

Abstract
Dynamic behaviors of liposomes caused by interactions between liposomal membranes and surfactant were studied by direct real-time observation by using high-intensity dark-field microscopy. Solubilization of liposomes by surfactants is thought to be a catastrophic event akin to the explosion of soap bubbles in the air; however, the actual process has not been clarified. We studied this process experimentally and found that liposomes exposed to various surfactants exhibited unusual behavior, namely continuous shrinkage accompanied by intermittent quakes, release of encapsulated liposomes, opening up, and inside–out topological inversion.
Liposomes (which are closed membrane vesicles) have been well studied as simplified models of biological membranes (1–5) and are now used in a number of applications (5, 6), for example, as carriers of drug or DNA delivery or as artificial membranes for reconstructing membranous enzyme activities (7–9). Recently, many important phenomena affecting lipid bilayers, including their detergent solubilization, have been explored by using liposomes; such studies promote a better understanding of the biophysical properties of bilayer membranes and moreover will improve the handling of membrane proteins when they are isolated from or reconstructed into lipid bilayers (10–13). However, studies of intermediate stages in the detergent solubilization of liposomes are only now in progress (14–17), and the interaction mechanism between membranes and surfactants has remained unclear. Therefore, real-time approaches by using optical microscopy to study the dynamic behavior of liposomes are very important.
High-intensity dark-field microscopy has enabled us to obtain real-time high-contrast images of giant unilamellar liposomes in aqueous solutions (18–22). In this study, we used such techniques to characterize the interactions between liposomal membranes and surfactants. Eight kinds of liposomes and various types of surfactants (Fig. 1) were mixed in all possible combinations in a mixing chamber to generate a concentration gradient of each surfactant for microscope specimens, and morphological changes of liposomes exposed to those surfactants were monitored (18, 23). In the absence of surfactant, liposomal membranes were spherical, and thermal fluctuations of their spherical shape were largely suppressed by the surface tension of their membranes. Hereafter, this morphological state of liposomes will be called tense. In this study, we found several unusual behaviors of liposomes (which are published as supplemental data on the PNAS web site, www.pnas.org).
Figure 1.

Summary of the solubilization processes. C, continuous-stepwise or -smooth shrinkage; I, inside-out inversion; O, opening up; B, burst. Lipids: PC, phosphatidylcholine; DMPC, dimyristoyl phosphatidylcholine; PG, phosphatidylglycerol; DMPG, dimyristoyl phosphatidylglycerol; PA, phosphatidic acid; DMPA, dimyristoyl phosphatidic acid; DMDAP, 1,2-dimyristoyl 3-dimethylammonium propane; DMTAP, 1,2-dimyristoyl 3-trimethylammonium propane. Among eight kinds of lipids, PC, PG, and PA are isolated from egg yolk or other natural sources, so that they had nonuniform lengths of acyl chains, and others had uniform tail lengths (14 C). PC, DMPC, PG, and DMPG had large head groups, and PA, DMPA, DMDAP and DMTAP had small head groups. Among the eight kinds of liposomes used, seven had binary lipid compositions (1:1 mol/mol). The charge carried by liposomes at neutral pH is shown. The surfactants shown here were added to each liposome solution. Abbreviations for surfactants: C12E8, polyoxyethylene 8 lauryl ether; CHAPS, 3-[(3-cholamidopropyl) dimethylammonio]-1-propane sulfonate; SB 3–14, N-tetradecyl-N, N-dimethyl-3-ammonio-1-propane sulfonate; HPC, hexadecyl pyridinium chloride; HTAB, hexadecyl trimethyl ammonium bromide. The charge carried by each surfactant at neutral pH and its critical micelle concentration (mM) is shown. We usually selected giant liposomes whose diameters exceeded 5 μm for observations to make them and later analysis easy. The experimental results shown here were essentially not altered even when the concentration of surfactant was changed severalfold; however, the rate of liposomal solubilization in each process was changed. It is noted that, by monitoring the changing of solution turbidity, we confirmed that liposomal solubilization started when the concentrations of each added surfactant were higher than its critical micelle concentration.
Materials and Methods
Preparation and Observation of Liposomes.
To prepare giant unilamellar liposome, liposome (total 1 mM lipid concentration) was made of phosphatidylcholine (PC) or of PC and one of seven other lipids (1:1, mol/mol) in Hepes buffer (10 mM Hepes–NaOH, pH 7.0), as described previously (18, 21, 22). Lipids were dissolved in a chloroform/methanol solution, 98:2 (vol/vol), and mixed. The organic solvent was evaporated under a flow of nitrogen gas, and the lipids were further dried in vacuo for at least 90 min. Hepes buffer was then added to the dried lipid films at 25°C. The lipid films immediately started swelling to form liposomes, and swelling was facilitated by agitating the test tubes occasionally by hand. Liposomes were observed at 25°C by using dark-field microscopy (BHF, Olympus, Tokyo), as described previously. Images were recorded by using an SIT video camera (C-2400–08, Hamamatsu Photonics, Hamamatsu City, Japan) and were further processed with ip-lab spectrum (Scanalytics, Fairfax, VA) and analyzed with NIH image and photoshop (Adobe Systems, Mountain View, CA).
Mixing of Liposomes with Surfactant.
Each giant liposome solution was mixed with each surfactant (100 mM) dissolved in Hepes buffer at 25°C. To make a concentration gradient of surfactant in a microscope specimen, we used a mixing chamber made of a glass slide and a coverslip, which were firmly fixed together with spacers (23). To apply each surfactant to a liposome solution, an excess volume of surfactant solution was placed on an open site of the mixing chamber into which a liposome solution had been injected from another open site. These two solutions are gently attached and mixed in the chamber, and thereby a concentration gradient of surfactant was spontaneously generated by diffusion. Slowly moving liposomes were followed in the microscope, and behaviors of liposomes in a concentration gradient of the surfactant were monitored.
Lipids and Surfactants.
Octyl glucoside, Triton X-100, and SDS were purchased from Nacalai Tesque (Kyoto). Tween 20 and 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS) were purchased from Wako Pure Chemicals (Osaka). Hexadecyl pyridinium chloride was purchased from Katayama Chemicals (Osaka). 1, 2-dimyristoyl 3-trimethylammonium propane (DMTAP) and 1, 2-dimyristoyl 3-dimethylammonium propane (DMDAP) were purchased from Avanti Polar Lipids. Other surfactants and lipids were purchased from Sigma.
Results
Shrinkage of Liposomes Caused by Lipid Solubilization with Surfactant.
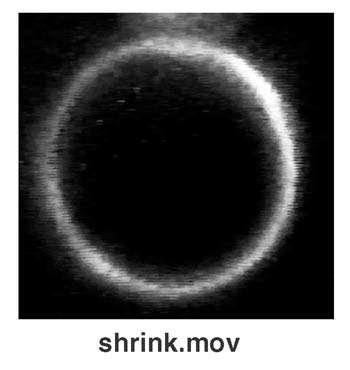
First, we describe intermittent quakes of liposomes coupled with their continuous-stepwise shrinkage (Fig. 2A). Liposomal quakes are transitions from the tense state to the transient state that are characterized by vigorous fluctuations in their spherical shape. Transition from the tense to the quaking state was a rapid process accomplished within the time-resolution limit (1/30 sec), whereas recovery from the quaking to the tense state was a slow process on the second scale. Liposomes quaked individually and intermittently and decreased their size step by step (Fig. 2B). The interval between quakes decreased as the liposomal size decreased (Fig. 2C). Our preparations of liposomes contained giant liposomes encapsulating baby liposomes (a few micrometers in size), and those giant liposomes never released their babies externally so long as they remained tense. However, giant liposomes directly released their babies during their quaking periods (Fig. 2D) (24). These observations indicate that liposomal membranes that are in contact with surfactant temporally open holes as large as a few micrometers in size, while they maintain their overall shape and continuity. Small liposomes encapsulated by giant ones never shrank; however, once exposed to the surfactant, they began to shrink similarly to the giant liposomes.
Figure 2.

Continuous shrinkage of a liposome. (A) A sequence of photographs showing a PC liposome that is alternating between tense (T) and quaking (Q) states in the presence of Triton X-100. (B) Stepwise shrinkage of the liposome shown in A. Duration of quakes can be obtained from gaps between the two solid lines. (C) Dependence of the cycle time on liposomal size. (D) Sequential photographs of a giant PC liposome releasing an encapsulated liposome in the presence of Triton X-100. The inner liposome was excluded when the outer liposome was quaking. (E) Sequential photographs of a PC giant liposome that is shrinking in the presence of SDS. (F) Time-dependent decrease in the surface area of a liposome; the fitted line is exponential. (G) A model for the process of continuous shrinkage of a liposome. (Bars = 5 μm.)
Continuous-stepwise shrinkage with intermittent quakes is the pathway of liposomal solubilization when liposomes are exposed to a nonionic surfactant (Fig. 1). However, when an ionic surfactant replaced a nonionic one, liposomes decreased their size smoothly (Fig. 2E). In this case, surface areas calculated from the diameters of liposomes decreased exponentially with time (Fig. 2F). We assume that liposomes quaked at various magnitudes depending on the nature of the surfactant, and that continuous-smooth shrinkage was brought about by invisible magnitudes of membrane quakes. In the case of continuous-stepwise shrinkage, which accompanies the intermittent quaking, lipids may also be excluded continuously from a liposomal membrane. The liposome could, however, discharge water molecules from its interior, as well as encapsulated vesicles, only during the intermittent quaking by perforation of its lipid bilayer (Fig. 2 B and G). Liposomes vigorously fluctuate their membranes within each quaking state because of membrane perforation and the sudden decrease in internal pressure that results from the discharge of the water within.
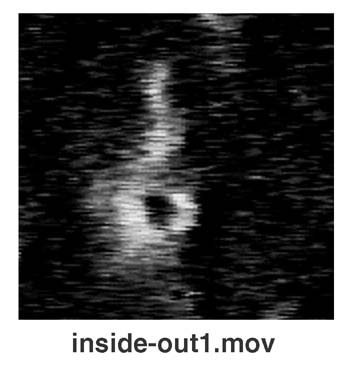
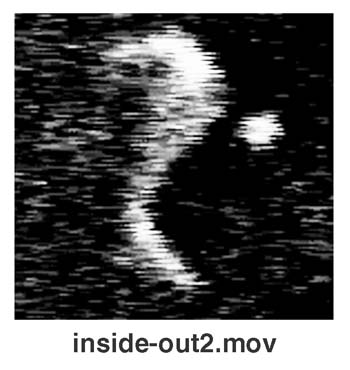
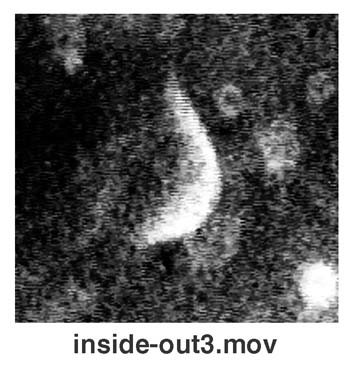
Liposomal Inside–Out Inversion.
A remarkable behavior of liposomes (Fig. 3A) was found when we examined the two combinations marked “I” in Fig. 1. In those cases, liposomal membranes were opened and transformed into lipid bilayer sheets, turned themselves inside out, and then closed again. The liposomal behavior of this inside–out topological inversion is demonstrated in this study. Liposomes repeated this topological inversion several times, decreasing their size step by step (Fig. 3B). After repeating such inside–out inversions several times, the lipid bilayer sheets often became unable to close and were then solubilized over their entire surface areas equally. In this case, it seems likely that the surfactant molecules penetrate into and exclude lipids from only the side of the lipid bilayer (the outer leaflet), which is exposed to the solution containing the surfactant (Fig. 3C). Although lipids may be continuously excluded from the outer leaflet of a liposomal membrane, the liposome cannot decrease its size and remains tense because the rapid escape of interior water molecules is prevented by the intact inner half layer of its membrane. If a flip-flop translocation of lipid molecules does not take place in the membrane, the surface tensions working in each of the two leaflets of the liposomal membrane will lose balance, resulting in the inside–out inversion that is eventually induced in the liposome (Fig. 3C).
Figure 3.

Inside-out inversion of a liposome. (A) A sequence of photographs showing a liposome (PC: DMTAP) undergoing inside-out inversion in the presence of hexadecyl trimethyl ammonium bromide (HTAB). (B) Stepwise shrinkage of a liposome (PC: DMTAP) that is repeating an inside-out inversion in the presence of HTAB. Open circles show the time course for the decreasing diameter of a liposome (shown in the photographic sequence). The diameter decreased only after each inside-out inversion (I) and kept nearly constant during the tense state (T). The time required for an inversion is obtained from the gap between the two dotted lines. (C) A model for the inside-out inversion process of a liposome. (Bars = 5 μm.)
It has been suggested that the molecular shape of lipids in a bilayer membrane can determine the membrane curvature and shape (25, 26). The inside–out topological inversion of liposomes described in this study could be the first direct demonstration showing that a lipid bilayer that has different numbers of lipid molecules in two leaflets can occasionally generate forces strong enough to change the membrane curvature and/or topology.
Opening Up of Liposomes Caused by Surfactant.
The next behavior is the opening up of liposomes followed by solubilization of cup-shaped liposomal membranes from their exposed edges (Fig. 4A). This was the pathway of liposomal solubilization in those combinations of liposomes and surfactants that are marked as “O” in Fig. 1. Even when the concentrations of added surfactants were increased, most liposomes maintained a single opening hole, although that hole opened larger and liposomes solubilized faster. This result indicates that the added surfactant molecules accumulate cooperatively along the membrane-free verge (Fig. 4B).
Figure 4.

Opening-up and subsequent solubilization of a liposome. (A) A sequence of photographs showing the solubilization of a liposome (PC: DMTAP) caused by SDS. (B) A model for the opening-up and subsequent solubilization of a liposome caused by a surfactant. The surfactant molecules probably accumulate cooperatively along the membrane free verge of the opened cup shape liposome. (Bar = 5 μm.)
We attempted to reseal the cup-shaped liposomal membranes by decreasing the concentrations of surfactants by dilution; however, this was never successful. These observations indicate that the opening up of liposomal membranes by surfactants is basically different from the reversible opening up induced by talin (22), a cytoskeleton associate protein.
Liposomal Burst.
Finally, we should refer to the bursting of liposomes (Fig. 5), which is catastrophic and akin to the bursting of soap bubbles in the air. This was the process of liposomal solubilization in combinations marked “B” in Fig. 1. Irrespective of liposomal size, such bursting was accomplished within several tens of milliseconds, which was nearly to the time-resolution limit. In rare cases, liposome showed membrane fluctuation immediately before the burst. Several liposomes in a microscope field burst temporally at random over a time span of a few seconds. Added surfactants are probably able to penetrate the lipid bilayer faster than their solubilizing action or ability to cooperatively extract membrane components by micelles, and this causes instantaneous solubilization of liposomes (Fig. 5B). The liposomal burst occurred frequently when liposomes made of mixed phospholipids possessing inhomogeneous acyl chain lengths were used or when ampholytic surfactant or surfactant, which has a relatively large hydrophilic region, was added to liposomes. Irregularity of acyl chain length of lipid molecules is known as the factor that influences membrane fluidity (27), indicating that molecular fluidity of membranes might be important for the liposomal burst.
Figure 5.

Bursting of a liposome. (A) Sequential photographs showing the bursting of a liposome [PC: phosphatidic acid (PA)] in the presence of 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS). The time resolution of our apparatus was 1/30 sec. (Bar = 5 μm.) (B) A model for the liposomal burst caused by the detergent solubilization.
General Features of the Transformation Processes.
Liposomes were solubilized via various processes, as shown in Fig. 1. However, with a given combination of liposome lipid composition and surfactant, all of the liposomes exhibited only one specific transformation process, even at various ratios of surfactant to lipid and observable liposome sizes between 1 and 20 μm (date not shown). Exceptions, however, were C/B or C/O in Fig. 1. In the case of C/B, approximately half of the liposomes showed continuous shrinkage, and another half displayed bursting. Similarly, in C/O, the frequency ratio of continuous shrinkage to opening up was about one to one. Within an area in a microscope specimen where liposomes and surfactant were appropriately mixed, all of the type of transformations shown in Fig. 1 occurred frequently.
Inside–out and bursting transformations appear to take place transiently in response to an increasing concentration of surfactant. On the other hand, cup-shaped liposomes produced in the opening-up process were relatively stable in a fixed concentration of surfactant, indicating an equilibrium-binding reaction of the surfactant and the liposome membrane.
Discussion
In this study, we observed a variety of phenomena exhibited by liposomal membranes interacting with surfactants, including intermittent quaking, inside-out inversion, opening up, and bursting. These phenomena might be based not only on the cooperative interaction between a lipid membrane and a surfactant but also on the cooperative movement of lipid molecules in a membrane (Figs. 2G, 3C, 4B, and 5B). Our results show that the transformation pathway depends on the combination a particular surfactant with a particular lipid membrane composition, and the electric charge carried by both the lipid and surfactant is an important factor in determining transformation pathway (Fig. 1). In contrast, the critical micelle concentration of the added surfactant, the sizes of the hydrophilic head, and the acyl chain length of lipid molecules composing the membrane are not strong factors in determining the pathway.
What mechanism works to determine which pathway takes place among various topological transformations of liposomes? Our data suggest that membrane solubilization progresses through the following steps: (i) Surfactant molecules associate with or penetrate into lipid bilayers; (ii) with increase in surfactant concentrations, a lipid bilayer is saturated with surfactant molecules, so that lipid molecules are extracted from the membrane by forming detergent-lipid micelles. In i, the main determining factor seems to be whether surfactant molecules penetrate into the membrane homogeneously or accumulate locally on a liposome. In ii, the primary factor seems to be whether the flip-flop translocation of lipid and/or surfactant molecules across the leaflets of the bilayer can take place or not, as shown in Figs. 2G and 3C (28, 29).
The inside-out inversion of liposomes found in this study could be one of the direct demonstrations showing the close relationship between membrane curvature and its topology. Shape transformation of a liposome caused by differences in expansivities of the two leaflets in a membrane has been investigated both theoretically and experimentally, by using the bilayer coupling model (30, 31). These studies, however, described only continuous processes such as budding formations or stomatocyte transformations. On the other hand, the liposomal inside-out inversion is an entirely discontinuous one. In our case, liposome membranes continuously lost their component lipids, probably from only their outer leaflets, by detergent solubilization. If there is no or little flip-flop movement between two leaflets, number density of lipid molecules in the outer leaflet will continuously decrease (Fig. 3C). As a result, a surface area of the outer leaflet tends to shrink, whereas that of the inner one is maintained. This imbalance in the surface areas causes potential force to invert a curvature of the bilayer membrane. Therefore, once a liposome membrane is damaged at some point, inside-out transformation will take place. After every inversion, the liposome decreased about 20% of its surface area (Fig. 3B).
Electric charges carried by lipid and surfactant molecules play a basic role in the interaction between them. When a membrane containing positively charged lipids was mixed with a surfactant that carries a positive charge, hexadecyl trimethyl ammonium bromide (HTAB) or SB 3–14, the inversion took place (Fig. 1). On the other hand, the inversion never happened when anionic liposomes were mixed with an anionic surfactant, so that simple electrical repulsion force cannot explain the mechanism. It has been reported that some cationic amphipathic reagents cannot move across the leaflets of a bilayer (32). Such restriction of cationic molecules on the translocation within lipid bilayer may play an important role in the liposomal inside-out inversion. By changing the pH or salt strength of the solution, we are currently investigating in more detail how electric charges carried by lipid and surfactant molecules affect liposome transformations.
Recently, a number of surfactants has been investigated on the effectiveness for studying membrane and membrane proteins. As shown in Fig. 1, octyl glucoside and cholate solubilized almost all liposomes via the continuous shrinkage process. This property may be a reason why these detergents have been found to be favorable for many studies of membrane and membrane proteins. Liposomes underwent the opening-up transformation either by cationic surfactants or with a membrane contained positively charged synthetic lipid. Moreover, when positively charged liposomes were mixed with a surfactant that carries positive charge, especially HTAB, the inversion transformation took place (Fig. 1). These results suggest a possible reason why a number of biological toxins (e.g., antibiotic peptides) that kill cells by attacking their surface membranes are cationic, and why positively charged lipids are uncommon in living cells.
Our study shows that a lipid membrane vesicle itself has unexpected capabilities for topological transformations depending on the solution conditions. Physiologically, biological organisms possess a variety of biosurfactants. These capabilities seem too dynamic for biological membranes, so that living organisms might have developed a more controllable vesicular system by using membrane proteins and membrane cytoskeletons. Finally, our mechanistic interpretation may be regarded as suggestive, rather than as providing firm new insights into the processes and factors affecting liposome stability and solubilization more generally, and thus this report impresses us as raising more questions than it answers. This study, however, may be useful for the design of new surfactants or for understanding how amphipathic toxins work and may contribute to pure and applied membrane sciences.
Supplementary Material
Acknowledgments
We thank Dr. Sho Asakura (Nagoya University) for his critical reading and valuable comments on the manuscript. This study was supported by Grants-in-Aid for Scientific Research (A) and (C) from the Ministry of Education, Science, Sports, and Culture of Japan.
Abbreviations
- DMTAP
1, 2-dimyristoyl 3-trimethylammonium propane
- PC
phosphatidylcholine
- PG
phosphatidylglycerol
Footnotes
This paper was submitted directly (Track II) to the PNAS office.
References
- 1.Bangham A D. BioEssays. 1995;17:1081–1088. doi: 10.1002/bies.950171213. [DOI] [PubMed] [Google Scholar]
- 2.Lipowsky R. Nature (London) 1991;349:475–481. doi: 10.1038/349475a0. [DOI] [PubMed] [Google Scholar]
- 3.Evans E A. Biophys J. 1974;14:923–931. doi: 10.1016/S0006-3495(74)85959-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Canham P B. J Theor Biol. 1970;26:61–81. doi: 10.1016/s0022-5193(70)80032-7. [DOI] [PubMed] [Google Scholar]
- 5.Sackmann E. Science. 1996;271:43–48. doi: 10.1126/science.271.5245.43. [DOI] [PubMed] [Google Scholar]
- 6.Lasic D D. In: Handbook of Biological Physics, Structure and Dynamics of Membranes. Lipowsky R, Sackmann E, editors. 1a. Amsterdam: Elsevier; 1995. pp. 491–519. [Google Scholar]
- 7.Lasic D D. Nature (London) 1996;380:561–562. doi: 10.1038/380561a0. [DOI] [PubMed] [Google Scholar]
- 8.Templeton N S, Lasic D D. Mol Biotechnol. 1999;11:175–180. doi: 10.1007/BF02915810. [DOI] [PubMed] [Google Scholar]
- 9.Steinberg-Yfrach G, Rigaud J L, Durantini E N, Moore A L, Gust D, Moore T A. Nature (London) 1998;392:479–482. doi: 10.1038/33116. [DOI] [PubMed] [Google Scholar]
- 10.Knol J, Sjollema K, Poolman B. Biochemistry. 1998;37:16410–16415. doi: 10.1021/bi981596u. [DOI] [PubMed] [Google Scholar]
- 11.Walter A. Mol Cell Biochem. 1990;99:117–123. doi: 10.1007/BF00230341. [DOI] [PubMed] [Google Scholar]
- 12.de Foresta B, le Maire M, Orlowski S, Champeil P, Lund S, Moller J V, Michelangeli F, Lee A G. Biochemistry. 1989;28:2558–2567. doi: 10.1021/bi00432a032. [DOI] [PubMed] [Google Scholar]
- 13.Womack M D, Kendall D A, MacDonald R C. Biochim Biophys Acta. 1983;733:210–215. doi: 10.1016/0005-2736(83)90524-2. [DOI] [PubMed] [Google Scholar]
- 14.Kragh-Hansen U, le Maire M, Moller J V. Biophys J. 1998;75:2932–2946. doi: 10.1016/S0006-3495(98)77735-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Lopez O, Cocera M, Wehrli E, Parra J L, de la Maza A. Arch Biochem Biophys. 1999;367:153–160. doi: 10.1006/abbi.1999.1267. [DOI] [PubMed] [Google Scholar]
- 16.de la Maza A, Parra J L. Biophys J. 1997;72:1668–1675. doi: 10.1016/S0006-3495(97)78812-X. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Urbaneja M A, Alonso A, Gonzalez-Manas J M, Goni F M, Partearroyo M A, Tribout M, Paredes S. Biochem J. 1990;270:305–308. doi: 10.1042/bj2700305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Hotani H. J Mol Biol. 1984;178:113–120. doi: 10.1016/0022-2836(84)90234-1. [DOI] [PubMed] [Google Scholar]
- 19.Hotani H, Miyamoto H. Adv Biophys. 1990;26:135–156. doi: 10.1016/0065-227x(90)90010-q. [DOI] [PubMed] [Google Scholar]
- 20.Miyata H, Hotani H. Proc Natl Acad Sci USA. 1992;89:11547–11551. doi: 10.1073/pnas.89.23.11547. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Honda M, Takiguchi K, Ishikawa S, Hotani H. J Mol Biol. 1999;287:293–300. doi: 10.1006/jmbi.1999.2592. [DOI] [PubMed] [Google Scholar]
- 22.Saitoh A, Takiguchi K, Tanaka Y, Hotani H. Proc Natl Acad Sci USA. 1998;95:1026–1031. doi: 10.1073/pnas.95.3.1026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Takiguchi K. J Biochem. 1991;109:520–527. doi: 10.1093/oxfordjournals.jbchem.a123414. [DOI] [PubMed] [Google Scholar]
- 24.Menger F M, Keiper J S. Curr Opin Chem Biol. 1998;2:726–732. doi: 10.1016/s1367-5931(98)80110-5. [DOI] [PubMed] [Google Scholar]
- 25.Scales S J, Scheller R H. Nature (London) 1999;401:123–124. doi: 10.1038/43582. [DOI] [PubMed] [Google Scholar]
- 26.Schmidt A, Wolde M, Thiele C, Fest W, Kratzin H, Podtelejnikov A V, Witke W, Huttner W B, Soling H D. Nature (London) 1999;401:133–141. doi: 10.1038/43613. [DOI] [PubMed] [Google Scholar]
- 27.Koynova R, Caffrey M. Biochim Biophys Acta. 1998;1376:91–145. doi: 10.1016/s0304-4157(98)00006-9. [DOI] [PubMed] [Google Scholar]
- 28.McNamee M G, McConnell H M. Biochemistry. 1973;12:2951–2958. doi: 10.1021/bi00740a001. [DOI] [PubMed] [Google Scholar]
- 29.Higgins C F. Cell. 1994;79:393–395. doi: 10.1016/0092-8674(94)90248-8. [DOI] [PubMed] [Google Scholar]
- 30.Seifert U, Berndl K, Lipowsky R. Phys Rev A. 1991;44:1182–1202. doi: 10.1103/physreva.44.1182. [DOI] [PubMed] [Google Scholar]
- 31.Käs J, Sackmann E. Biophys J. 1991;60:825–844. doi: 10.1016/S0006-3495(91)82117-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Betz W J, Mao F, Smith C B. Curr Opin Neurobiol. 1996;6:365–371. doi: 10.1016/s0959-4388(96)80121-8. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}