | 2 | −0.6 ± 1.1 | 0.2 ± 1.6 | 2 | 6.0 ± 0.4 |
| P. troglodytes c | M | 8 | 6.6 ± 1.5 | |||||||||
| P. troglodytes | C | 1 | 6.1 | |||||||||
| P. troglodytes mean | 9 | 6.1 ± 0.7 | ||||||||||
| Pan mean | 2 | 0.6 ± 2.4 | 0.1 ± 0.6 | 1 | 0.4 | −1.7 | 2 | −0.6 ± 1.1 | 0.2 ± 1.6 | 11 | 6.1 ± 0.6 | |
| Pongo pygmaeus | C | 2 | 0.1 ± 1.1 | 1.6 ± 0.6 | 2 | 6.0 ± 0.5 | ||||||
aHabitat classes are moist (M), dry (D), and captive (C). Moist and dry habitats are differentiated based on rainfall where “Moist” habitats receive >1,000 mm annual precipitation
bApparent enrichment values are reported in parts per thousand (‰)
cData from Carter (2001)
dData from O’Regan et al. (2008)
eData from O’Connell et al. (2001)
Fig. 2.

Mean ε*collagen–keratin, ε*collagen–muscle, ε*muscle-keratin, and ε*carbonate-collagen, for carbon (ε13*) and nitrogen (ε15*) ± 1 standard deviation for each strepsirrhine genus. Phylogeny based on Orlando et al. (2008). Illustrations by Stephen D. Nash/Conservation International, used with permission
Fig. 3.

Mean ε*collagen–keratin, ε*collagen–muscle, ε*muscle–keratin, and ε*carbonate-collagen for carbon (ε13*) and nitrogen (ε15*) ± 1 standard deviation for each haplorrhine genus. Phylogeny based on (Groves 2001). Homo sapiens data from O’Connell et al. (2001), Macaca mulatta data from O’Regan et al. (2008), and P. badius, Cercopithecus ascanius, and wild Pan troglodytes data from Carter (2001). Illustrations by Stephen D. Nash/Conservation International, used with permission
We found small but significant variation among habitat types for both carbon and nitrogen ε*collagen–keratin values (carbon: F 2,82 = 3.36, p = 0.040; nitrogen: F 2,81 = 6.73, p = 0.020). Captive animals had significantly larger ε*collagen–keratin values than those from moist habitats (Table 4). Our results for ε15*collagen–muscle values showed a similar pattern (F 2,44 = 7.03, p = 0.0023), but ε13*collagen–muscle values did not differ significantly among habitat types (p > 0.05). Mean ε13*muscle–keratin and ε13*carbonate–collagen values also did not differ significantly among habitats.
Table 4.
Mean carbon and nitrogen apparent enrichment (ε*) values ± one standard deviation for primates living in dry, moist, and captive settings
| n | ε*acollagen–keratin | n | ε*collagen–muscle | n | ε*muscle-keratin | n | ε*carbonate- collagen | |
|---|---|---|---|---|---|---|---|---|
| Carbon | ||||||||
| Dry | 15 | 0.8 ± 0.8 AB | 9 | 0.3 ± 0.6 A | 6 | 0.4 ± 0.9 A | 29 | 5.5 ± 0.8 A |
| Moist | 25 | 0.4 ± 1.2 B | 12 | 0.9 ± 0.9 A | 4 | −0.7 ± 1.2 A | 75 | 5.5 ± 1.2 A |
| Captive | 43 | 1.1 ± 1.0 A | 25 | 1.3 ± 1.2 A | 30 | −0.05 ± 1.3 A | 35 | 5.7 ± 0.8 A |
| Nitrogen | ||||||||
| Dry | 15 | 0.7 ± 0.6 AB | 8 | −0.1 ± 1.0 AB | 6 | 1.3 ± 0.6 A | n.a. | |
| Moist | 24 | 0.3 ± 0.9 B | 12 | −0.8 ± 0.8 B | 4 | 1.2 ± 1.1 A | n.a. | |
| Captive | 43 | 1.1 ± 0.9 A | 25 | 0.3 ± 0.9 A | 30 | 0.8 ± 0.8 A | n.a. | |
n.a. Not applicable
aApparent enrichment values are reported in parts per thousand (‰). Mean ε* values in the same homogenous subset are given the same letters (α set at 0.05)
With the exception of ε13*collagen–muscle, ε* values between proteinaceous tissues did not correlate with body size (p > 0.05; Table 5). If we excluded two captive Microcebus individuals, the relationship between ε13*collagen–muscle and body size was insignificant (r 2 = 0.06, p = 0.10). The relationship between ε13*carbonate–collagen and body mass was significant (r 2 = 0.031, p = 0.038; Fig. 4; Table 5). However, because the slope is near 0 and the r 2 value is low, we suspect that this result is an artifact of sample size. The range in ε13*carbonate–collagen values for the smallest and largest species (Microcebus spp. and Gorilla gorilla) are similar (4.5–6.9 and 5.5–7.1‰, respectively), and the lowest and highest ε13*carbonate–collagen values, 3.6 and 8.6‰, come from two similar-sized species, P. badius and A. palliata (Tables 2 and 3).
Table 5.
Regression results for ε* versus the natural logarithm of body mass
| n | Carbon | p | n | Nitrogen | p | |
|---|---|---|---|---|---|---|
| r 2 | r 2 | |||||
| Collagen–keratin | 83 | 0.046 | 0.051 | 82 | 0.021 | 0.19 |
| Collagen–muscle | 46 | 0.110 | 0.015 | 45 | −0.023 | 0.98 |
| Muscle–keratin | 36 | −0.024 | 0.670 | 36 | −0.019 | 0.55 |
| Carbonate–collagen | 140 | 0.031 | 0.038 | n.a. | n.a. | n.a. |
Fig. 4.

The relationship between the natural log of body mass (kg) and ε13*carbonate-collagen (ε*carbonate-collagen = 5.24 + 0.163*ln body mass, r 2 = 0.031, p = 0.038)
Finally, ε* values did not differ among males and females (p > 0.05; ESM Table S2), manner of death (p > 0.05; ESM Table S3), or age class (p > 0.05; ESM Table S4). Given the overall consistency of our results, we combined data from all individuals and calculated mean primate ε* values between all proteinaceous tissues, and between carbonate and collagen (Table 6).
Table 6.
Suggested ε* values for comparing different primate tissue types
| Tissue comparison | Carbon | Nitrogen |
|---|---|---|
| Mean ± 1 SD (‰) | Mean ± 1 SD (‰) | |
| Collagen–keratin | 0.9 ± 1.1 | 0.8 ± 0.9 |
| Collagen–muscle | 1.0 ± 1.1 | −0.1 ± 1.0 |
| Muscle–keratin | −0.04 ± 1.2 | 0.9 ± 0.8 |
| Carbonate–collagen | 5.6 ± 1.0 | n.a. |
Discussion
Variation in ε* values among proteinaceous tissues
We expected some variation based on differences in amino acid compositions, but that such differences would be small and consistent across taxa. In line with our expectations, we found small (≤1‰) ε* values between collagen and muscle, collagen and keratin, and muscle and keratin for both carbon and nitrogen (Table 6). These mean values are smaller than the majority of the Δ values reported for captive or wild animals (Table 1). It appears that because each tissue is composed of multiple AAs, the effects of isotopic differences among specific AAs are minimized. For example, relatively 13C-enriched glycine in collagen, serine, and cysteine in keratin, and glutamate in muscle may be driving similar δ13C values in all three tissues (Fig. 1). O’Connell et al. (2001) suggest that the relatively elevated levels of serine and threonine in keratin (6–7% vs. ~2% in collagen) tend to lower keratin δ15N values relative to collagen. The 15N-enriched glutamate in muscle may increase its δ15N values relative to keratin.
Variation in ε13*carbonate–collagen
We had anticipated that differences in diet (e.g., δ13C differences in dietary sources, differences in herbivory vs. faunivory), and digestive physiology (e.g., degree of fermentation) would lead to differences in ε13*carbonate–collagen. Our results, however, suggest that all primates have comparable ε13*carbonate–collagen values regardless of variation in the variables that covary with diet and digestive physiology such as phylogeny, body size, and habitat. Our mean ε13*carbonate–collagen value of 5.6‰ for primates is similar to the mean fractionation factor (Δcarbonate–collagen) for wild omnivores (5.5‰), captive omnivorous rodents fed mixed and uniform diets (5.5 and 5.4‰, respectively), and captive pigs fed uniform diets (6.0‰; Table 1). The mean primate ε13*carbonate–collagen value is larger than the Δcarbonate–collagen value reported for carnivores (3.0‰), and smaller than the reported values for both wild ruminant and non-ruminant herbivores (9.0 and 7.8‰, respectively; Table 1).1 Based on the consistency of our results, we conclude that (1) δ13C values for dietary protein did not differ substantially from whole diet δ13C values for either captive or wild primates, and (2) that microbial fermentation, to the extent that it occurred in the primates in our study, failed to significantly label the blood pool with 13C-enriched bicarbonate, irrespective of differences in habitat, gut physiology or body size.
Diet
We had anticipated that more herbivorous primates would have larger ε13*carbonate–collagen values than more faunivorous primates. We found some variation but no consistent trends. We found no differences in ε13*carbonate–collagen values with body size, despite probable dietary differences between the smallest primates, Galago and Microcebus spp., which likely consumed more insect matter, and the largest primates, Gorilla, Pan, and Pongo, which likely consumed more vegetation. A single aye-aye (Daubentonia madagascariensis), which relies largely on invertebrate prey, had a carnivore-like ε13*carbonate–collagen value of 3.7‰. However, ε13*carbonate–collagen values for the white-faced capuchin (Cebus capucinus), which also consumes animal matter, resembled the overall primate mean (5.0 and 5.8‰ in dry and moist habitat, respectively). Our results likely reflect underlying dietary similarities among all primate species. In spite of apparent differences in the consumption of animal matter, all primates have a predominantly vegetarian diet (Milton 1987). These results agree with the recent findings of Smith et al. (2010), who showed that collagen δ13C values did not differ between male and female chimpanzees despite observations that males consumed substantially greater amounts of red colobus meat. These authors speculated that either meat consumption did not noticeably affect male collagen δ13C values, or that consumption of termites elevated female δ13C values (Smith et al. 2010).
With the exception of a few M. mulatta individuals (O’Regan et al. 2008; ESM Table S1), none of the wild primates in our study ate C4 or marine foods (mean δ13C apatite = −16.7‰ ± 1.4, n = 105; mean δ13C collagen = −22.1‰ ± 1.5, n = 110). Conversely, the majority of our captive primates appear to have incorporated some C4 foods into their diets (mean δ13C apatite = −12.7‰ ± 1.6, n = 36; mean δ13C collagen = −18.5‰ ± 1.8, n = 45; ESM Table S1). However, despite this addition of C4 foods, ε13*carbonate–collagen values for captive and wild primates do not differ (Table 4). We cannot assess dietary composition in the captive primates quantitatively. Nevertheless, our results suggest that protein and whole diet δ13C values did not differ substantially for captive animals. We note that the laboratory diets for some of the controlled feeding studies listed in Table 1 were designed to maximize possible isotopic differences among tissues. The majority of these diets were not designed to maintain healthy individuals, and most laboratory animals were sacrificed at a young age. In contrast, captive primates are given balanced diets designed to maintain their health and increase their longevity. As a result, diets for captive primates tend to be much more isotopically restricted than experimental laboratory diets consisting of mixed C3, C4, and marine components. In line with this reasoning, the range in captive primate ε13*carbonate–collagen values (3.8–7.1‰) is similar to the range in Δcarbonate–collagen values reported for captive animals fed isotopically homogenous diets (4.5–6.7‰), but much smaller than the ranges reported for captive animals fed experimental diets incorporating a mix of C3, C4, and marine components (−0.8 to 11.3‰, Table 1).
Physiology and fermentation
Despite differences in diet, all primates ferment their food to some degree. More folivorous, gumnivorous, and faunivorous primates break down the structural carbohydrates in vegetation, plant exudates, and arthropod exoskeletons, respectively (Lambert 1998). Nevertheless, carbohydrate fermentation in primates does not appear to produce enough methane and associated 13C-enriched CO2 to significantly label blood bicarbonate or bone carbonate. We might have anticipated that taxa with long measured retention times such as Gorilla, Pongo, Lophocebus, Chlorocebus, and Cercopithecus might have larger ε13*carbonate–collagen values (Kleiber 1961; Langer 1987). Increased retention time may increase methane production during fermentation, and the degree to which 13C-enriched CO2 diffuses into the blood (Kleiber 1961; Langer 1987). Our results do not support these expectations. Mean ε13*carbonate–collagen values for the hominoids (6.2 and 6.0‰, respectively), and the cercopithecines (5.9, 5.9, and 4.3‰, respectively; Table 1) are comparable to or only slightly larger than our mean primate ε13*carbonate–collagen value (5.6‰). Conversely, the ε13*carbonate–collagen value for Ateles geoffroyi, which has a fast retention time (6.8‰), is substantially larger than the average primate value.
We had also anticipated that colobine monkeys, represented by P. badius and Semnopithecus entellus, and the ateline monkey A. palliata, would have higher ε13*carbonate–collagen values associated with fermentation in their enlarged stomachs and caeca, respectively. Our results do not support these expectations. Despite their potential for increased levels of methane production, both wild and captive colobine monkeys in our dataset had ε13*carbonate–collagen values comparable to other primate species (Fig. 2; Tables 3 and ESM S1). Our lowest reported ε13*carbonate–collagen value (3.6‰) is from a wild P. badius individual. This result is in agreement with the lack of methane production observed in two wild Colobus polykomos individuals (Ohwaki et al. 1974). It appears that, despite their large “ruminant-like” stomachs, colobines produce little to no methane and associated 13C-enriched CO2, and their digestion resembles that of small simple-stomached animals rather than ruminants.
We did find a large mean ε13*carbonate–collagen value for the mantled howling monkey (A. palliata) in a rainforest habitat (7.6‰). However, we also found a large ε13*carbonate–collagen value (8.4‰) for rainforest-dwelling black-handed spider monkeys (A. geoffroyi), which does not have a gut designed for extensive fermentation (Chivers and Hladik 1980). Intriguingly, these two species had comparable but lower ε13*carbonate–collagen values similar to our primate mean in a seasonally dry forest habitat (5.7 and 5.3‰, respectively). Although A. palliata and A. geoffroyi are typically categorized as folivorous and frugivorous, respectively, both of these species have been observed to have highly variable diets (Cristóbal-Azkarate and Arroyo-Rodríguez 2007; González-Zamora et al. 2009). It is possible that they shared dietary items in the rainforest habitat that were rich in non-starch polysaccharides (NPS), the breakdown of which has been associated with increased methane production in pigs (Jensen 1996). Alternatively, it is possible that the two species shared a food item with elevated δ13C values, (e.g., a CAM plant) which increased their whole diet δ13C values without affecting their dietary protein. This result is interesting and suggests that future work examining species-specific ε13*carbonate–collagen values with varying diets could be enlightening. Nevertheless, these are the only two taxa that demonstrate substantial differences in apparent enrichment values among habitats. For example, ε13*carbonate–collagen values for C. capucinus from the same two habitats are much more similar (5.8 and 5.0‰ in the moist and dry habitats, respectively). Pan troglodytes exhibits similar ε13*carbonate–collagen values among captive and moist habitats (6.1 and 6.6‰, respectively), and all Microcebus taxa have similar ε13*carbonate–collagen values in all three habitat types (5.4, 6.0, and 5.7‰ in captive, moist, and dry habitats, respectively). Based on the data available, we therefore advocate using our mean primate ε13*carbonate–collagen value (5.6‰) to compare collagen and carbonate δ13C values among primates.
Verification of ε* values
An important outcome of our analyses is the ability to determine mean apparent enrichment values that can be used in existing and future comparisons based on mixed tissues or samples. To validate primate ε* values, we estimated keratin δ13C and δ15N values by applying mean ε* collagen–keratin values to measured collagen δ13C and δ15N values for wild primate populations not included in our apparent enrichment dataset. We then compared these estimated keratin values to measured keratin values from different individuals within the same wild populations (Table 7). Compellingly, the range of estimated keratin isotope values closely matches the measured keratin isotope values.
Table 7.
Comparing estimated keratin δ13C and δ15N values with measured keratin δ13C and δ15N values from wild and captive primate populations
| Isotope | n | Mean collagen ± 1 SD (‰) | Estimated keratin (‰) | Measured mean keratin ± 1 SD (‰) | n | Source | |
|---|---|---|---|---|---|---|---|
| Wild | |||||||
| Cercopithecus ascanius | Carbon | 3 | −21.1 ± 0.3 | −22.0 | −22.7 ± 0.2 | 2 | 1 |
| Lophocebus albigena | Carbon | 1 | −20.8 | −21.7 | −21.7 ± 0.2 | 2 | 1 |
| Pan troglodytes | Carbon | 9 | −21.5 ± 0.7 | −22.4 | −21.8 | 1 | 1 |
| Piliocolobus badius | Carbon | 12 | −21.0 ± 0.5 | −21.9 | −22.5 ± 0.4 | 6 | 1 |
| Propithecus verreauxi | Carbon | 7 | −21.2 ± 0.7 | −22.1 | −23.1 ± 1.0 | 5 | 2 |
| Nitrogen | 7 | 7.3 ± 0.7 | 6.5 | 6.3 ± 1.2 | 5 | 2 | |
| Captive | |||||||
| Pan paniscus | Carbon | 1 | −20.2 | −21.1 | −20.7 ± 1.2 | 3 | 2 |
| Nitrogen | 1 | 8.3 | 7.5 | 8.0 ± 2.1 | 3 | 2 | |
| Semnopithecus entellus | Carbon | 3 | −16.5 ± 0.5 | −17.4 | −17.2 ± 0.7 | 2 | 2 |
Conclusions
We have presented data on the apparent isotopic enrichment in carbon and nitrogen isotopes between collagen and keratin, collagen and muscle, and apatite carbonate and collagen in primates. Primates are an extremely diverse group of animals in terms of diet, body size, and gut morphology, yet ε* values are relatively invariant across the order. We recommend applying our calculated mean ε* values when comparing isotope values from different modern primate tissues. Additionally, using these mean apparent enrichment values will be essential for accurately predicting how the isotopic niches of extinct primates compare with those of modern extant primates.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Acknowledgments
We are grateful to institutions that donated cadaveric tissues to the Department of Anthropology, UC Santa Cruz (Oklahoma Zoo, San Francisco Zoo, Ft. Worth Zoo, Milwaukee Zoo, Humboldt Zoo, Chaffee Zoological Gardens, Santa Anna Zoo, Duke Lemur Center, The Gorilla Foundation, The Gibbon Conservation Center). We thank M.J. Schoeninger for providing raw collagen and carbonate δ13C values for wild African herbivores. We are also grateful to the following individuals for samples and assistance: M.R. Blanco, A.D. Cunningham, K.A. Dingess, P. Dolhinow, K.E. Glander, L.R. Godfrey, W. McCandless, S. Matarazzo, I. Mesen, A. Mootnick, G. Pieraccini, M.A. Ramsier, R.B. Segura, C. Underwood, E.R. Vogel, P.C. Wright, and S. Zehr. We thank two anonymous reviewers for useful comments on earlier versions of this manuscript. The importation and use of animal tissues was approved by the United States Fish and Wildlife Service (CITES permit nos. 06US130146/9 and 007319). Funding was provided by the David and Lucile Packard Foundation. This is DLC publication #1181.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Footnotes
We acknowledge that the observed difference between primates and non-ruminant herbivores may stem entirely from a lack of broad comparative data. Non-ruminant data are derived from equids and hippos, two groups of herbivores with substantial methane production rates (Crutzen et al. 1986), and camelids, which have Δcarbonate–collagen values comparable to ruminants.
Contributor Information
Brooke E. Crowley, Phone: +1-831-4591963, FAX: +1-831-4595900, Email: bcrowley@pmc.ucsc.edu
Melinda L. Carter, Email: mcarter3@siumed.edu
Sarah M. Karpanty, Email: karpanty@vt.edu
Adrienne L. Zihlman, Email: azihlman@ucsc.edu
Paul L. Koch, Email: pkoch@pmc.ucsc.edu
Nathaniel J. Dominy, Email: njdominy@ucsc.edu
References
- Ambrose SH, DeNiro MJ. The isotopic ecology of East African mammals. Oecologia. 1986;69:395–406. doi: 10.1007/BF00377062. [DOI] [PubMed] [Google Scholar]
- Ambrose SH, Norr L. Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. In: Lambert JB, Grupe G, editors. Prehistoric human bone—archaeology at the molecular level. Berlin: Springer; 1993. pp. 1–37. [Google Scholar]
- Ambrose SH, Butler BM, Hanson DB, Hunter-Anderson RL, Krueger HW. Stable isotopic analysis of human diet in the Marianas Archipelago, Western Pacific. Am J Phys Anthropol. 1997;104:343–361. doi: 10.1002/(SICI)1096-8644(199711)104:3<343::AID-AJPA5>3.0.CO;2-W. [DOI] [PubMed] [Google Scholar]
- Bergström J, Furst P, Noree L-O, Vinnars E. Intracellular free amino acid concentration in human muscle tissue. J Appl Physiol. 1974;36:693–697. doi: 10.1152/jappl.1974.36.6.693. [DOI] [PubMed] [Google Scholar]
- Bininda-Emonds ORP, Cardillo M, Jones KE, MacPhee RDE, Beck RMD, Grenyer R, Price SA, Vos RA, Gittleman JL, Purvis A. The delayed rise of present-day mammals. Nature. 2007;446:507–512. doi: 10.1038/nature05634. [DOI] [PubMed] [Google Scholar]
- Boesch C, Boesch-Achermann H. The Chimpanzees of the Taï Forest: behavioral ecology and evolution. Oxford: Oxford University Press; 2000. [Google Scholar]
- Carter M (2001) Sensitivity of stable isotopes (13C, 15N, and 18O) in bone to dietary specialization and niche separation among sympatric primates in Kibale National Park, Uganda. PhD dissertation, University of Chicago, Chicago
- Cerling TE, Harris JM. Carbon isotope fractionation between diet and bioapatite in ungulate mammals and implications for ecological and paleoecological studies. Oecologia. 1999;120:247–363. doi: 10.1007/s004420050868. [DOI] [PubMed] [Google Scholar]
- Chapman CA, Chapman LJ, Gillespie TR. Scale issues in the study of primate foraging: red colobus of Kibale National Park. Am J Phys Anthropol. 2002;117:349–363. doi: 10.1002/ajpa.10053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chapman CA, Chapman LJ, Cords M, Gathua JM, Gautier-Hion A, Lambert JA, Rode K, Tutin CEG, White LJT. Variation in the diets of Cercopithecus species: differences within forests, among forests, and across species. In: Glenn M, Cords M, editors. The Guenons: diversity and adaptation in African monkeys. New York: Kluwer/Plenum; 2002. pp. 325–350. [Google Scholar]
- Chivers DJ, Hladik CM. Morphology of the gastrointestinal tract in primates: comparisons with other mammals in relation to diet. J Morphol. 1980;166:337–386. doi: 10.1002/jmor.1051660306. [DOI] [PubMed] [Google Scholar]
- Codron D, Luyt J, Lee-Thorp JA, Sponheimer M, de Ruiter D, Codron J. Utilization of savanna-based resources by Plio-Pleistocene baboons. S Afr J Sci. 2005;101:245–248. [Google Scholar]
- Codron D, Lee-Thorp JA, Sponheimer M, de Ruiter D, Codron J. Inter- and intrahabitat dietary variability of chacma baboons (Papio ursinus) in South African savannas based on fecal δ13C, δ15N, and %N. Am J Phys Anthropol. 2006;129:204–214. doi: 10.1002/ajpa.20253. [DOI] [PubMed] [Google Scholar]
- Corr LT, Sealy JC, Horton MC, Evershed RP. A novel marine dietary indicator utilizing compound-specific bone collagen amino acid δ13C values of ancient humans. J Archaeol Sci. 2005;32:321–330. doi: 10.1016/j.jas.2004.10.002. [DOI] [Google Scholar]
- Cristóbal-Azkarate J, Arroyo-Rodríguez V. Diet and activity pattern of howler monkeys (Alouatta palliata) in Los Tuxtlas, Mexico: effects of habitat fragmentation and implications for conservation. Am J Primatol. 2007;69:1013–1029. doi: 10.1002/ajp.20420. [DOI] [PubMed] [Google Scholar]
- Crutzen PJ, Aselmann I, Seiler W. Methane production by domestic animals, wild ruminants, and other herbivore fauna and humans. Tellus Series B. 1986;38:271–284. doi: 10.1111/j.1600-0889.1986.tb00193.x. [DOI] [Google Scholar]
- DeNiro MJ, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta. 1978;42:495–506. doi: 10.1016/0016-7037(78)90199-0. [DOI] [Google Scholar]
- DeNiro MJ, Epstein S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochim Cosmochim Acta. 1981;45:341–351. doi: 10.1016/0016-7037(81)90244-1. [DOI] [Google Scholar]
- Fleagle JG. Primate adaptation and evolution. 2. San Diego: Academic; 1999. [Google Scholar]
- Fogel ML, Tuross N. Extending the limits of paleodietary studies of humans with compound specific carbon isotope analysis of amino acids. J Archaeol Sci. 2003;30:535–545. doi: 10.1016/S0305-4403(02)00199-1. [DOI] [Google Scholar]
- Fourie NH, Lee-Thorp JA, Ackermann RR. Biogeochemical and craniometric investigation of dietary ecology, niche separation, and taxonomy of Plio-Pleistocene cercopithecoids from the Makapansgat Limeworks. Am J Phys Anthropol. 2008;135:121–135. doi: 10.1002/ajpa.20713. [DOI] [PubMed] [Google Scholar]
- Fox-Dobbs K, Bump JK, Peterson RO, Fox DL, Koch PL. Carnivore-specific stable isotope variables and variation in the foraging ecology of modern and ancient wolf populations: case studies from Isle Royale, Minnesota, and La Brea. Can J Zool. 2007;85:458–471. doi: 10.1139/Z07-018. [DOI] [Google Scholar]
- Génin F. Life in unpredictable environments: first investigation of the natural history of Microcebus grseorufus. Int J Primatol. 2008;29:303–321. doi: 10.1007/s10764-008-9243-z. [DOI] [Google Scholar]
- González-Zamora A, Arroyo-Rodríguez V, Chaves OM, Sánchez-Lopez S, Stoner KE, Riba-Hernández P. Diet of spider monkeys (Ateles geoffroyi) in Mesoamerica: current knowledge and future directions. Am J Primatol. 2009;71:8–20. doi: 10.1002/ajp.20625. [DOI] [PubMed] [Google Scholar]
- Groves C. Primate taxonomy. Washington DC: Smithsonian Institution Press; 2001. [Google Scholar]
- Hare PE, Estep MLF. Carbon and nitrogen isotopic composition of amino acids in modern and fossil collagens. Year B Carnegie Inst Wash. 1983;82:410–414. [Google Scholar]
- Hare PE, Fogel ML, Stafford TW, Mitchell AD, Hoering TC. The isotopic composition of carbon and nitrogen in individual amino acids isolated from modern and fossil proteins. J Archaeol Sci. 1991;18:277–292. doi: 10.1016/0305-4403(91)90066-X. [DOI] [Google Scholar]
- Hedges REM. On bone collagen-apatite-carbonate isotopic relationships. Int J Osteoarchaeol. 2003;13:66–79. doi: 10.1002/oa.660. [DOI] [Google Scholar]
- Howland MR, Corr LT, Young SMM, Jones V, Jim S, van der Merwe NJ, Mitchell AD, Evershed RP. Expression of the dietary isotope signal in the compound-specific δ13C values of pig bone lipids and amino acids. Int J Osteoarchaeol. 2003;13:54–65. doi: 10.1002/oa.658. [DOI] [Google Scholar]
- Hrdy D, Baden HP. Biochemical variation of hair keratins in man and non-human primates. Am J Phys Anthropol. 1973;39:19–24. doi: 10.1002/ajpa.1330390104. [DOI] [PubMed] [Google Scholar]
- Jensen BB. Methanogenesis in monogastric animals. Environ Monit Assess. 1996;42:99–112. doi: 10.1007/BF00394044. [DOI] [PubMed] [Google Scholar]
- Jim S, Ambrose SH, Evershed RP. Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: implications for their use in palaeodietary reconstruction. Geochim Cosmochim Acta. 2004;68:61–72. doi: 10.1016/S0016-7037(03)00216-3. [DOI] [Google Scholar]
- Jim S, Jones V, Ambrose SH, Evershed RP. Quantifying dietary macronutrient sources of carbon for bone collagen biosynthesis using natural abundance stable carbon isotope analysis. Br J Nutr. 2006;95:1055–1062. doi: 10.1079/BJN20051685. [DOI] [PubMed] [Google Scholar]
- Kay RNB, Davies AG. Digestive physiology. In: Davies AG, Oates JF, editors. Colobine monkeys: their ecology, behavior and evolution. Cambridge: Cambridge University Press; 1994. pp. 229–248. [Google Scholar]
- Kellner CM, Schoeninger M. A simple carbon isotope model for reconstructing prehistoric human diet. Am J Phys Anthropol. 2007;133:1112–1127. doi: 10.1002/ajpa.20618. [DOI] [PubMed] [Google Scholar]
- Kleiber M. The fire of life: an introduction to animal energetics. New York: Wiley; 1961. [Google Scholar]
- Koch PL, Tuross N, Fogel ML. The effects of sample treatment and diagenesis on the isotopic integrity of carbonate in biogenic hydroxylapatite. J Archaeol Sci. 1997;24:417–429. doi: 10.1006/jasc.1996.0126. [DOI] [Google Scholar]
- Krueger HW, Sullivan CH. Models for carbon isotope fractionation between diet and bone. In: Turnland JF, Johnson PE, editors. Stable isotopes in nutrition. Washington: American Chemical Society; 1984. pp. 205–222. [Google Scholar]
- Lambert JE. Primate digestion: interactions among anatomy, physiology, and feeding ecology. Evol Anthropol. 1998;7:8–20. doi: 10.1002/(SICI)1520-6505(1998)7:1<8::AID-EVAN3>3.0.CO;2-C. [DOI] [Google Scholar]
- Langer P. Evolutionary patterns of Perissodactyla and Artiodactyla (Mammalia) with different types of digestion. Z Syst Evo1ut –forsch. 1987;25:212–236. doi: 10.1111/j.1439-0469.1987.tb00605.x. [DOI] [Google Scholar]
- Lee-Thorp JA, Sealy JC, van der Merwe NJ. Stable carbon isotope ratio differences between bone collagen and bone apatite, and their relationship to diet. J Archaeol Sci. 1989;16:585–599. doi: 10.1016/0305-4403(89)90024-1. [DOI] [Google Scholar]
- Maddison WP, Maddison DR (2008) A modular system for evolutionary analysis, version 2.5. http://www.mesquiteproject.org. Accessed 05 May 2009
- Martínez del Rio C, Wolf BO. Mass-balance models for animal ecology. In: Stark JM, Wang T, editors. Physiological and ecological adaptations to feeding in vertebrates. Enfield: Science; 2005. pp. 141–174. [Google Scholar]
- Martínez del Rio C, Wolf N, Carleton SA, Gannes LZ. Isotopic ecology ten years after a call for more laboratory experiments. Biol Rev Camb Philos Soc. 2009;84:91–111. doi: 10.1111/j.1469-185X.2008.00064.x. [DOI] [PubMed] [Google Scholar]
- McClelland JW, Montoya JP. Trophic relationships and the nitrogen isotopic composition of amino acids in plankton. Ecology. 2002;83:2173–2180. doi: 10.1890/0012-9658(2002)083[2173:TRATNI]2.0.CO;2. [DOI] [Google Scholar]
- Metges C, Kempe K, Schmidt H-L. Dependence of the carbon-isotope contents of breath, carbon-dioxide, milk, serum and rumen fermentation products on the δ13C value of food in dairy-cows. Br J Nutr. 1990;63:187–196. doi: 10.1079/BJN19900106. [DOI] [PubMed] [Google Scholar]
- Milton K. Primate diets and gut morphology: implications for human evolution. In: Harris M, Ross EB, editors. Food and evolution: toward a theory of human food habits. Philadelphia: Temple University Press; 1987. pp. 93–116. [Google Scholar]
- Milton K, May ML. Body weight, diet and home range area in primates. Nature. 1976;259:459–462. doi: 10.1038/259459a0. [DOI] [PubMed] [Google Scholar]
- Nardoto GB, de Godoy PB, de Barros Ferraz ES, Ometto JPHB, Martinelli LA. Stable carbon and nitrogen isotopic fractionation between diet and swine tissues. Sci Agric. 2006;63:579–582. doi: 10.1590/S0103-90162006000600012. [DOI] [Google Scholar]
- Nelson BK, DeNiro MJ, Schoeninger MJ, De Paolo DJ, Hare PE. Effects of diagenesis on strontium, carbon, nitrogen and oxygen concentration and isotopic composition of bone. Geochim Cosmochim Acta. 1986;50:1941–1949. doi: 10.1016/0016-7037(86)90250-4. [DOI] [Google Scholar]
- O’Connell TC, Hedges REM, Healey MA, Simpson AHRW. Isotopic comparison of hair, nail and bone: modern analyses. J Archaeol Sci. 2001;28:1247–1255. doi: 10.1006/jasc.2001.0698. [DOI] [Google Scholar]
- O’Regan HJ, Chenery C, Lamb AL, Stevens RE, Rook L, Elton S. Modern macaque dietary heterogeneity assessed using stable isotope analysis of hair and bone. J Hum Evol. 2008;44:617–626. doi: 10.1016/j.jhevol.2008.05.001. [DOI] [PubMed] [Google Scholar]
- Ohwaki K, Hungate R, Lotter L, Homann R, Maloiy G. Stomach fermentation in East African colobus monkeys in their natural state. Appl Microbiol. 1974;27:713–723. doi: 10.1128/am.27.4.713-723.1974. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Orlando L, Calvignac S, Schnebelen C, Douady CJ, Godfrey LR, Hänni C. DNA from extinct giant lemurs links archaeolemurids to extant indriids. BMC Evol Biol. 2008;8:121–129. doi: 10.1186/1471-2148-8-121. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Passey BH, Robinoson TF, Ayliffe LK, Cerling TE, Sponheimer M, Dearing MD, Roeder BL, Ehleringer JR. Carbon isotope fractionation between diet, breath CO2, and bioapatite in different mammals. J Archaeol Sci. 2005;32:1459–1470. doi: 10.1016/j.jas.2005.03.015. [DOI] [Google Scholar]
- Popp BN, Graham BS, Olson RJ, Hannides CCS, Lott MJ, López-Ibarra GA, Galván-Magaña, Fry B (2007) Insight into the trophic ecology of yellowfin tuna, Thunnus albacares, from compound-specific nitrogen isotope analysis of proteinaceous amino acids. In: Dawson T, Siegwolf R (eds) Stable isotopes as indicators of ecological change. Elsevier/Academic, San Diego, pp 173–190
- Roth JD, Hobson KA (2000) Stable carbon and nitrogen isotopic fractionation between diet and tissue of captive red fox: implications for dietary reconstruction. Can J Zool 78:848–852
- Rowe N. The pictorial guide to living primates. Patagonias: East Hampton; 1996. [Google Scholar]
- Schoeninger MJ (2010) Toward a δ13C isoscape for primates. In: West JB, Bowen GJ, Dawson TE, Tu KP (eds) Isoscapes: understanding movement, pattern, and process on Earth through isotope mapping. Springer, New York, pp 319–333
- Schoeninger MJ, DeNiro MJ. Carbon isotope ratios of apatite from fossil bone cannot be used to reconstruct diets of animals. Nature. 1982;297:577–578. doi: 10.1038/297577a0. [DOI] [PubMed] [Google Scholar]
- Schoeninger MJ, Iwaniec UT, Glander KE. Stable isotope ratios indicate diet and habitat use in new world monkeys. Am J Phys Anthropol. 1997;103:69–83. doi: 10.1002/(SICI)1096-8644(199705)103:1<69::AID-AJPA5>3.0.CO;2-8. [DOI] [PubMed] [Google Scholar]
- Schulze E, Lohmeyer S, Giese W. Determination of 13C/12C-ratios in rumen produced methane and CO2 of cows, sheep and camels. Isotopes Environ Health Stud. 1997;33:75–79. doi: 10.1080/10256019708036334. [DOI] [PubMed] [Google Scholar]
- Smith RJ, Jungers WL. Body mass in comparative anatomy. J Hum Evol. 1997;32:523–559. doi: 10.1006/jhev.1996.0122. [DOI] [PubMed] [Google Scholar]
- Smith CC, Morgan ME, Pilbeam D. Isotopic ecology and dietary profiles of Liberian chimpanzees. J Hum Evol. 2010;58:43–55. doi: 10.1016/j.jhevol.2009.08.001. [DOI] [PubMed] [Google Scholar]
- Sponheimer M, Loudon JE, Codron D, Howells ME, Pruetz JD, Codron J, de Ruiter DJ, Lee-Thorp JA. Do “Savanna” chimpanzees consume C4 resources? J Hum Evol. 2006;51:128–133. doi: 10.1016/j.jhevol.2006.02.002. [DOI] [PubMed] [Google Scholar]
- Styring AK, Sealy JC, Evershed RP. Resolving the builk δ15N values of ancient human and animal bone collagen via compound-specific nitrogen isotope analysis of contituent amino acids. Geochim Cosmochim Acta. 2010;74:241–251. doi: 10.1016/j.gca.2009.09.022. [DOI] [Google Scholar]
- Sullivan CH, Krueger DA. Carbon isotope analysis of separate chemical phases in modern and fossil bone. Nature. 1981;292:333–335. doi: 10.1038/292333a0. [DOI] [PubMed] [Google Scholar]
- Thackeray JF, Henzi SP, Brain CK. Stable carbon and nitrogen isotope analysis of bone collagen in Papiocynocephalus ursinus: comparison with ungulates and Homo sapiens from southern and East African environments. S Afr J Sci. 1996;92:209–212. [Google Scholar]
- Tieszen LL, Fagre T. Effect of diet quality and composition on the isotopic composition of respiratory CO2, bone collagen, bioapatite, and soft tissues. In: Lambert JB, Grupe G, editors. Prehistoric human bone—archaeology at the molecular level. Berlin: Springer; 1993. pp. 121–155. [Google Scholar]
- Tieszen LL, Boutton TW, Tesdahl KG, Slade NA. Fractionation and turnover of stable carbon isotopes in animal tissues: Implications for δ13C analysis of diet. Oecologia. 1983;57:32–37. doi: 10.1007/BF00379558. [DOI] [PubMed] [Google Scholar]
- Vogel JC. Isotopic assessment of the dietary habits of ungulates. S Afr J Sci. 1978;74:298–301. [Google Scholar]
- West JB, Bowen GJ, Cerling TE, Ehleringer JR. Stable isotopes as one of nature’s ecological recorders. Trends Ecol Evol. 2006;21:408–414. doi: 10.1016/j.tree.2006.04.002. [DOI] [PubMed] [Google Scholar]
- Yamashita N (2002) Diets of two lemur species in different microhabitats in Beza Mahafaly special reserve, Madagascar. Int J Primatol 23:1025–1051
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.