
Figure 3.
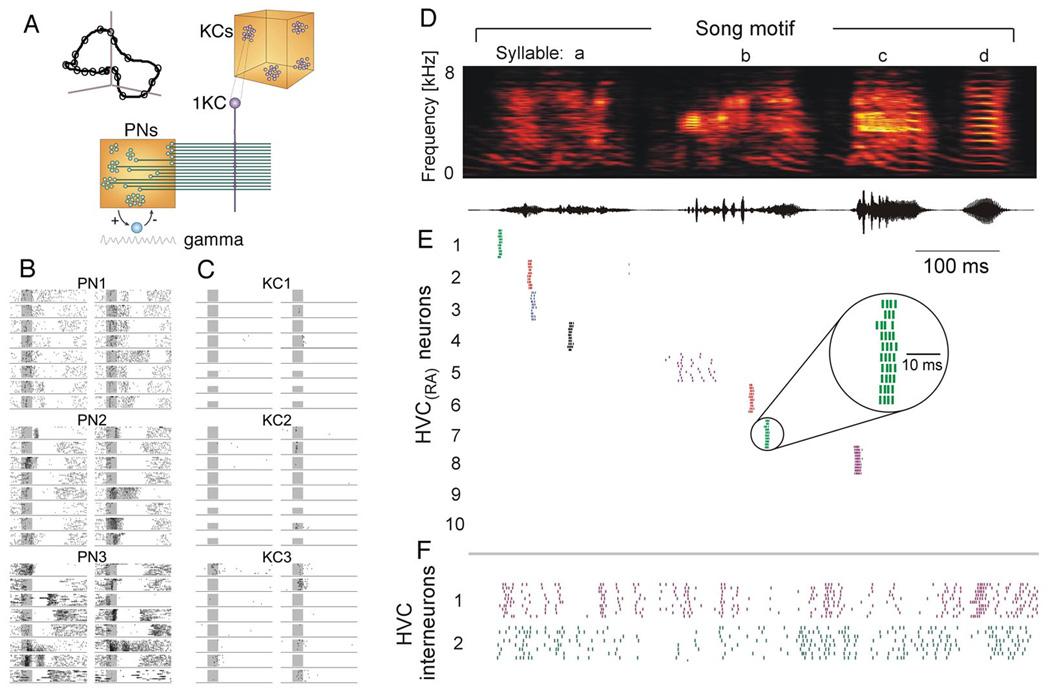
Externally-triggered and internally-generated assembly sequences. A. Wiring diagram of the early olfactory system of the locust. An odorant evokes an odor-specific temporal pattern in several of recurrently connected antennal lobe (AL) neurons, coordinated by a 20–30 Hz (gamma) oscillation. Kenyon cells (KC) of the mushroom body (MB) are the readers of the activity of AL projection neurons (PNs) and integrate their spikes. B. Firing patterns of 3 AL neurons (PN1–3) in response to 16 different odors. In each segment of time (e.g., a gamma oscillation cycle), a different constellation of AL neurons fires. This constellation is referred to as the ‘population vector’ or ‘state’ of the network, and the time-varying population vector (i.e., the shifting states) is described as a trajectory. Although each state of the trajectory codes for the same odor, the state evolves over a few hundred milliseconds before relaxing back to baseline activity (illustrated by the curve in the inset in A). C. Activity of 3 KCs. Each KC carries out a pattern matching operation between its connection vector and the PN population activity vector. The AL output synchrony is strongest at its early phase of evolution and evokes a single burst in the reader KC (‘sparse coding’). D. Time-frequency spectrum of a zebra finch song and its amplitude envelope. E. Spike raster plot of eight projection neurons in the high vocal center (HVC). There is not a one-to-one correspondence between the song syntax and cell assemblies (neural ‘letters’ in HVC). Rather, neurons in HVC generate a temporal ‘state’ sequence (i.e., a trajectory), allowing the target neuronal activity and consequent motor actions unfold. F. Interneuronal activity is also temporally organized and relates to the syntactic structure of the song (F). A to C, modified after Laurent (2002). D to F, modified after Hahnloser et al. (2002).