


Abstract
DNA double-strand breaks (DSBs) in eukaryotic cells can be repaired by non-homologous end-joining or homologous recombination. The complex containing the Mre11, Rad50 and Nbs1 proteins has been implicated in both DSB repair pathways, even though they are mechanistically different. To get a better understanding of the properties of the human Mre11 (hMre11) protein, we investigated some of its biochemical activities. We found that hMre11 binds both double- and single-stranded (ss)DNA, with a preference for ssDNA. hMre11 does not require DNA ends for efficient binding. Interestingly, hMre11 mediates the annealing of complementary ssDNA molecules. In contrast to the annealing activity of the homologous recombination protein hRad52, the activity of hMre11 is abrogated by the ssDNA binding protein hRPA. We discuss the possible implications of the results for the role(s) of hMre11 in both DSB repair pathways.
INTRODUCTION
DNA double-strand breaks (DSBs) can be required intermediates in specialized aspects of DNA metabolism such as immunoglobin gene rearrangements and meiosis. In contrast to these programmed DSBs, potentially harmful DSBs can be induced by endogenous or exogenous DNA-damaging agents. Unrepaired DSBs can be lethal, whereas misrepaired DSBs can cause chromosomal fragmentation, translocations and deletions. Such lesions are potential inducers of carcinogenesis through activation of proto-oncogenes, inactivation of tumor suppressor genes or loss of heterozygosity. Therefore, effective repair of DSBs is of great importance for the maintenance of genome stability and prevention of carcinogenesis (1).
A number of fundamentally different DSB repair pathways are available in eukaryotic cells, two of which will be discussed further. First, non-homologous end-joining (NHEJ) mediates joining of DSB ends after no or limited processing. DSB repair through NHEJ can result in deletions through the use of short direct repeats on either side of the break, referred to as microhomologies (2–10). Genes involved in NHEJ, most of which were first identified in mutant rodent cell lines, are XRCC4 to XRCC7 (encoding Xrcc4, Ku80, Ku70 and DNA-PKcs proteins, respectively) and LIG4 (encoding DNA ligase IV) (1,11,12). Subsequent analysis of the homologous genes in yeast identified a NHEJ pathway in this organism as well. Interestingly, in addition to the Ku and ligase IV homologs, the RAD50, MRE11 and XRS2 genes were also found to be involved in NHEJ (8,13,14).
In the second repair pathway, homologous recombination, the repair machinery uses a homologous DNA molecule, i.e. the sister chromatid or the homologous chromosome, as a template for accurate repair of both programmed DSBs in meiosis and induced DSBs during mitosis. Homologous recombination requires the products of the RAD52 epistasis group of genes, first identified in the yeast Saccharomyces cerevisiae. This epistasis group includes RAD50, RAD51, RAD52, RAD54, RAD55, RAD57, RAD59, MRE11 and XRS2 (1,15). The RAD50, MRE11 and XRS2 gene products function in the initial steps of meiotic recombination but are also thought to be involved in recombination processes in mitotic cells (16–26).
The Rad50–Mre11–Xrs2 protein complex, like many other DNA repair factors, has been conserved from yeast to mammals (27–33). Disruption of the mammalian RAD50 or MRE11 genes results in inviable cells (34,35). Mutations in the human ortholog of XRS2, the NBS1 gene, can result in Nijmegen breakage syndrome (NBS), a recessive disorder with some phenotypic similarities to ataxia telangiectasia (AT) (31,33,36–38). Furthermore, non-null mutations in human MRE11 (hMRE11) have been linked to ataxia telangiectasia-like disorder (ATLD) (39). Cells from NBS, AT and ATLD patients are hypersensitive to DSB-inducing agents and show radio-resistant DNA synthesis after exposure to ionizing radiation. Recent experiments have shown that ATM, the kinase defective in AT cells, phosphorylates Nbs1 upon the induction of DNA damage (40–43). These observations provide a rationale for the similarities between the NBS, ATLD and AT phenotypes and suggest a link between DNA-damage signaling and the Rad50–Mre11–Nbs1 complex. Except for functions in recombination processes, NHEJ and DNA damage sensing, the complex is thought to have additional functions in telomere maintenance (14,44–49).
Biochemical studies of the yeast and human protein complexes have shown that Mre11 has a 3′ to 5′ Mn2+-dependent exonuclease activity on DNA substrates with blunt or 5′ protruding ends and endonuclease activity on hairpin and single-stranded (ss)DNA substrates (19,47,50,51). The complex containing hRad50 and Nbs1 in addition to the hMre11 protein can also mediate limited DNA duplex unwinding and more efficient hairpin opening, stimulated by ATP. The presence of ATP also allows the complex to endonucleolytically cut a 3′ overhang at a single-strand–double-strand transition (52). In combination with DNA ligase I or IV, hMre11 stimulates the use of microhomologies in an in vitro joining reaction (50). The exonuclease activity of hMre11 is stimulated by mismatched ends, but inhibited if cohesive ends are present and delayed if an internal microhomology is found (53). This suggests that hMre11 might be a nuclease that stimulates the use of microhomology during NHEJ.
Although the details of the biochemical activities of the hMre11 protein are now beginning to emerge, several questions remain about its precise functions in microhomology-dependent NHEJ and homologous recombination. In this study, we further characterize the interactions of hMre11 protein with DNA. In addition to binding of both ssDNA and double-stranded (ds)DNA, the protein can mediate annealing of complementary ssDNA molecules. This annealing activity is both quantitatively and qualitatively different from the annealing activity of another DSB repair protein, hRad52. We discuss a possible common function of the hMre11-containing complex in both microhomology-dependent NHEJ and homologous recombination.
MATERIALS AND METHODS
Protein expression and purification
Human Mre11 protein was produced by infection of Sf21 cells with baculoviruses expressing hMre11 containing a C-terminal 6-histidine (his6) tag (a generous gift of T.Paull and M.Gellert). Sf21 cells were infected at multiplicity of infection (m.o.i.) 10 and harvested after 48 h. The purification protocol was based on a method described previously (50). Briefly, infected cells were collected, washed three times in PBS and frozen in liquid nitrogen. Cells were thawed and resuspended in 5 ml (5 packed cell volumes) buffer A (20 mM Tris–HCl pH 7.9, 500 mM NaCl, 2 mM β-mercaptoethanol) containing 5 mM imidazole and 1 mM Pefablock (Merck). Then, the cells were disrupted by 30 strokes of a type B pestle in a Dounce homogenizer. After 1 h centrifugation at 80 000 g, the soluble fraction was loaded on a 1 ml Ni2+-NTA agarose column (Qiagen), equilibrated in buffer A. The column was washed with 10 vol buffer A, then with 10 vol buffer A containing 40 mM imidazole and subsequently with 10 vol buffer B (20 mM Tris–HCl pH 7.9, 100 mM NaCl, 2 mM β-mercaptoethanol) containing 40 mM imidazole. Bound proteins were eluted in buffer B containing 200 mM imidazole and diluted 1:1 in buffer B containing 2 mM dithiothreitol (DTT). This preparation was loaded on a 1 ml MonoQ column (Pharmacia) equilibrated in buffer B containing 2 mM DTT. After washing the column with 10 column volumes, the proteins were eluted by a 10 ml linear salt gradient from 100 to 500 mM NaCl. Fractions were analyzed by Coomassie-stained sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE). Fractions containing hMre11 were pooled and dialyzed against dialysis buffer (25 mM Tris–HCl pH 8.0, 150 mM KCl, 10% glycerol, 2 mM β-mercaptoethanol and 2 mM DTT). The resulting protein preparation was aliquoted, frozen in liquid nitrogen and stored at –80°C. The nuclease activity of the hMre11 preparation was assayed as described (50).
The human Ku70/80 heterodimer was produced and purified using a protocol similar to the one described above for hMre11. The proteins were produced by co-infection of Sf21 cells with baculoviruses expressing hKu70 containing a C-terminal his6 tag and hKu80 (54). Sf21 cells were infected at m.o.i. 2 and 6 for the hKu70- and hKu80-producing viruses, respectively, to ensure that all recovered hKu70 would be complexed with hKu80. The human RPA heterotrimer was produced by co-expression of its three subunits in Escherichia coli from a polycistronic messenger RNA and purified as described previously (55). Human Rad52 protein containing an N-terminal his6 tag was produced in E.coli and purified as described (56).
DNA substrates
Oligonucleotides used in this study were: MJ19 (GGATGATTATGGTTATACGTGATTAGTAGCGACGAACATTTTGTAGCAGC) and the complementary oligonucleotide MJ20, MJ21 = MJ19 + 3′-TTTT, MJ22 = MJ20 + 3′-TTTT, MJ23 = MJ19 + 5′-TTTT, MJ24 = MJ20 + 5′-TTTT, MJ29 = 5′-end biotinylated MJ19; DG61 (GATCTGGCCTGTCTTACACAGTGGTAGTACTCCACTGTCTGGCTGTACAAAAACCCTCGGG) and the complementary DG62; DG73 (CTAGACCGGACAGCTGGTGTCACGATGTCTGACCTTGTTTTTGGGACGTC) and the complementary DG74. Circular ssDNA used in this study was φX174 virion DNA (NEB). Circular dsDNA was φX174 RF-I DNA. φX174 RF-I DNA was digested with ApaL1 to generate linear dsDNA, which was denatured and subsequently chilled on ice to generate linear ssDNA.
Oligonucleotides were 5′-end labeled with 32P using polynucleotide kinase and purified through a 1 ml G25 Sepharose column.
Generation of anti-hMre11 antibodies
A 1.9 kb XhoI–PstI fragment from the hMRE11 cDNA, obtained by RT–PCR using HeLa cell mRNA, was subcloned into pMal-C2 (NEB). The fusion protein derived from this plasmid contained the maltose binding protein fused to the C-terminal 634 amino acids of hMre11 and was produced in E.coli strain FB810. The protein was present in the insoluble fraction and was dissolved in 6 M urea. It was purified by preparative PAGE, and used to immunize two rabbits. The antibodies were affinity-purified using fusion protein immobilized on a nitrocellulose filter.
DNA-binding reactions
To analyze the DNA-binding properties of hMre11, 32P-5′-end labeled DNA substrate molecules (MJ19 or MJ19 annealed to MJ20), were incubated at 2.5 nM with hMre11 at 50 nM or the indicated concentrations in binding buffer (50 mM HEPES–KOH pH 8.0, 10 mM Tris–HCl, 100 mM KCl, 4.2% glycerol and 0.5 mg/ml BSA) for 15 min at 25°C in a volume of 20 µl. In competition experiments, 5 µl of non-labeled competitor DNA solution was added to 15 µl of initial binding reaction and incubation was continued for 15 min at 25°C. Subsequently, reaction mixtures were analyzed on a 4% native polyacrylamide gel in 0.5× TBE. Gels were analyzed by autoradiography or phosphoimaging. To compare the affinity for different DNA substrates, the remaining percentage of DNA–protein complex was determined and normalized to non-competed reactions. Reciprocal values of these percentages yield a straight line with slope 1 as a function of molar excess for identical competitor DNA. Therefore, the slopes of the curves for different competitor DNA molecules indicate the competitive capacity of the specific competitor.
To recover hMre11 bound to DNA, the 5′-biotinylated MJ29 was incubated for 30 min at room temperature in coupling buffer (5 mM Tris–HCl pH 7.5, 0.5 mM EDTA and 1 M NaCl) with streptavidin-coated magnetic beads (Dynal) that had been prewashed in coupling buffer. The beads were washed three times with coupling buffer, once with non-fat milk and incubated for 2 h with non-fat milk at room temperature. Then, the beads were washed three times with binding buffer as used for standard binding experiments supplemented with 0.1% NP-40. Next, hMre11 was incubated at 24 nM with the DNA coupled to the beads at 20 nM for 30 min in standard binding buffer. Beads were collected and washed three times with binding buffer containing 0.1% NP-40. Bound fractions were analyzed on an 8% SDS–PAGE gel followed by immunoblotting using affinity-purified anti-hMre11 antisera.
Strand annealing reactions
To assay for strand-annealing activities, 32P-5′-end labeled MJ19 was incubated with MJ20 (both at 0.5 nM) and hMre11, hRad52 or hKu70/80 at a concentration of 10 nM. Reactions were done in annealing buffer (50 mM HEPES–KOH pH 8.0, 10 mM KCl, 0.5 mg/ml BSA) for 1 h at 16°C in a volume of 100 µl. To determine the effect of hRPA on the annealing reactions, 32P-5′-end labeled DG61 and unlabeled DG62 were incubated separately at 1.0 nM with 3.4 nM or the indicated concentrations of hRPA for 15 min at 25°C in annealing buffer. The reaction mixtures were then combined and the reaction was continued for 1 h at 16°C after addition of 10 nM or indicated concentrations of hMre11, hRad52 or hKu70/80. Reactions were terminated by phenol extraction and analyzed on native 10% polyacrylamide gels, containing 0.5× TBE. Reaction products were visualized and quantified by autoradiography or phosphoimaging.
RESULTS
DNA binding of hMre11
Recombinant proteins were produced in Sf21 insect cells using a baculovirus expression system or in E.coli. Figure 1 shows a SDS–PAGE analysis of the purified proteins. Titration and time course experiments showed that the specific nuclease activity of the hMre11 preparation was similar to the activity reported previously (50).
Figure 1.
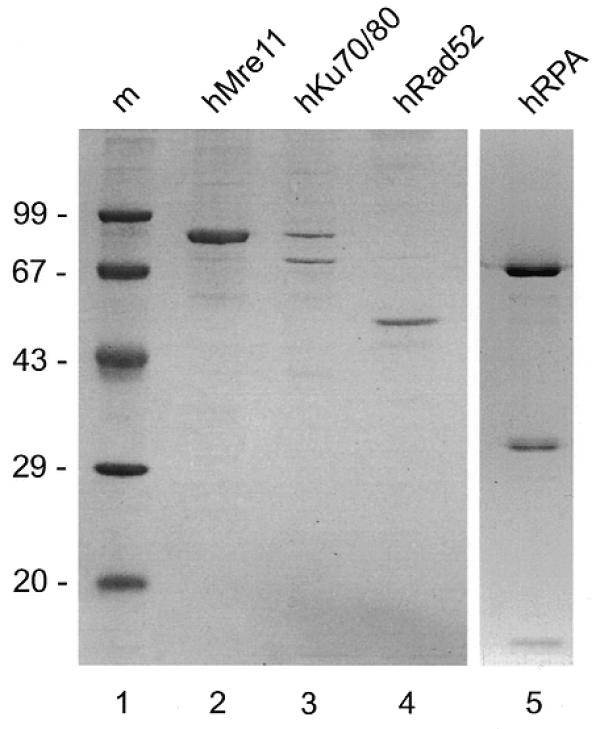
Purified protein preparations. Protein preparations were analyzed by SDS–PAGE and Coomassie blue staining. Lane 1, molecular size marker (m, molecular mass indicated in kDa); lane 2, hMre11-his6 preparation; lane 3, hKu70-his6/hKu80 preparation; lane 4, hRad52-his6 preparation; lane 5, hRPA preparation.
To investigate the complex formation of hMre11 and DNA, the protein was incubated with radiolabeled 50 nt ssDNA and blunt-ended 50 bp dsDNA oligonucleotides without addition of divalent cations. As shown in Figure 2A, hMre11 formed distinct complexes with ssDNA molecules (lanes 1–5) and with dsDNA molecules (lanes 6–10). Incubation with dsDNA resulted in at least two distinct protein–DNA complexes at the higher concentrations of hMre11. Two-dimensional gel analysis of the protein–DNA complexes (native PAGE in the first dimension and SDS–PAGE for separation of ssDNA and dsDNA in the second dimension), showed that both bands in the dsDNA bandshift contain indeed only dsDNA (data not shown) suggesting that these bands represent protein–dsDNA complexes that differ in amount of protein and/or DNA molecules or in conformation. Similar protein–DNA complex formation was also observed with at least eight other ssDNA or dsDNA oligonucleotides that varied in length from 20 to 60 nt/bp and that were unrelated in nucleotide sequence (data not shown). The presence of the divalent cations Mg2+ or Ca2+ did not result in different reaction products (data not shown).
Figure 2.
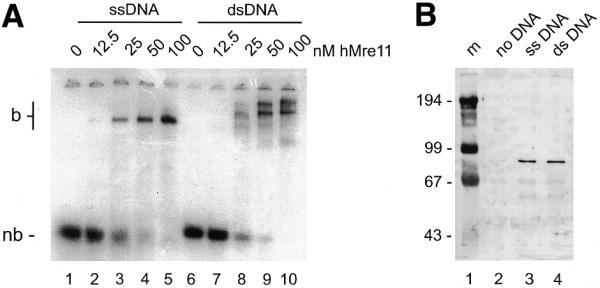
hMre11 binds to ssDNA and dsDNA molecules. (A) Radiolabeled 50 nt ssDNA or blunt-ended dsDNA (MJ19 and MJ19/20, respectively) were incubated at 1.25 nM with the indicated amounts of hMre11 under standard conditions in the absence of divalent cations. Samples were analyzed by native PAGE, followed by autoradiography. nb, non-bound probe; b, bound probe; DNA–protein complex. (B) Biotinylated 50 nt ssDNA or blunt-ended dsDNA (MJ29 and MJ29/20, respectively) were coupled to streptavidin-coated magnetic beads and incubated at 20 nM with 24 nM hMre11 preparation under standard conditions supplemented with 0.1% NP-40. Bound fractions were analyzed by SDS–PAGE and immunoblotting with affinity-purified α-hMre11 antibodies. Lane 1, size marker (m, molecular mass indicated in kDa); lane 2, incubation with control beads lacking DNA; lane 3, incubation with beads coupled to ssDNA; lane 4, incubation with beads coupled to dsDNA.
To confirm that the observed protein–DNA complexes were due to hMre11 binding to both ssDNA and dsDNA molecules, biotinylated ssDNA or blunt-ended dsDNA oligonucleotides were coupled to streptavidin-coated magnetic beads. These beads were incubated with the hMre11 preparation under conditions that resulted in protein–DNA complex formation. Subsequently, the beads were washed with binding buffer and bound proteins were analyzed by SDS–PAGE and immunoblotting. As shown in Figure 2B, incubation of the beads coupled to either ssDNA or dsDNA with the hMre11 protein preparation resulted in recovery of hMre11 (lanes 3 and 4, respectively), whereas no hMre11 protein was recovered in the absence of DNA (lane 2). We conclude that the hMre11 protein binds both ssDNA and dsDNA in a sequence-independent manner.
Characterization of hMre11 DNA-binding properties
The exonuclease activity of hMre11 might suggest a binding preference for DNA ends. We therefore determined whether hMre11 requires free DNA ends to bind its substrate molecule. A competition experiment was performed in which hMre11 was first incubated with a radiolabeled 50 nt ssDNA oligonucleotide under standard assay conditions. Subsequently, non-labeled ssDNA oligonucleotide or circular φX174 ssDNA was added on an equal nucleotide basis and the incubation was continued (Fig. 3A, left). Circular φX174 DNA competed even better than ssDNA oligonucleotides, suggesting that hMre11 might load more efficiently on longer DNA molecules. A similar result was obtained when the complex of hMre11 and dsDNA oligonucleotide was competed with dsDNA oligonucleotide or circular φX174 dsDNA (Fig. 3A, right). No difference was observed between circular and linearized φX174 molecules (data not shown). Therefore, we conclude that hMre11 does not preferentially bind DNA ends.
Figure 3.
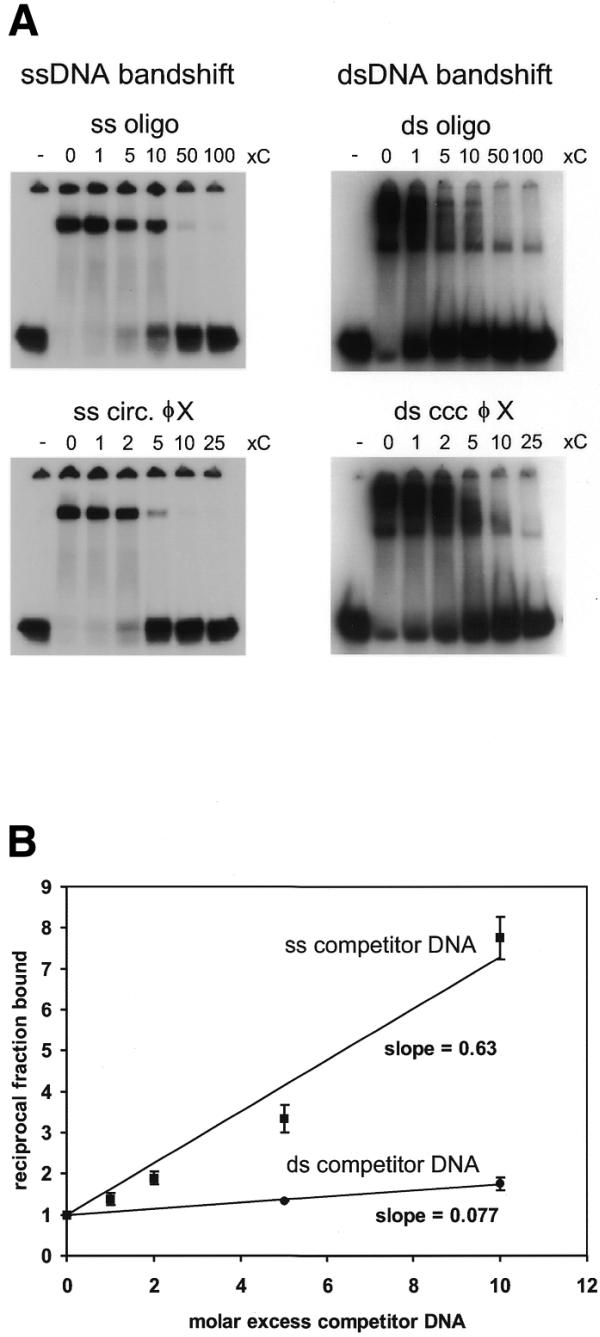
Characterization of hMre11 DNA-binding properties. (A) Radiolabeled 50 nt ssDNA (MJ19; left) or dsDNA (MJ19/MJ20; right) was incubated at 1.25 nM with hMre11 at 50 nM. After 15 min incubation, the indicated nucleotide excess non-radioactively-labeled competitor DNA was added and incubation was continued for an additional 15 min. Competitor DNA was single-stranded oligonucleotide (MJ19) or circular φX174 ssDNA (left) and blunt-ended double-stranded oligonucleotide (MJ19/MJ20) or circular φX174 dsDNA (right). Lane ‘–’, incubation without hMre11. Samples were analyzed by native PAGE, followed by autoradiography. (B) Radiolabeled 50 nt ssDNA (MJ19) was incubated at 1.25 nM with 50 nM hMre11 as in (A). Non-labeled, non-identical ssDNA (DG73) or dsDNA (DG73/DG74) oligonucleotide DNA was used as a competitor. Values on the y-axis are reciprocal fractions of DNA–protein complex, normalized to the fraction of DNA–protein complex in non-competed reactions (Materials and Methods). Bars indicate standard deviations of mean values of triplicate reactions. Slopes of linear regression lines through the datapoints are indicated.
The relative affinity of hMre11 for ssDNA and dsDNA was investigated in competition experiments. After incubation of hMre11 with a radiolabeled 50 nt ssDNA oligonucleotide, non-identical ssDNA or dsDNA competitor oligonucleotide was titrated in and the reaction was continued. The fraction of radiolabeled DNA oligonucleotide bound by hMre11 was determined and normalized to the fraction of probe bound in the absence of competitor DNA. The inverse of this normalized fraction is a linear function of the molar excess of competitor DNA added (Fig. 3B). The slope of the line is indicative of the effectiveness of the competitor. We conclude that hMre11 had an ~9-fold higher binding affinity for ssDNA than for dsDNA molecules. The presence of short (4 nt) 3′ or 5′ ssDNA overhangs did not result in a higher affinity compared to blunt-ended dsDNA (data not shown).
Strand annealing by hMre11
In competition assays with complementary DNA, formation of dsDNA was observed. To investigate this effect in more detail, two complementary 50 nt ssDNA oligonucleotides were incubated with hMre11 without addition of divalent cations. After deproteinization, samples were analyzed by native PAGE. As shown in Figure 4, hMre11 stimulated annealing of the two complementary single strands. As negative and positive controls for these experiments we used the hKu70/80 and hRad52 proteins. The hKu70/80 heterodimer did not support annealing. In contrast, the hRad52 protein displayed a robust annealing activity consistent with previous reports for both the S.cerevisiae and human Rad52 proteins (57–60). The annealing activities for both hMre11 and hRad52 were also observed with other sets of complementary ssDNA oligonucleotides that were unrelated in nucleotide sequence to the oligonucleotide shown in Figure 4 (data not shown).
Figure 4.
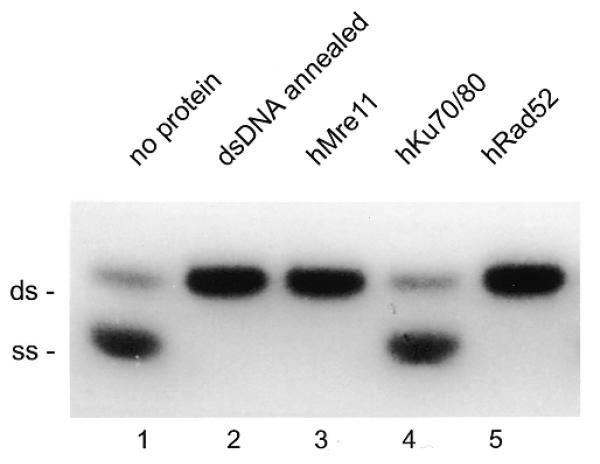
hMre11 anneals complementary ssDNA molecules. Two complementary 50 nt ssDNAs (MJ19 and MJ20), one of which was radiolabeled, were incubated with 10 nM hMre11, hKu70/80, hRad52 or without protein. After deproteinization, samples were analyzed by native PAGE and autoradiography. Lane 1, incubation without protein; lane 2, dsDNA, annealed by heating and slow cooling; lane 3, incubation with hMre11; lane 4, incubation with hKu70/80 heterodimer; lane 5, incubation with hRad52. The positions of ssDNA and dsDNA are indicated.
Kinetics of strand annealing by hMre11 and hRad52
The kinetics of the annealing reaction were determined in a time course experiment in which two complementary 50 nt oligonucleotides were incubated in the presence of hMre11, hRad52 or hKu70/80 or in the absence of protein. The percentage of dsDNA as a function of time was determined (Fig. 5A). Although both hMre11 and hRad52 efficiently annealed complementary DNA molecules, the annealing rate of hRad52 was ~2-fold higher than that of hMre11. To further quantify this difference, the initial reaction rates were determined for different concentrations of hMre11 and hRad52 (Fig. 5B). The maximal annealing rate of hRad52 was approximately twice as high as that of hMre11.
Figure 5.

Kinetics of hMre11-mediated oligonucleotide annealing. (A) Two complementary 50 nt ssDNAs (MJ19 and MJ20), one of which was radiolabeled, were incubated at 0.5 nM with hMre11, hRad52 or hKu70/80, at 1.25 nM, or without protein. Samples were taken at 0, 1, 2, 3, 5, 7.5, 10, 15, 20 and 40 min. Annealing was analyzed as in Figure 4. Bars indicate standard deviation of mean values of triplicate experiments. (B) Annealing reactions as in (A), with the indicated concentrations of either hMre11 or hRad52. Samples were taken at 0, 1 and 2 min. Initial annealing rates, normalized to highest measured annealing rate of hRad52, were determined from the slopes of the time courses. Bars indicate standard deviations of mean values of triplicate reactions.
hMre11-mediated strand annealing is inhibited by hRPA
Most ssDNA in vivo is likely to be covered by ssDNA binding proteins, such as the human RPA heterotrimer. We therefore investigated whether coating the ssDNA oligonucleotides with hRPA would influence strand annealing by hMre11. Complementary ssDNA molecules 61 nt in length were first incubated separately with hRPA. The concentration of hRPA (3.4 nM) was such that all substrate molecules were covered by hRPA, as determined by mobility shift assays (data not shown). After incubation with hRPA, the reaction mixtures containing the complementary oligonucleotides were combined, hMre11 was added at 10 nM and the incubation was continued. As shown in Figure 6A, preincubation of hRPA with the oligonucleotides completely inhibited the strand-annealing activity of hMre11. In contrast, the strand-annealing activity of hRad52 was much less affected. As expected, the presence of hRPA had no effect on the absence of annealing activity by hKu70/80.
Figure 6.
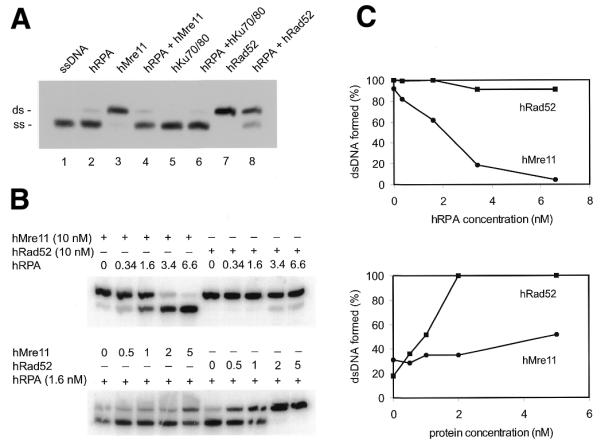
hMre11, but not hRad52-annealing activity is inhibited by precoating of substrate oligonucleotides with hRPA. (A) Complementary 61 nt ssDNAs (DG61 and DG62) were incubated separately at 1 nM with 3.4 nM hRPA for 15 min at 25°C. Reaction mixtures were combined and incubations were continued for 60 min at 16°C after addition of hMre11, hRad52 or hKu70/80 at 10 nM each, or without protein, as indicated. Annealing was analyzed by native PAGE after deproteinization, followed by autoradiography. Lane 1, no proteins added; lane 2, preincubation with hRPA only; lane 3, incubation with hMre11 only; lane 4, preincubation with hRPA, followed by addition of hMre11; lane 5, incubation with hKu70/80 only; lane 6, preincubation with hRPA, followed by addition of hKu70/80; lane 7, incubation with hRad52 only; lane 8, preincubation with hRPA, followed by addition of hRad52. (B) Complementary 61 nt ssDNAs (DG61 and DG62) were incubated separately at 1 nM with the indicated concentrations of hRPA in the top panel or 1.6 nM hRPA in the bottom panel for 15 min at 25°C. Reaction mixtures were combined and incubations were continued after addition of hMre11 or hRad52 at 10 nM in the top panel or at the indicated concentrations in the bottom panel, for 60 min at 16°C. Annealing was analyzed by native PAGE after deproteinization. (C) Quantification of the percentages dsDNA formed in (B). Values are expressed as percentage dsDNA for hMre11 and hRad52.
The effect of precoating the substrate oligonucleotides with hRPA was further analyzed by titration of hRPA, while keeping the concentrations of hMre11 or hRad52 constant (Fig. 6B, top). Alternatively, different concentrations of hMre11 or hRad52 were added after preincubation with a fixed concentration of hRPA (Fig. 6B, bottom). The presence of hRPA at 2 nM reduced hMre11-mediated strand annealing by 50% compared to the absence of hRPA, while hRad52-mediated strand annealing was not affected. Higher concentrations of hRPA only mildly affected hRad52-mediated annealing (Fig. 6C, top). Similarly, preincubation with 1.6 nM hRPA inhibited strand annealing at a low concentration of hMre11, while increasing amounts of hMre11 could only partially overcome the inhibitory effect (50% annealing activity at 5 nM hMre11). In contrast, the hRad52 annealing activity was completely restored at 2 nM (Fig. 6C, bottom).
DISCUSSION
In this paper we describe a number of biochemical properties of the hMre11 protein. We show that hMre11 forms complexes with both ssDNA and dsDNA. The affinity for ssDNA is higher than for dsDNA and hMre11 binds to DNA without the need for free DNA ends. Furthermore, hMre11 can promote annealing of complementary ssDNA molecules, with a slightly slower rate than hRad52. The hMre11 annealing activity is, in contrast to hRad52, inhibited by the presence of hRPA. Although strand-annealing activity has been reported for a number of proteins, the biological significance cannot always be readily assessed. However, strand-annealing activity is a key feature of certain DSB repair pathways. Below we will discuss how this activity of hMre11 might contribute to the repair of DSBs.
The importance of the Mre11 protein in DSB repair is highlighted by a number of observations implicating Mre11 in both major DSB repair pathways in yeast as well as vertebrate cells. In yeast, the involvement of Mre11 in NHEJ and homologous recombination has been revealed by plasmid rejoining assays and genetic experiments, respectively (8,13,14,16–20,22,25,26,32,61). In vertebrate cells a requirement for Mre11 in NHEJ and homologous recombination has been inferred from biochemical experiments and epistasis analysis, respectively (21,50,53). Below we discuss the implications of the DNA binding and complementary ssDNA annealing activities of hMre11 for its potential roles in both NHEJ and homologous recombination.
First we will consider the role of Mre11 in NHEJ. In vivo plasmid rejoining assays have shown that DNA ends are preferentially joined on short direct repeats (i.e. microhomologies) on either side of the DSB in cells from the fission yeast Schizosaccharomyces pombe. Studies with rad32 deletion mutants, the S.pombe homolog of hMRE11, have revealed that this gene is required for microhomology-dependent end-joining (14). In vitro experiments have shown that DNA ligase I or IV can join DNA molecules using microhomologies when combined with the hMre11 protein (50). Furthermore, the hMre11 exonuclease activity is stimulated by mismatched ends, but inhibited by cohesive ends and delayed when an internal microhomology is present (53). The initial step in the microhomology-dependent end-joining pathway could be formation of protruding ssDNA tails to expose the microhomologies. These tails could be created by the 3′ to 5′ exonuclease activity of the Rad50–Mre11–Nbs1 complex (19,47,50,51). Alternatively, the DNA ends could be partially unwound by the Rad50–Mre11–Nbs1 complex and annealed on microhomologies. Subsequently, the displaced ssDNA tails could be removed endonucleolytically (52; Fig. 7). The human ssDNA binding protein RPA will probably not be able to inhibit hMre11 strand-annealing activity, since the microhomology-containing tails are presumably too short for recognition by hRPA (3,6,7,62). The two DNA ends could be held in proximity by the Rad50 protein, which has similarities to the SMC family of proteins. Members of this protein family are involved in processes that require bridging of distant sites on DNA, such as cohesion between chromatids or condensation of DNA (63,64). Further processing of this intermediate in DSB repair could occur by other nucleases, DNA polymerases, ligases and their respective accessory factors.
Figure 7.
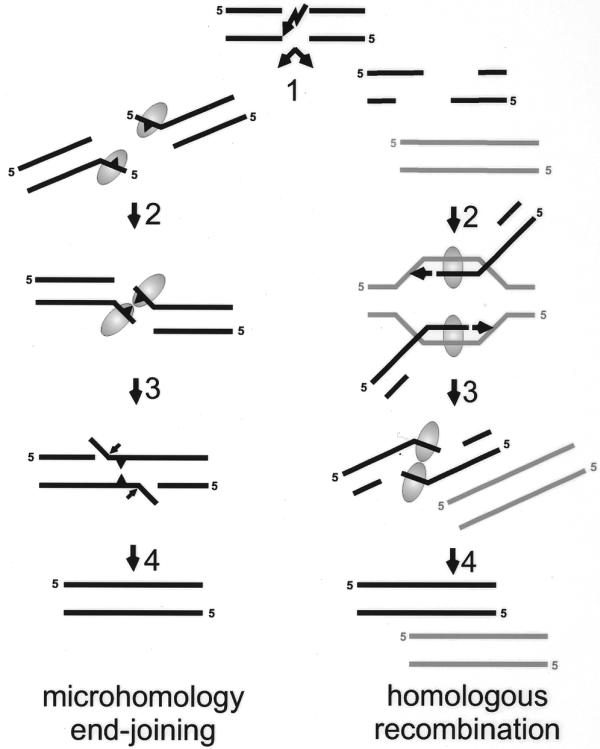
Proposed models for the possible functions of the Mre11 complex in end-joining and homologous recombination. If a DSB is repaired via microhomology-directed end-joining, both ends can be either nucleolytically processed or partially unwound (left, step 1: microhomologies symbolized by triangle). The Mre11-containing complex (symbolized by the oval) may bind the short ssDNA tails. The tails could be kept in close proximity through interaction between two Mre11 protomers either directly or as constituents of the same Rad50/Mre11 complex (left, step 2). The single-strand-annealing activity of Mre11 promotes base pairing between complementary DNA sequences (left, step 3). In the last step the activities of processing enzymes such as flap-endonucleases, polymerases and ligase(s) are required to restore the continuity of the DNA molecule (left, step 4). If the DSB is repaired via homologous recombination, extensive degradation of both ends results in long 3′ ssDNA tails (right, step 1). After strand invasion and homologous pairing (right, step 2) by the Rad52 epistasis group proteins and RPA (not indicated), new DNA is synthesized (arrowhead). Restoration of the lost sequence information will be complete when both newly-synthesized strands overlap (right, step 3). A possible resolution step of this recombination intermediate would involve unwinding of the newly-synthesized strand from the template DNA followed by annealing of the complementary newly-synthesized strands, possibly by Mre11. Finally, single-strand gaps are filled in and ligated, completing the repair of the damaged DNA molecule (right, step 4).
The alternative major DSB repair pathway in eukaryotes is homologous recombination. Genetic experiments with S.cerevisiae clearly implicate ScMre11 in both meiotic and mitotic homologous recombination (16–20,22,25,26,32,61). The protein plays an early role during meiotic recombination because its nuclease activity is required for the processing of meiosis-specific DSBs made by the Spo11 protein (47). Although ScMre11 plays a role in the initiation of meiotic recombination, an early role in mitotic recombination is less likely. An early event in mitotic DSB repair through homologous recombination is the processing of the DSB ends into 3′ ssDNA tails that are required for the invasion of the broken DNA into the duplex homologous repair template DNA. However, its polarity of exonuclease activity make the Mre11 protein a less likely candidate to generate these recombination intermediates (19,50,51). Another possible early role in homologous recombination could be similar to the proposed annealing activity of the Rad52 protein in joint molecule formation (57–60,65–68). However, the ssDNA tail intermediates will most probably be covered by RPA in vivo, which would strongly inhibit the hMre11-mediated annealing activity but not the annealing activity of hRad52. Therefore, a role for hMre11 in this step is less likely, although it is still possible that the presence of the other complex components or additional factors could overcome the inhibitory effect of hRPA.
Instead of an early role in mitotic DSB repair, it is possible that the Mre11 complex functions at a later stage of recombination such as resolution (Fig. 7). If one or both 3′ ssDNA tails invade the intact homologous strands of the sister chromatid or the homologous chromosome, the intermediate structure would consist of an intact chromosome with one or two 3′ ends from which DNA synthesis can start (15). If synthesis proceeds beyond the point of the original break, all lost sequence information has been regained. However, the two ends from the broken chromatid or chromosome have to find each other to restore the continuity of the chromosome. This would require melting of the newly-synthesized strands from their templates and annealing to each other. The unwinding activity detected for the purified human Rad50–Mre11–Nbs1 complex might be useful to initiate the unwinding of the newly-synthesized strands from the template strand (52). Then, hMre11 could be loaded preferentially on the resulting ssDNA and subsequently anneal the complementary newly-synthesized strands. During this reaction the ssDNA could be of limited length, thereby preventing RPA from binding and inhibiting Mre11 annealing activity or the unwinding and annealing could be accomplished in a concerted reaction. The signal for inducing this activity of the Rad50–Mre11–Nbs1 complex could be the proximity of the two DNA polymerase complexes that are moving towards each other.
This potential role for the Mre11 strand-annealing activity, but not necessarily the Mre11 nuclease activity, in resolution of recombination intermediates is consistent with the lack of a recombination and DNA repair-deficient phenotype of nuclease defective mre11 mutants in S.cerevisiae (22). According to the model presented above (Fig. 7), the usual interstrand short-track gene conversion events might not be resolved in mre11Δ cells. Instead, they could result in replication of a large part of the homologous chromosome (or sister chromatid) and therefore an increased rate of recombination between markers on different ends of a chromosome would result. This so-called ‘hyper-recombination’ phenotype has been observed in mitotic mre11Δ and rad50Δ cells (18,69). In addition to the proposed role of the strand-annealing activity of Mre11 in restoring DNA damage induced by exogenous agents, this activity could also be required for the rescue of collapsed replication forks. While Mre11 is dispensable in yeast cells, it is essential for growth of vertebrate cells (21,32,35,70). Because S.cerevisiae chromosomes are in general much shorter than vertebrate chromosomes, S.cerevisiae mre11Δ cells could be viable because the break induced replication can more easily continue to the end of the chromosome in S.cerevisiae cells than in vertebrate cells.
The proposed functions for the hMre11 complex in end-joining and homologous recombination are not mutually exclusive. Most of the activities described in both models are conceptually similar; first to disrupt base pairing at the DSB end in NHEJ or with the template strand in homologous recombination, and subsequently to anneal homologous ssDNA strands which are held in close proximity (Fig. 7). Therefore, these fundamentally different repair processes might both rely on the use of a similar enzymatic activity of the Rad50–Mre11–Nbs1 complex.
Acknowledgments
ACKNOWLEDGEMENTS
We thank T.Paull and M.Gellert for the generous gift of the hMre11-producing baculovirus and J.H.J.Hoeijmakers for carefully reading the manuscript. This work was supported by grants from the Dutch Cancer Society (KWF), the Dutch Organisation for Scientific Research (NWO) and the Human Frontiers Science Project Organization (HFSPO). D.C.v.G. is an academy fellow of the Royal Netherlands Academy for Arts and Sciences (KNAW).
References
- 1.Kanaar R., Hoeijmakers,J.H. and van Gent,D.C. (1998) Molecular mechanisms of DNA double strand break repair. Trends Cell Biol., 8, 483–489. [DOI] [PubMed] [Google Scholar]
- 2.Kramer K.M., Brock,J.A., Bloom,K., Moore,J.K. and Haber,J.E. (1994) Two different types of double-strand breaks in Saccharomyces cerevisiae are repaired by similar RAD52-independent, nonhomologous recombination events. Mol. Cell. Biol., 14, 1293–1301. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Roth D.B. and Wilson,J.H. (1986) Nonhomologous recombination in mammalian cells: role for short sequence homologies in the joining reaction. Mol. Cell. Biol., 6, 4295–4304. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Thode S., Schafer,A., Pfeiffer,P. and Vielmetter,W. (1990) A novel pathway of DNA end-to-end joining. Cell, 60, 921–928. [DOI] [PubMed] [Google Scholar]
- 5.Pfeiffer P., Thode,S., Hancke,J. and Vielmetter,W. (1994) Mechanisms of overlap formation in nonhomologous DNA end joining. Mol. Cell. Biol., 14, 888–895. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Thacker J., Chalk,J., Ganesh,A. and North,P. (1992) A mechanism for deletion formation in DNA by human cell extracts: the involvement of short sequence repeats. Nucleic Acids Res., 20, 6183–6188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Roth D.B., Porter,T.N. and Wilson,J.H. (1985) Mechanisms of nonhomologous recombination in mammalian cells. Mol. Cell. Biol., 5, 2599–2607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Moore J.K. and Haber,J.E. (1996) Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae. Mol. Cell. Biol., 16, 2164–2173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mason R.M., Thacker,J. and Fairman,M.P. (1996) The joining of non-complementary DNA double-strand breaks by mammalian extracts. Nucleic Acids Res., 24, 4946–4953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Ducau J., Bregliano,J. and de La Roche Saint-Andre,C. (2000) Gamma-irradiation stimulates homology-directed DNA double-strand break repair in Drosophila embryo. Mutat. Res., 460, 69–80. [DOI] [PubMed] [Google Scholar]
- 11.Tsukamoto Y. and Ikeda,H. (1998) Double-strand break repair mediated by DNA end-joining. Genes Cell., 3, 135–144. [DOI] [PubMed] [Google Scholar]
- 12.Lewis L.K. and Resnick,M.A. (2000) Tying up loose ends: nonhomologous end-joining in Saccharomyces cerevisiae. Mutat. Res., 451, 71–89. [DOI] [PubMed] [Google Scholar]
- 13.Tsukamoto Y., Kato,J. and Ikeda,H. (1997) Budding yeast Rad50, Mre11, Xrs2 and Hdf1, but not Rad52, are involved in the formation of deletions on a dicentric plasmid. Mol. Gen. Genet., 255, 543–547. [DOI] [PubMed] [Google Scholar]
- 14.Wilson S., Warr,N., Taylor,D.L. and Watts,F.Z. (1999) The role of Schizosaccharomyces pombe Rad32, the Mre11 homologue and other DNA damage response proteins in non-homologous end joining and telomere length maintenance. Nucleic Acids Res., 27, 2655–2661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Paques F. and Haber,J.E. (1999) Multiple pathways of recombination induced by double-strand breaks in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev., 63, 349–404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tsubouchi H. and Ogawa,H. (1998) A novel mre11 mutation impairs processing of double-strand breaks of DNA during both mitosis and meiosis. Mol. Cell. Biol., 18, 260–268. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ivanov E.L., Sugawara,N., White,C.I., Fabre,F. and Haber,J.E. (1994) Mutations in XRS2 and RAD50 delay but do not prevent mating-type switching in Saccharomyces cerevisiae. Mol. Cell. Biol., 14, 3414–3425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Bressan D.A., Olivares,H.A., Nelms,B.E. and Petrini,J.H. (1998) Alteration of N-terminal phosphoesterase signature motifs inactivates Saccharomyces cerevisiae Mre11. Genetics, 150, 591–600. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Furuse M., Nagase,Y., Tsubouchi,H., Murakami-Murofushi,K., Shibata,T. and Ohta,K. (1998) Distinct roles of two separable in vitro activities of yeast Mre11 in mitotic and meiotic recombination. EMBO J., 17, 6412–6425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Usui T., Ohta,T., Oshiumi,H., Tomizawa,J., Ogawa,H. and Ogawa,T. (1998) Complex formation and functional versatility of Mre11 of budding yeast in recombination. Cell, 95, 705–716. [DOI] [PubMed] [Google Scholar]
- 21.Yamaguchi-Iwai Y., Sonoda,E., Sasaki,M.S., Morrison,C., Haraguchi,T., Hiraoka,Y., Yamashita,Y.M., Yagi,T., Takata,M., Price,C., Kakazu,N. and Takeda,S. (1999) Mre11 is essential for the maintenance of chromosomal DNA in vertebrate cells. EMBO J., 18, 6619–6629. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Bressan D.A., Baxter,B.K. and Petrini,J.H. (1999) The Mre11–Rad50–Xrs2 protein complex facilitates homologous recombination-based double-strand break repair in Saccharomyces cerevisiae. Mol. Cell. Biol., 19, 7681–7687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Sugawara N. and Haber,J.E. (1992) Characterization of double-strand break-induced recombination: homology requirements and single-stranded DNA formation. Mol. Cell. Biol., 12, 563–575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Alani E., Padmore,R. and Kleckner,N. (1990) Analysis of wild-type and rad50 mutants of yeast suggests an intimate relationship between meiotic chromosome synapsis and recombination. Cell, 61, 419–436. [DOI] [PubMed] [Google Scholar]
- 25.Johzuka K. and Ogawa,H. (1995) Interaction of Mre11 and Rad50: two proteins required for DNA repair and meiosis-specific double-strand break formation in Saccharomyces cerevisiae. Genetics, 139, 1521–1532. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ogawa H., Johzuka,K., Nakagawa,T., Leem,S.H. and Hagihara,A.H. (1995) Functions of the yeast meiotic recombination genes, MRE11 and MRE2. Adv. Biophys., 31, 67–76. [DOI] [PubMed] [Google Scholar]
- 27.Sharples G.J. and Leach,D.R. (1995) Structural and functional similarities between the SbcCD proteins of Escherichia coli and the RAD50 and MRE11 (RAD32) recombination and repair proteins of yeast. Mol. Microbiol., 17, 1215–1217. [DOI] [PubMed] [Google Scholar]
- 28.Dolganov G.M., Maser,R.S., Novikov,A., Tosto,L., Chong,S., Bressan,D.A. and Petrini,J.H. (1996) Human Rad50 is physically associated with human Mre11: identification of a conserved multiprotein complex implicated in recombinational DNA repair. Mol. Cell. Biol., 16, 4832–4841. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Petrini J.H., Walsh,M.E., DiMare,C., Chen,X.N., Korenberg,J.R. and Weaver,D.T. (1995) Isolation and characterization of the human MRE11 homologue. Genomics, 29, 80–86. [DOI] [PubMed] [Google Scholar]
- 30.Carney J.P., Maser,R.S., Olivares,H., Davis,E.M., Le Beau,M., Yates,J.R., Hays,L., Morgan,W.F. and Petrini,J.H. (1998) The hMre11/hRad50 protein complex and Nijmegen breakage syndrome: linkage of double-strand break repair to the cellular DNA damage response. Cell, 93, 477–486. [DOI] [PubMed] [Google Scholar]
- 31.Matsuura S., Tauchi,H., Nakamura,A., Kondo,N., Sakamoto,S., Endo,S., Smeets,D., Solder,B., Belohradsky,B.H., Der Kaloustian,V.M., Oshimura,M., Isomura,M., Nakamura,Y. and Komatsu,K. (1998) Positional cloning of the gene for Nijmegen breakage syndrome. Nature Genet., 19, 179–181. [DOI] [PubMed] [Google Scholar]
- 32.Tavassoli M., Shayeghi,M., Nasim,A. and Watts,F.Z. (1995) Cloning and characterisation of the Schizosaccharomyces pombe rad32 gene: a gene required for repair of double strand breaks and recombination. Nucleic Acids Res., 23, 383–388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Varon R., Vissinga,C., Platzer,M., Cerosaletti,K.M., Chrzanowska,K.H., Saar,K., Beckmann,G., Seemanova,E., Cooper,P.R., Nowak,N.J., Stumm,M., Weemaes,C.M., Gatti,R.A., Wilson,R.K., Digweed,M., Rosenthal,A., Sperling,K., Concannon,P. and Reis,A. (1998) Nibrin, a novel DNA double-strand break repair protein, is mutated in Nijmegen breakage syndrome. Cell, 93, 467–476. [DOI] [PubMed] [Google Scholar]
- 34.Luo G., Yao,M.S., Bender,C.F., Mills,M., Bladl,A.R., Bradley,A. and Petrini,J.H. (1999) Disruption of mRad50 causes embryonic stem cell lethality, abnormal embryonic development and sensitivity to ionizing radiation. Proc. Natl Acad. Sci. USA, 96, 7376–7381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Xiao Y. and Weaver,D.T. (1997) Conditional gene targeted deletion by Cre recombinase demonstrates the requirement for the double-strand break repair Mre11 protein in murine embryonic stem cells. Nucleic Acids Res., 25, 2985–2991. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Shiloh Y. (1997) Ataxia-telangiectasia and the Nijmegen breakage syndrome: related disorders but genes apart. Annu. Rev. Genet., 31, 635–662. [DOI] [PubMed] [Google Scholar]
- 37.Digweed M., Reis,A. and Sperling,K. (1999) Nijmegen breakage syndrome: consequences of defective DNA double strand break repair. Bioessays, 21, 649–656. [DOI] [PubMed] [Google Scholar]
- 38.Ito A., Tauchi,H., Kobayashi,J., Morishima,K., Nakamura,A., Hirokawa,Y., Matsuura,S., Ito,K. and Komatsu,K. (1999) Expression of full-length NBS1 protein restores normal radiation responses in cells from Nijmegen breakage syndrome patients. Biochem. Biophys. Res. Commun., 265, 716–721. [DOI] [PubMed] [Google Scholar]
- 39.Stewart G.S., Maser,R.S., Stankovic,T., Bressan,D.A., Kaplan,M.I., Jaspers,N.G., Raams,A., Byrd,P.J., Petrini,J.H. and Taylor,A.M. (1999) The DNA double-strand break repair gene hMRE11 is mutated in individuals with an ataxia-telangiectasia-like disorder. Cell, 99, 577–587. [DOI] [PubMed] [Google Scholar]
- 40.Gatei M., Young,D., Cerosaletti,K.M., Desai-Mehta,A., Spring,K., Kozlov,S., Lavin,M.F., Gatti,R.A., Concannon,P. and Khanna,K. (2000) ATM-dependent phosphorylation of nibrin in response to radiation exposure. Nature Genet., 25, 115–119. [DOI] [PubMed] [Google Scholar]
- 41.Kim S.T., Lim,D.S., Canman,C.E. and Kastan,M.B. (1999) Substrate specificities and identification of putative substrates of ATM kinase family members. J. Biol. Chem., 274, 37538–37543. [DOI] [PubMed] [Google Scholar]
- 42.Lim D.S., Kim,S.T., Xu,B., Maser,R.S., Lin,J., Petrini,J.H. and Kastan,M.B. (2000) ATM phosphorylates p95/nbs1 in an S-phase checkpoint pathway. Nature, 404, 613–617. [DOI] [PubMed] [Google Scholar]
- 43.Zhao S., Weng,Y.C., Yuan,S.S., Lin,Y.T., Hsu,H.C., Lin,S.C., Gerbino,E., Song,M.H., Zdzienicka,M.Z., Gatti,R.A., Shay,J.W., Ziv,Y., Shiloh,Y. and Lee,E.Y. (2000) Functional link between ataxia-telangiectasia and Nijmegen breakage syndrome gene products. Nature, 405, 473–477. [DOI] [PubMed] [Google Scholar]
- 44.Lombard D.B. and Guarente,L. (2000) Nijmegen breakage syndrome disease protein and MRE11 at PML nuclear bodies and meiotic telomeres. Cancer Res., 60, 2331–2334. [PubMed] [Google Scholar]
- 45.Bertuch A. and Lundblad,V. (1998) Telomeres and double-strand breaks: trying to make ends meet. Trends Cell Biol., 8, 339–342. [DOI] [PubMed] [Google Scholar]
- 46.Boulton S.J. and Jackson,S.P. (1998) Components of the Ku-dependent non-homologous end-joining pathway are involved in telomeric length maintenance and telomeric silencing. EMBO J., 17, 1819–1828. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Moreau S., Ferguson,J.R. and Symington,L.S. (1999) The nuclease activity of Mre11 is required for meiosis but not for mating type switching, end joining, or telomere maintenance. Mol. Cell. Biol., 19, 556–566. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Chamankhah M. and Xiao,W. (1999) Formation of the yeast Mre11–Rad50–Xrs2 complex is correlated with DNA repair and telomere maintenance. Nucleic Acids Res., 27, 2072–2079. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Zhu X.D., Kuster,B., Mann,M., Petrini,J.H. and Lange,T. (2000) Cell-cycle-regulated association of RAD50/MRE11/NBS1 with TRF2 and human telomeres. Nature Genet., 25, 347–352. [DOI] [PubMed] [Google Scholar]
- 50.Paull T.T. and Gellert,M. (1998) The 3′ to 5′ exonuclease activity of Mre 11 facilitates repair of DNA double-strand breaks. Mol. Cell, 1, 969–979. [DOI] [PubMed] [Google Scholar]
- 51.Trujillo K.M., Yuan,S.S., Lee,E.Y. and Sung,P. (1998) Nuclease activities in a complex of human recombination and DNA repair factors Rad50, Mre11 and p95. J. Biol. Chem., 273, 21447–21450. [DOI] [PubMed] [Google Scholar]
- 52.Paull T.T. and Gellert,M. (1999) Nbs1 potentiates ATP-driven DNA unwinding and endonuclease cleavage by the Mre11/Rad50 complex. Genes Dev., 13, 1276–1288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Paull T.T. and Gellert,M. (2000) A mechanistic basis for Mre11-directed DNA joining at microhomologies. Proc. Natl Acad. Sci. USA, 97, 6409–6414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ono M., Tucker,P.W. and Capra,J.D. (1994) Production and characterization of recombinant human Ku antigen. Nucleic Acids Res., 22, 3918–3924. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Henricksen L.A., Umbricht,C.B. and Wold,M.S. (1994) Recombinant replication protein A: expression, complex formation and functional characterization. J. Biol. Chem., 269, 11121–11132. [PubMed] [Google Scholar]
- 56.Benson F.E., Baumann,P. and West,S.C. (1998) Synergistic actions of Rad51 and Rad52 in recombination and DNA repair. Nature, 391, 401–404. [DOI] [PubMed] [Google Scholar]
- 57.Sugiyama T., New,J.H. and Kowalczykowski,S.C. (1998) DNA annealing by RAD52 protein is stimulated by specific interaction with the complex of replication protein A and single-stranded DNA. Proc. Natl Acad. Sci. USA, 95, 6049–6054. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Shinohara A., Shinohara,M., Ohta,T., Matsuda,S. and Ogawa,T. (1998) Rad52 forms ring structures and co-operates with RPA in single-strand DNA annealing. Genes Cell., 3, 145–156. [DOI] [PubMed] [Google Scholar]
- 59.Mortensen U.H., Bendixen,C., Sunjevaric,I. and Rothstein,R. (1996) DNA strand annealing is promoted by the yeast Rad52 protein. Proc. Natl Acad. Sci. USA, 93, 10729–10734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Reddy G., Golub,E.I. and Radding,C.M. (1997) Human Rad52 protein promotes single-strand DNA annealing followed by branch migration. Mutat. Res., 377, 53–59. [DOI] [PubMed] [Google Scholar]
- 61.Gerecke E.E. and Zolan,M.E. (2000) An mre11 mutant of coprinus cinereus has defects in meiotic chromosome pairing, condensation and synapsis. Genetics, 154, 1125–1139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Tsukamoto Y., Kato,J. and Ikeda,H. (1996) Effects of mutations of RAD50, RAD51, RAD52 and related genes on illegitimate recombination in Saccharomyces cerevisiae. Genetics, 142, 383–391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Hopfner K.P., Karcher,A., Shin,D.S., Craig,L., Arthur,L.M., Carney,J.P. and Tainer,J.A. (2000) Structural biology of Rad50 ATPase: ATP-driven conformational control in DNA double-strand break repair and the ABC-ATPase superfamily. Cell, 101, 789–800. [DOI] [PubMed] [Google Scholar]
- 64.Melby T.E., Ciampaglio,C.N., Briscoe,G. and Erickson,H.P. (1998) The symmetrical structure of structural maintenance of chromosomes (SMC) and MukB proteins: long, antiparallel coiled coils, folded at a flexible hinge. J. Cell Biol., 142, 1595–1604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.New J.H., Sugiyama,T., Zaitseva,E. and Kowalczykowski,S.C. (1998) Rad52 protein stimulates DNA strand exchange by Rad51 and replication protein A. Nature, 391, 407–410. [DOI] [PubMed] [Google Scholar]
- 66.Shinohara A., Ogawa,H. and Ogawa,T. (1992) Rad51 protein involved in repair and recombination in S.cerevisiae is a RecA-like protein. Cell, 69, 457–470. [DOI] [PubMed] [Google Scholar]
- 67.Shinohara A. and Ogawa,T. (1998) Stimulation by Rad52 of yeast Rad51-mediated recombination. Nature, 391, 404–407. [DOI] [PubMed] [Google Scholar]
- 68.Sung P. (1997) Function of yeast Rad52 protein as a mediator between replication protein A and the Rad51 recombinase. J. Biol. Chem., 272, 28194–28197. [DOI] [PubMed] [Google Scholar]
- 69.Malone R.E., Ward,T., Lin,S. and Waring,J. (1990) The RAD50 gene, a member of the double strand break repair epistasis group, is not required for spontaneous mitotic recombination in yeast. Curr. Genet., 18, 111–116. [DOI] [PubMed] [Google Scholar]
- 70.Glasunov A.V., Glaser,V.M. and Kapultsevich,Y.G. (1989) Two pathways of DNA double-strand break repair in G1 cells of Saccharomyces cerevisiae. Yeast, 5, 131–139. [DOI] [PubMed] [Google Scholar]