

Abstract
The moss Physcomitrella patens is increasingly being used as a model for plant systems biology studies. While genomic and transcriptomic resources are in place, tools and experimental conditions for proteomic studies need to be developed. In the present study we describe a rapid and efficient protocol for the simultaneous isolation of chloroplasts and mitochondria from moss protonema. Routinely, 60–100 μg mitochondrial and 3–5 mg chloroplast proteins, respectively, were obtained from 20 g fresh weight of green moss tissue. Using 14 plant compartment marker antibodies derived from seed plant and algal protein sequences, respectively, the evolutionary conservation of the compartment marker proteins in the moss was demonstrated and purity and intactness of the extracted organelles confirmed. This isolation protocol and these validated compartment markers may serve as basis for sub-cellular proteomics in P. patens and other mosses.
Electronic supplementary material
The online version of this article (doi:10.1007/s00299-010-0935-4) contains supplementary material, which is available to authorized users.
Keywords: Organelles, Compartment marker, Chloroplast proteins, Mitochondrial proteins, Physcomitrella, Bryophyte
Introduction
As the first non-seed plant with a completely sequenced genome (Rensing et al. 2008) the moss Physcomitrella patens has been established as a plant system to investigate the evolution of stress adaptation (Frank et al. 2007; Khandelwal et al. 2010) and of signalling events (Heintz et al. 2004, 2006) in early land plants. Along with these studies a wide range of high-throughput molecular biology tools has been developed and implemented in recent years (Richardt et al. 2007, 2010) paving the way for the use of this model organism for systems biology studies (Decker et al. 2006).
Focussing on plant organelles in a moss such as P. patens can be of special interest to obtain information on the evolution of metabolic compartmentalisation (Kopriva et al. 2007; Wiedemann et al. 2010), biosynthetic pathways (Stumpe et al. 2006) and protein sorting mechanisms (Kiessling et al. 2004, Mitschke et al. 2009, Richter et al. 2002). Of special interest are chloroplasts and mitochondria as they are semi-autonomous organelles of endosymbiotic origin with own DNA that encodes only for a small subset of proteins localised to these organelles. Hence, most of the proteins are nuclear-encoded and have to be imported into chloroplasts and mitochondria, respectively (Gray et al. 1999; Reski 2009; Strittmatter et al. 2010). The prediction of sub-cellular protein localisation, however, is error prone because the transit peptides are not well conserved (Bruce 2001) and prediction algorithms are usually trained on the basis of proteins from seed plants. Experimental data sets have shown that the tools currently available for the prediction of sub-cellular localisation can only identify about 50% of the proteins targeted to organelles (Heazlewood et al. 2004; Kleffmann et al. 2004). These limitations can only be overcome by the generation of species-specific training data sets for the respective organelles, the data sets being very much dependent on the specificity, i.e. correct prediction of the protein localisation (Baginski and Gruissem 2004; Salvi et al. 2008b). The generation of reliable data sets is, however, difficult as contaminations with proteins from other organelles and from the cytosol can never be ruled out during the isolation of single organelles.
Many protocols for the isolation of plant organelles in seed plants are established and have been used for subsequent high-throughput shotgun proteomic studies of Arabidopsis chloroplasts (Kleffmann et al. 2004; Baginski et al. 2005) and mitochondria (Heazlewood et al. 2004; Millar et al. 2001a, b; Sweetlove et al. 2007) or for example, the analysis of mitochondria in rice (Heazlewood et al. 2003; Huang et al. 2009). All these studies employ density gradients for the purification of organelles, sometimes combining it with free flow electrophoresis (FFE) to separate chloroplasts from mitochondria (Eubel et al. 2007; Huang et al. 2009; Lee et al. 2008). However, losses of about 50% of the organelle material can occur (Eubel et al. 2007), creating a need for the adaptation of existing protocols for each model species (Sweetlove et al. 2007).
For the moss P. patens protocols for the isolation of organelles via density gradients have been reported (Kabeya and Sato 2005; Kasten et al. 1997; Marienfeld et al. 1989). However, the moss material used in these experiments was always subjected to protoplastation, which besides from being a laborious and costly pre-treatment of the material might also have an effect on the physiological status of the cell and, hence, its proteome.
The aim of this study was to set up a protocol for the simultaneous isolation of highly enriched fractions of pure and intact chloroplasts and mitochondria from protonema tissue of P. patens. Integrity and purity of these fractions as well as potential contaminations were assessed using a set of plant compartment marker antibodies. The protocol presented here enables for the isolation of intact chloroplasts and mitochondria and delivers protein yields that are sufficient for sub-cellular proteomic studies in Physcomitrella. Such studies can provide the basis for a large-scale analysis of protein sorting mechanisms in moss and, in addition, unravel the evolution of metabolic and biosynthetic processes occurring in plant organelles.
Materials and methods
Plant material and growth conditions
Protonema of Physcomitrella patens (Hedw.) Bruch & Schimp. was cultured in modified liquid Knop medium according to Reski and Abel (1985) containing 250 mg/l KH2PO4, 250 mg/l KCl, 250 mg/l MgSO4 × 7 H2O, 1,000 mg/l Ca(NO3)2 × 4H2O and 12.5 mg/l FeSO4 × 7H2O (pH 5.8).
Protonema filaments were disrupted with an Ultra-Turrax (IKA, Staufen, Germany) at 18,000 rpm for 90 s before inoculation. Round-bottom flasks containing 5 l of medium were inoculated with 0.3 g dry weight and aerated with 0.3 vvm at 25°C under long day conditions [16 h light, 8 h dark, Osram TLD 36 W/25, 70 μmol/(m2 s)]. After 7 days, moss was harvested and filter-dried using a vacuum pump and a Büchner funnel.
Organelle isolation
All following steps were performed at 4°C and, where applicable, on wet ice. Typically, two 5 l flasks were used per experiment, which corresponds to 20 g fresh weight of moss. The vacuum filtrated protonema was chopped in organelle isolation buffer [1% (w/v) polyvinylpolypyrrolidone (PVPP), 300 mM d-sorbitol, 50 mM HEPES, 2 mM Na-EDTA, 1 mM MgCl2 and 0.1% BSA] in the presence of a protease inhibitor [0.1% (v/v) Sigma Plant Protease Inhibitor Cocktail P 9599] using a household vegetable chopping device. After 100–150 strokes with this device the chopped moss was filtered through 3 layers of Miracloth (Calbiochem). The filtrate was transferred to a 50 ml centrifuge tube (Oak Ridge, Nalgene) and the chloroplasts were pelleted at 1,500×g for 10 min (Beckman Coulter Avanti Centrifuge J-25, fixed angle rotor Ja 25.50). The supernatant was decanted into new centrifuge tubes and used for the isolation of mitochondria (see below).
A workflow diagram outlining the isolation protocol is given in Fig. 1.
Fig. 1.
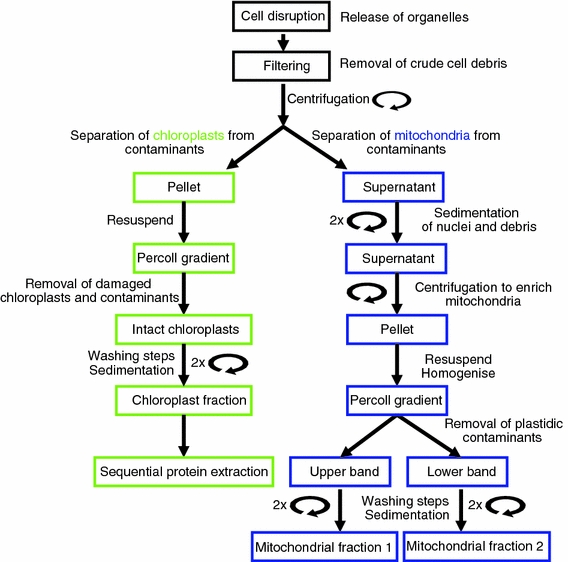
Workflow of the isolation of chloroplasts and mitochondria. Schematic illustration of the different steps during the simultaneous isolation of chloroplasts and mitochondria from moss protonema. After cell disruption, filtration and the first centrifugation step, intact and highly enriched chloroplasts and mitochondria can be isolated from the pellet and the supernatant, respectively, using further differential centrifugation and Percoll density gradients
Chloroplast isolation
The crude chloroplast pellet was resuspended in 3 ml resuspension buffer (300 mM d-sorbitol, 50 mM HEPES, 2 mM Na-EDTA, 1 mM MgCl2 and 0.1% BSA) using a fine paint brush. The resuspended sample was carefully loaded onto several Percoll density gradients (top to bottom: 5 ml 10% Percoll, 5 ml 40% Percoll, 5 ml 80% Percoll) prepared in resuspension buffer. The Percoll gradients were centrifuged for 20 min at 16,000×g. Broken chloroplasts accumulated at the 10–40% interface, while intact chloroplasts formed a band at the 40–80% interface. The fractions containing intact chloroplast were collected and combined using a thin glass pipette. The chloroplasts were washed twice with three volumes of washing buffer (300 mM d-sorbitol, 50 mM HEPES, 2 mM Na-EDTA, 1 mM MgCl2) and centrifuged for 10 min at 1,500×g. Organelle purity was monitored via fluorescence microscopy. Pellets were frozen at −80°C.
Mitochondria isolation
Mitochondria were purified from the 1,500×g supernatant adapting a protocol of Kabeya and Sato (2005) using differential centrifugation and a Percoll density gradient. The supernatant was centrifuged at 3,000×g for 5 min followed by 6,000×g for 5 min in the same tube in order to pellet nuclei and cellular debris. Afterwards the resulting supernatant was decanted to a new tube and centrifuged at 18,000×g for 20 min yielding a greenish pellet enriched in mitochondria.
The pellet was carefully resuspended in 1 ml of washing buffer using a fine paint brush. Several pellets were combined in a potter (Braun, Melsungen, Germany) to a final volume of 4.8 ml and homogenised with 10 strokes. Subsequently 100% Percoll was added to the sample to a final concentration of 20%.
A stepwise Percoll density gradient was assembled by overlaying 5 ml of 80% Percoll in washing buffer with 5 ml of 33% Percoll in washing buffer. The homogenised sample in 20% Percoll was transferred on top of this gradient and centrifuged at 18,000×g for 1 h.
Two pale whitish mitochondrial bands were extracted at the 20–33% interface and the 33–80% interface, respectively. They are subsequently referred to as bands M1 and M2. Mitochondria were carefully collected using thin glass pipettes and freed of Percoll by adding 15 ml of washing buffer, gentle mixing and centrifugation at 18,000×g for 20 min (slow deceleration), repeated twice. The upper band yielded a still greenish, loose mitochondria pellet whereas the lower band formed a stable whitish pellet also containing residual PVPP. Pellets were frozen at −80°C.
Protein extraction
Proteins were extracted from frozen organelle pellets and from frozen protonema using lysis buffer (7.5 M urea, 2.5 M thiourea, 12.5% glycerol, 62.5 mM tris–HCl, 2.5% n-octylglycosid, 1.25 mM protease inhibitor) and subsequently precipitated with methanol/chloroform (modified after Wessel and Flügge 1984). The protein pellet was resuspended in urea buffer (6 M urea, 25 mM bis–tris, 0.2% n-octylglucopyranosid, pH 8.5), measured with a nanodrop (Thermo Scientific) and mixed with SDS sample buffer (Bio-Rad) for SDS-PAGE. The measured protein concentration was confirmed by Coomassie (Fermentas PageBlue) staining of SDS gels. Supplementary Fig. 1 shows an example of a silver-stained SDS-PAGE equally loaded with 4 μg organellar protein extracts prepared using lysis buffer.
Sequential chloroplast protein extraction was performed after a protocol adapted from Molloy et al. (1998) to obtain a soluble fraction (Cs) and a chloroplast membrane fraction (Cm). Frozen chloroplast pellets were resuspended in up to 500 μl chloroplast protein extraction buffer I [CPE I; 2.5 mM EDTA, 5 mM MgCl2, 10 mM KCl, 20 mM HEPES, 300 mM d-sorbitol, 1% (w/v) PVPP, 0.1% (v/v) plant protease inhibitor] followed by subsequent incubation in an ultrasonic ice bath for 20 min. The supernatant was collected after centrifugation (20 min, 4°C, 20,000×g) representing fraction Cs. The residual green pellet was resuspended in up to 500 μl CPE II [CPE I with 1% (v/v) Triton-X 100], followed by a subsequent incubation in an ultrasonic ice bath for 20 min. The supernatant was collected, representing fraction Cm.
The soluble total protein fraction (Ts) was prepared using Tris-buffer [40 mM tris-(hydroxymethyl)-aminomethane, 0.5% (w/v) PVPP, 1% (v/v) plant protease inhibitor]. Frozen protonema (500 mg) was homogenised with mortar and pestle and thawed in the presence of Tris-buffer. The homogenate was incubated for 20 min in an ultrasonic ice bath. Subsequently the homogenate was centrifuged for 90 min at 121,000×g (Beckman Coulter Optima L-60, rotor type Ti 70.1) at 2°C. The clear supernatant was transferred into Teflon-centrifuge tubes (Oak-Ridge, Nalgene) and precipitated overnight with five volumes of acetone with 0.2% dithiothreitol (DTT) at −20°C. Afterwards the proteins were pelleted at 12,000×g for 10 min and 0°C. The protein pellet was resuspended in 20 ml acetone with 0.2% DTT and incubated for 1 h at −20°C. After centrifugation as described above, the air-dried protein was resuspended in lysis buffer [8 M urea, 4% 3((3-cholamidopropyl)-dimethylammonio)-propansulfat, 100 mM DTT, 40 mM Tris-(hydroxylmethyl)-aminomethane]. Protein concentration of the fractions was determined via Bradford assay (Bradford 1976) and mixed with SDS sample buffer (Bio-Rad) for SDS-PAGE.
Western blotting
After gel electrophoresis using ready-made SDS-gels (Ready gels, 12% Tris–HCl, Bio-Rad), the stacking gel was cut off and discarded. The remaining gel was immersed in 100 ml cathode-buffer [25 mM tris, 40 mM glycine, 10% (v/v) methanol] for 15 min. Transfer membranes (Immobilon-P, Millipore) were incubated in 100% methanol for 15 s, washed for 2 min in water and subsequently equilibrated for at least 5 min in 100 ml anode-buffer II [25 mM Tris, 10% (v/v) methanol]. Six pieces of filter paper (Grade GB003, Whatman) in the size of the membranes were incubated as follows: three in 100 ml cathode-buffer, two in 100 ml anode-buffer I [0.3 M Tris, 10% (v/v) methanol] and one together with the membrane in 100 ml anode-buffer II (filter paper on top and membrane below the paper).
The transfer sandwich was placed and assembled directly on the anode-plate of the semi-dry blotting device (Trans-Blot SD Semi-dry transfer cell, Bio-Rad). Assembly of the transfer sandwich from anode to cathode: two filter papers with anode-buffer I, one filter paper with anode-buffer II, membrane, separating gel and three filter papers with cathode-buffer. Transfer was performed at 1.2 mA/cm2 for an hour.
Membranes were blocked with 4% blocking reagent (ECL Advance, GE Healthcare) in TBST [20 mM Tris–HCl, pH 7.6, 137 mM NaCl, 0.1% (v/v) Tween 20] for an hour. Membranes were washed five times: two short immersions, then a 15 min wash followed by two 5 min washing steps. Incubation with primary antibodies was performed at room temperature for an hour followed by the previously described washing procedure. The final concentration for the primary antibody was determined empirically for each antibody (see Table 1 for details). Membranes were incubated with the secondary antibody (anti-rabbit HRP-conjugate, Amersham) at a final concentration of 1:500,000 followed by washing steps. Reactive bands were detected using ECL detection (ECL Advance detection kit, GE Healthcare).
Table 1.
Antibody and protein data
| Antibody name/product numbera | Protein | Expected subcellular localisation | Protein used to raise antibodyb | Organism | Expected/apparent MW (kDa) | Apparent MW in P. patens (kDa) | Dilution of primary antibody used for ECL detection |
|---|---|---|---|---|---|---|---|
| CPX1/AS06 123 | Coproporphyrinogen III oxidase, isoform 1 | Chloroplast | Q9S7V1 | C. reinhardtii | 41.4/38 | 38 | 1:2,000 |
| CRD1/AS06 122 | Cyanobacterial homolog of plant CHL27 cyclase | Chloroplast thylakoid and envelope membranes | Q9M591 | A. thaliana | 47/40 | 48 | 1:5,000 |
| CSP41b/AS08 298 | Ribosome associated endonuclease (CRB) | Chloroplast ribosome | Q9SA52 | A. thaliana | 42/39 | 42 | 1:5,000 |
| Cyt f/AS06 119 | Cytochrome f protein (PetA) of thylakoid Cyt b6/f-complex | Chloroplast thylakoid membrane | P23577 | C. reinhardtii | 34/31–32 | 34 | 1:100,000 |
| HSP70B/AS06 175 | Stromal α-HSP70 | Chloroplast | C. reinhardtii | 71.9 | 72 | 1:10,000 | |
| Lhcb2/AS01 003 | LHCII type II chlorophyll a/b-binding protein | Chloroplast; thylakoid membrane | At2g05100 At2g05070 At3g27690 AF134124 | A. thaliana | 25 | 26 | 1:500,000 |
| PsaD/AS09 461 | PSI reaction centre subunit II | Chloroplast; thylakoid membrane | At1g03130 At4g02770 | A. thaliana | 17.9 | 18/20 | 1:5,000 |
| PsbP/AS06 167 | 23 kDa protein of the oxygen evolving complex (OEC) of PSII | Chloroplast; thylakoid membrane | Q42029 | A. thaliana | 28/23 | 20/23 | 1:10,000 |
| GLN1/GLN2/AS08 295 | Glutamine synthetase | GLN1: cytoplasm | GLN1-1 GLN1-2 | A. thaliana | 39–40 (GLN1) | 42–43 | 1:12,500 |
| GLN2: chloroplast | GLN1-3 GLN1-4 | 44–45 (GLN2) | |||||
| Arf1/AS08 325 | ADP-ribosylation factor 1 | Golgi apparatus | At2g47170; P36397 | A. thaliana | 21 | 19 | 1:4,000 |
| AOX1/2/AS04 054 | Plant alternative oxidase 1 and 2 | Mitochondria inner membrane | At3g22370 At5g64210 | A. thaliana | 36–40 | 35 | 1:1,000 |
| COX II/AS04 053A | Cytochrome c oxidase subunit 2 | Mitochondria inner membrane | AtmG00160 | A. thaliana | 29.4 | 31 | 1:1,000 |
| P04373 | O. sativa | ||||||
| Q1XGA9 | P. patens | ||||||
| VDAC1/AS07 212 | Voltage-dependent anion-selective channel protein 1 | Mitochondria outer membrane | At3g01280 | A. thaliana | 29 | 31 | 1:5,000 |
| V-type ATPase/AS07 213 | Epsilon subunit of tonoplast H+ ATPase | Tonoplast membrane | At4g11150 | A. thaliana | 26 | 25 | 1:2,000 |
Summarised information on antibody names, full name of their target protein and predicted protein localisations. For the proteins used to raise the antibodies, UniProt or TAIR accession numbers, organism of origin and expected/apparent molecular weights are given. For the Western blots shown in this study, the apparent molecular weights in P. patens and the antibody dilutions used are depicted
aAgrisera product number (http://www.agrisera.com)
bProtein accession number, source: http://www.ncbi.nlm.nih.gov/genbank/
Antibody production
Polyclonal antibodies were raised in rabbits against KLH-conjugated peptides derived from conserved regions of the target protein across a range of species including mono- and dicotyledonous plants and P. patens or Chlamydomonas reinhardtii. The antibodies were validated using available positive and negative controls. A summary of antibody names, full names of their target proteins, predicted protein localisations, UniProt or TAIR accession numbers, organism of origin and expected/apparent molecular weights is given in Table 1.
Fluorescence microscopy
To stain mitochondria and verify mitochondrial integrity 200 μl of mitochondrial fractions (M1 and M2) were taken from the density gradient and stained with 200 nM MitoTracker Green FM and MitoTracker Red CMXRos (Molecular Probes), respectively, in washing buffer. After 15 min of gentle shaking at room temperature, samples were observed under a fluorescence microscope (Axioplan, Zeiss, Jena, Germany) using a bandpass 450–490 nm excitation filter with a 520 nm long-pass emission filter for MitoTracker Green FM stained samples and a bandpass 546/12 excitation filter with a 590 long-pass emission filter for MitoTracker Red CMXRos stained samples.
Results
Protein yield
Simple chopping of moss protonema yielded a cell homogenate that can be easily and rapidly fractionated for the simultaneous isolation of chloroplasts and mitochondria via differential centrifugation and Percoll density gradients (Fig. 2a, b). Twenty gram fresh weight of moss material (corresponding to 2 g dry weight) yielded protein amounts of 3–5 mg chloroplast protein and 60–100 μg mitochondrial protein, i.e. 150–250 μg of chloroplast protein and 3–5 μg mitochondrial protein per gram of fresh weight of moss material. After isolation, organelle pellets were frozen and the proteins from the respective pellets extracted for subsequent experiments.
Fig. 2.

Percoll gradients after centrifugation. Percentages of Percoll in buffer are given for the different layers. a Purification of intact chloroplasts, D debris, C intact chloroplasts b Purification of mitochondria, M1 mitochondrial fraction 1, M2 mitochondrial fraction 2; the greenish band in the upper part of the gradient consists of chloroplast debris
Purity and integrity of mitochondria
Two distinct whitish bands were observed in the Percoll density gradient during purification of mitochondria, one at the 20–33% interface (M1) just below the green band of chloroplast debris and another one at the 33–80% interface (M2) (Fig. 2b). Samples from both bands were analysed by fluorescence microscopy and the presence of mitochondria was confirmed by MitoTracker Green staining (Supplementary Fig. 2a, b). Additional staining with MitoTracker Red CMXRos confirmed the existence of a membrane potential (Supplementary Fig. 2c, d).
Impurities in the mitochondrial fraction were assessed using antibodies against plastidic contaminants, namely, the membrane localised light-harvesting complex II type II chlorophyll a/b-binding protein (Lhcb2) and the plastidic stroma localised glutamine synthetase (GLN1/GLN2), and antibodies against Golgi apparatus and tonoplast membrane contaminants, namely, the ADP-ribosylation factor 1 (Arf1) and the Epsilon subunit of the tonoplast H+ ATPase (V-type ATPase) localised in Golgi and tonoplast, respectively (Fig. 3).
Fig. 3.
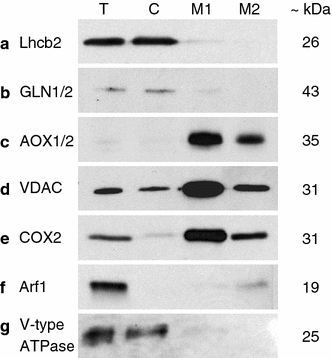
Immunoblots assessing the organellar purity of urea-extracted proteins. T total protein, C chloroplast protein, M1 mitochondrial fraction 1 protein, M2 mitochondrial fraction 2 protein, loading amounts: 10 μg/lane (a, e), 2.5 μg/lane (b–d, f, g); for dilutions of primary antibodies, see Table 1
Immunoblot signals for the mitochondrial fraction from the 20–33% interface (M1) were weak for plastidic markers and absent for Golgi vesicle and tonoplast markers (Fig. 3a, b, f, g), while the mitochondrial fraction from the 33–80% interface (M2) showed only a weak positive signal for contamination with Golgi vesicles (Fig. 3f).
The signal strength for the three mitochondrial marker antibodies used, the plant alternative oxidase 1 and 2 (AOX1/2) and the cytochrome c oxidase subunit 2 (COX II) which are both localised in the inner mitochondrial membrane, and the antibody against the voltage-dependent anion-selective channel protein 1 (VDAC1), a protein of the outer mitochondrial membrane, was higher for the mitochondrial protein extracts from the M1 fraction (Fig. 3c, d). Integrity of the mitochondria of both fractions was confirmed by the positive immunoblot signal for VDAC1 (Fig. 3d).
Purity of chloroplasts
During the purification in the Percoll density gradient, the intact chloroplasts accumulated at the 40–80% interface, while the broken chloroplasts were found at the 10–40% interface (Fig. 2a).
To detect impurities in the chloroplast fraction (C) the same set of antibodies as for the mitochondrial fraction was used (Fig. 3). Immunoblots showed that the chloroplasts were free of contamination by Golgi (Fig. 3f) but were contaminated to some extent with tonoplast membranes (Fig. 3g). The signal strength of the mitochondrial marker antibodies against chloroplast proteins varied strongly. The AOX1/2 antibody gave no signal whereas the signal for COX II was weak and the one for VDAC1 was clear (Fig. 3c–e). The immunoblot signals were all at the expected size as indicated by the manufacturer. The total protein extract (T) used as a control showed clear signals for each of the antibodies tested besides from the AOX1/2, which was very weak (Fig. 3a–g).
The success of the organelle isolation procedure was further assessed for chloroplast proteins via the application of a series of chloroplast specific antibodies against the sequentially extracted soluble (Cs) and membrane proteins (Cm) from intact chloroplasts (Fig. 4).
Fig. 4.
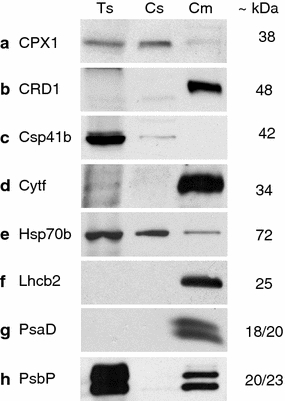
Immunoblots from sequentially extracted proteins of chloroplasts. Ts total soluble protein, Cs chloroplast soluble protein, Cm chloroplast membrane protein; loading amounts: 5 μg/lane; for dilutions of primary antibodies, see Table 1
All the antibodies against chloroplast proteins not localised to the membranes, i.e. the coproporphyrinogen III oxidase, isoform 1 (CPX1), the ribosome associated endonuclease (CSP41b) and the stromal heat-shock protein (Hsp70b), showed a clear signal in the soluble chloroplast protein fraction whereas for the proteins extracted from the membrane fraction only the CPX1 and the Hsp70b resulted in a very weak and a weak signal, respectively (Fig. 4a, c, e).
Immunoblot signals for the antibodies against proteins localised to the chloroplast membranes, namely, the cyanobacterial homolog of the plant CHL27 cyclase (CRD1), the cytochrome f protein of the thylakoid Cyt b6/f-complex (Cyt f), the light-harvesting complex II type II chlorophyll a/b-binding protein (Lhcb2), the photosystem I reaction centre subunit II (PsaD) and the 23 kDa protein of the oxygen evolving complex of the photosystem II (PsbP) were all very strong in the protein extract from the chloroplast membrane fraction (Fig. 4b, d, f–h). For both antibodies, PsaD and PsbP, two signals were obtained, namely, at 18 and 20 kDa for PsaD and at 20 and 23 kDa for PsbP. None of these five antibodies against membrane proteins of the chloroplast delivered a signal in the protein extracts of the chloroplast soluble fraction. For the protein extracts from the soluble total protein sample (Ts) immunoblot signals were obtained for all the proteins not localised to the membranes (CPX1, CSP41b, Hsp70b) (Fig. 4a, c, e). No signal was obtained for the membrane localised chloroplast proteins, besides for PsbP with which two highly saturated signals were recorded (Fig. 4h).
Discussion
Protein yield
Previous publications using mitochondria for proteomic studies (Heazlewood et al. 2004; Kruft et al. 2001; Millar et al. 2001a, b) did not indicate the amount of mitochondrial protein recovered per gram of fresh weight of starting material. It is certain that in moss a reduced amount of starting material is available as compared to seed plants. However, the optimised cultivation of moss protonema in aerated liquid culture allows for a yield of 60–100 μg mitochondrial protein per isolation. The protein yield from isolated chloroplasts is well in the range of previously reported studies. While Kley et al. (2010) reported a range of 149–499 μg Arabidopsis chloroplast proteins per gram of rosette leaves, Salvi et al. (2008a) recovered about 100–150 μg protein per gram using 400–500 g of leaf material. Previous protocols for organelle extraction in moss (Kabeya and Sato, 2005; Marienfeld et al. 1989) did not report protein yield as reference value. Taking into account that the moss material used in this study is derived from a liquid protonema culture, the amount of 20 g fresh weight of moss and the protein yield extracted from this material are very acceptable to routinely isolate both organellar fractions and perform subsequent proteomics analyses.
Purity and integrity of mitochondria
Surprisingly, the isolation protocol for mitochondria reported here revealed two bands of different densities, indicating two populations of mitochondria in moss protonema. In contrast, Kabeya and Sato (2005) reported only one single band at the 33–80% Percoll interface. It is possible that the homogenisation step using a potter before the density gradient purification causes disruption of some mitochondria. However, the clear signal in the immunoblot obtained for the mitochondrial outer membrane protein VDAC1 as well as the staining with Mitotracker Red CMXRos, which stains only intact mitochondria with membrane potential (Gilmore and Wilson 1999; Metivier et al. 1998; Poot et al. 1996) indicate that both mitochondrial fractions contain intact mitochondria. So far, very few reports of a second mitochondrial band in a density gradient exist (Logan et al. 2001; Segui-Simarro et al. 2008). While Logan et al. (2001) found two subpopulations of mitochondria differing in the degree of cristae organisation using electron microscopy techniques on germinating maize embryos, Segui-Simarro et al. (2008) reported for A. thaliana that in meristematic cells two subpopulations of mitochondria coexist. The mitochondria extracted from maize were obtained using a sucrose gradient where the organelles accumulated at a density of 22–28% sucrose and at a density of 37–42%, the latter “heavy” band being the described and expected one. These authors concluded that the mitochondria accumulating in the “light” fraction were remains of mitochondria that were active in the developing embryo during the maturation phase of seed development (Logan et al. 2001). A comparison of these results with those from P. patens would be far-fetched as there are no such developmental stages in moss. Observations by Segui-Simarro et al. (2008) in Arabidopsis were based on reconstructed thin-sections of electron micrographs and not on isolated mitochondria and, hence, have to be considered as an in planta finding. The two different mitochondrial fractions in moss as presented here are isolated from protonema without previous protoplastation, in contrast to the protocol of Kabeya and Sato (2005). Protoplastation may lead to a unification of the cells which in turn could reduce or abolish potential functional or morphological differences between organelles in distinct cell types. To date it is unclear if these two populations of mitochondria originate from mitochondria present in different cell types (as e.g. from caulonema and chloronema cells within the protonema tissue) or from two subpopulations of mitochondria from the same set of cells.
The results from the immunoblot analysis demonstrate that the plastidic contamination of both mitochondrial fractions is minimal or not existent, since the signal from the very abundant light-harvesting complex protein (Lhcb2) is absent in the lower mitochondrial fraction M2 and very weak in the upper mitochondrial band M1. In addition, this difference might be caused by residual contamination due to the green layer of broken cell debris accumulating above the M1 band in the density gradient.
The pronounced increase in signal intensity for the three mitochondrial markers in both mitochondrial protein fractions as compared to the total protein and chloroplast protein extracts, which is best observed for the AOX1/2 immunoblot results, confirms that the protocol besides yielding intact and pure mitochondria delivers a highly enriched fraction of organelles. Interestingly, the signal for all mitochondrial markers used is stronger in the newly described M1 fraction which hints to differences in protein content between the mitochondrial fractions.
Potential contaminations of the M2 fraction by Golgi and a closer characterisation of both mitochondrial bands at the level of proteins will have to be analysed in subsequent proteomics studies.
Purity of chloroplasts
Comparable to Mason et al. (1991), the intact chloroplast fraction isolated from moss protonema accumulated at the 40–80% Percoll interface. Immunoblot results for the proteins extracted from this fraction suggest that the contamination of the chloroplast fraction with outer mitochondrial membranes represented by the VDAC protein is higher than with inner mitochondrial membrane proteins such as COX II and AOX1/2. The contamination with outer mitochondrial membranes might be caused by an attachment of membrane pieces deriving from broken mitochondria to intact chloroplasts during the isolation. A slight contamination due to the handling procedure cannot be ruled out as the removal of the intact chloroplasts from the density gradient with a Pasteur pipette requires penetrating the broken cell debris layer, which might automatically lead to a residual contamination of the glass pipette surface.
Immunoblotting results with the urea-extracted proteins using the plant compartment marker antibodies indicate that there is a contamination with mitochondria in the chloroplast fraction and vice versa. However, contaminations by the Golgi apparatus are absent while there is a clear signal from contaminants of the tonoplast.
The use of several different chloroplast protein markers clearly demonstrates that the method for extraction of a soluble and a membrane chloroplast protein fraction worked very well. All the proteins reported to be part of the chloroplast stroma resulted in clear immunoblot signals for the soluble protein fraction, whereas none of the membrane-localised chloroplast proteins gave a signal. On the other hand, the proteins extracted from the chloroplast membrane fraction resulted in a clear and strong immunoblot signal. The immunoreactive bands observed for CPX1 and Hsp70b in the chloroplast membrane protein fraction might be the result of contaminations due to handling of samples since during sequential chloroplast protein extraction it is not improbable that residues of the soluble fraction are still present in samples of membrane protein. Therefore, even though a residual contamination of soluble proteins in the membrane fraction is likely, there is no contamination of the soluble fraction with membrane proteins. Our protocol might therefore be used in subsequent studies to specifically analyse the proteins present in the chloroplast stroma and to investigate the proteins of a highly enriched thylakoid membrane protein fraction.
Specific enrichment using the sequential extraction method could be confirmed via the immunoblot results with total soluble proteins. None of the chloroplast membrane protein markers resulted in a signal for the total soluble proteins besides for the membrane localised PsbP.
Interestingly, the anti-PsbP as well as the anti-PsaD antibodies gave two signals in the immunoblot. The expected unique 23 kDa signal for A. thaliana matched with the apparent size of one of the bands for the P. patens chloroplast membrane protein extract, while the second minor band at 20 kDa had not been expected. Yi et al. (2007) also reported two signals for the PsbP protein from A. thaliana, which is a highly conserved protein in seed plants (Bricker and Burnap 2005; Roose et al. 2007). Although recent findings in A. thaliana have demonstrated that there are two putative genes that encode for PsbP-1 and PsbP-2, respectively, the minor band does not correspond to the PsbP-2 protein because it migrates at significantly lower apparent molecular mass (Goulas et al. 2006).
Similarly, although it has been found in other species such as A. thaliana that there can be a second immunoreactive band at 20 kDa, i.e. a slightly higher band than the expected unique 18 kDa band using the PsaD antibody, the origin of this second signal is unclear. The question whether the two signals found here with moss represent processed or modified forms of the proteins remains elusive and might be answered using sequencing techniques.
General remarks and conclusions
Our protocol for a simultaneous isolation of chloroplasts and mitochondria from the same biological sample offers several advantages as it will facilitate a direct comparison of organelles under identical physiological conditions. Protonema tissue has only few different cell types (Reski 1998) and thus is a rather homogeneous material, well suited for systems biology approaches (Decker et al. 2006). In addition, the rapid protocol using intact tissue rather than protoplasts described here, will promote such studies in P. patens. Furthermore, employing this protocol we found two distinct mitochondrial bands, a finding that has not been reported before for moss and will be investigated further.
The use of plant compartment marker antibodies designed for algae or seed-plants in this study supports two main points. On the one hand, it demonstrates that plant antibodies can be used across species if their target sequences are conserved and, on the other hand, it has shown for this study that both organellar fractions are highly enriched with only minor contaminations by other organelles. Both aspects paired with the good protein yields obtained from the extractions will allow the analysis of sub-cellular proteomes of this non-flowering plant, thus helping to increase our understanding of the evolution of metabolic compartmentalisation, of biosynthetic pathways and of protein sorting mechanisms in land plants.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Silver staining of a SDS-PAGE (12%) loaded with 4 μg of protein extracts; L: 5 μl of Fermentas Prestained PageRuler™ protein ladder, T: total protein, C: chloroplast protein, M2: mitochondrial fraction 2 protein, M1: mitochondrial fraction 1 protein. Protein extracts from organelle fractions contain residual BSA (~66 kDa) present in the isolation buffer (TIFF 201 kb)
Fluorescence microscopy images of stained mitochondria. 200 μl of mitochondrial fractions M1 (A,C) and M2 (B,D) were stained with MitoTracker Green FM (A,B) or MitoTracker Red CMXRos and observed under a fluorescence microscope (Axioplan, Zeiss, Jena, Germany). Scale bar: 50 μm (TIFF 533 kb)
Acknowledgments
This work was supported by the Excellence Initiative of the German Federal and State Governments (GSC-4, Spemann Graduate School of Biology and Medicine), by the German Federal Ministry of Education and Research BMBF (0313921-FRISYS) and by the MINT-Initiative of the State of Baden-Württemberg. We thank Christine Glockner for technical assistance.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Abbreviations
- AOX
Alternative oxidase
- Arf
ADP-ribosylation factor
- Csp41b
Ribosome associated endonuclease
- COX
Cytochrome c oxidase
- CPX1
Coproporphyrinogen III oxidase
- CRD1
Cyanobacterial homolog of plant CHL27 cyclase
- Cyt f
Cytochrome f protein of thylakoid cytochrome b6/f-complex
- GLN
Glutamine synthetase
- Hsp70b
Stromal alfa-heat shock protein 70
- Lhcb2
Light harvesting complex II chlorophyll a/b-binding protein
- PsaD
Photosystem I reaction centre subunit II
- PsbP
23 kDa protein of the oxygen evolving complex of photosystem II
- VDAC
Voltage dependent anion channel
- V-type ATPase
Vacuolar-type ATPase
Footnotes
E. G. E. Lang and S. J. Mueller contributed equally to this work.
References
- Baginski S, Gruissem W. Chloroplast proteomics: potentials and challenges. J Exp Bot. 2004;55:1213–1220. doi: 10.1093/jxb/erh104. [DOI] [PubMed] [Google Scholar]
- Baginski S, Kleffmann T, von Zychlinski A, Gruissem W. Analysis of shotgun proteomics and RNA profiling data from Arabidopsis thaliana chloroplasts. J Prot Res. 2005;4:637–640. doi: 10.1021/pr049764u. [DOI] [PubMed] [Google Scholar]
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. doi: 10.1016/0003-2697(76)90527-3. [DOI] [PubMed] [Google Scholar]
- Bricker T, Burnap R. The extrinsic proteins of photosystem II. In: Wydrzynski TJ, Kimiyuki S, editors. Photosystem II: the light-driven water: plastoquinone oxidoreductase. Dordrecht: Springer; 2005. pp. 95–120. [Google Scholar]
- Bruce BD. The paradox of plastid transit peptides: conservation of function despite divergence in primary structure. Biochim Biophys Acta. 2001;1541:2–21. doi: 10.1016/S0167-4889(01)00149-5. [DOI] [PubMed] [Google Scholar]
- Decker EL, Frank W, Sarnighausen E, Reski R. Moss systems biology en route: phytohormones in Physcomitrella development. Plant Biol. 2006;8:397–405. doi: 10.1055/s-2006-923952. [DOI] [PubMed] [Google Scholar]
- Eubel H, Lee CP, Kuo J, Meyer EH, Taylor NL, Millar AH. Free-flow electrophoresis for purification of plant mitochondria by surface charge. Plant J. 2007;52:583–594. doi: 10.1111/j.1365-313X.2007.03253.x. [DOI] [PubMed] [Google Scholar]
- Frank W, Baar KM, Qudeimat E, Woriedh M, Alawady A, Ratnadewi D, Gremillon L, Grimm B, Reski R. A mitochondrial protein homologous to the mammalian peripheral-type benzodiazepine receptor is essential for stress adaptation in plants. Plant J. 2007;51:1004–1018. doi: 10.1111/j.1365-313X.2007.03198.x. [DOI] [PubMed] [Google Scholar]
- Gilmore K, Wilson M. The use of chloromethyl-X-rosamine (Mitotracker red) to measure loss of mitochondrial membrane potential in apoptotic cells is incompatible with cell fixation. Cytometry. 1999;36:355–358. doi: 10.1002/(SICI)1097-0320(19990801)36:4<355::AID-CYTO11>3.0.CO;2-9. [DOI] [PubMed] [Google Scholar]
- Goulas E, Schubert M, Kieselbach T, Kleczkowski LA, Gardestrom P, Schroder W, Hurry V. The chloroplast lumen and stromal proteomes of Arabidopsis thaliana show differential sensitivity to short- and long-term exposure to low temperature. Plant J. 2006;47:720–734. doi: 10.1111/j.1365-313X.2006.02821.x. [DOI] [PubMed] [Google Scholar]
- Gray MW, Burger G, Lang BF. Mitochondrial evolution. Science. 1999;283:1476–1481. doi: 10.1126/science.283.5407.1476. [DOI] [PubMed] [Google Scholar]
- Heazlewood JL, Howell KA, Whelan J, Millar AH. Towards an analysis of the rice mitochondrial proteome. Plant Physiol. 2003;132:230–242. doi: 10.1104/pp.102.018986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heazlewood JL, Tonti-Filippini JS, Gout AM, Day DA, Whelan J, Millar AH. Experimental analysis of the Arabidopsis mitochondrial proteome highlights signaling and regulatory components, provides assessment of targeting prediction programs, and indicates plant-specific mitochondrial proteins. Plant Cell. 2004;16:241–256. doi: 10.1105/tpc.016055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Heintz D, Wurtz V, High AA, Van Dorsselaer A, Reski R, Sarnighausen E. An efficient protocol for the identification of protein phosphorylation in a seedless plant, sensitive enough to detect members of signalling cascades. Electrophoresis. 2004;25:1149–1159. doi: 10.1002/elps.200305795. [DOI] [PubMed] [Google Scholar]
- Heintz D, Erxleben A, High AA, Wurtz V, Reski R, Van Dorsselaer A, Sarnighausen E. Rapid alteration of the phosphoproteome in the moss Physcomitrella patens after cytokinin treatment. J Proteome Res. 2006;5:2283–2293. doi: 10.1021/pr060152e. [DOI] [PubMed] [Google Scholar]
- Huang S, Taylor NL, Narsai R, Eubel H, Whelan J, Millar AH. Experimental analysis of the rice mitochondrial proteome, its biogenesis, and heterogeneity. Plant Physiol. 2009;149:719–734. doi: 10.1104/pp.108.131300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kabeya Y, Sato N. Unique translation initiation at the second AUG codon determines mitochondrial localization of the phage-type RNA polymerases in the moss Physcomitrella patens. Plant Physiol. 2005;138:369–382. doi: 10.1104/pp.105.059501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kasten B, Buck F, Nuske J, Reski R. Cytokinin affects nuclear- and plastome-encoded energy-converting plastid enzymes. Planta. 1997;201:261–272. doi: 10.1007/s004250050065. [DOI] [PubMed] [Google Scholar]
- Khandelwal A, Cho SH, Marella H, Sakata Y, Perroud PF, Pan A, Quatrano RS. Role of ABA and ABI3 in desiccation tolerance. Science. 2010;327:546. doi: 10.1126/science.1183672. [DOI] [PubMed] [Google Scholar]
- Kiessling J, Martin A, Gremillon L, Rensing SA, Nick P, Sarnighausen E, Decker EL, Reski R. Dual targeting of plastid division protein FtsZ to chloroplasts and the cytoplasm. EMBO Rep. 2004;5:889–894. doi: 10.1038/sj.embor.7400238. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kleffmann T, Russenberger D, von Zychlinski A, Christopher W, Sjolander K, Gruissem W, Baginsky S. The Arabidopsis thaliana chloroplast proteome reveals pathway abundance and novel protein functions. Curr Biol. 2004;14:354–362. doi: 10.1016/j.cub.2004.02.039. [DOI] [PubMed] [Google Scholar]
- Kley J, Heil M, Muck A, Svatos A, Boland W. Isolating intact chloroplasts from small Arabidopsis samples for proteomic studies. Anal Biochem. 2010;398:198–202. doi: 10.1016/j.ab.2009.11.016. [DOI] [PubMed] [Google Scholar]
- Kopriva S, Fritzemeier K, Wiedemann G, Reski R. The putative moss 3′phosphoadenosine 5′phosphosulfate reductase is a novel form of adenosine 5′phosphosulfate reductase without an iron sulfur cluster. J Biol Chem. 2007;282:22930–22938. doi: 10.1074/jbc.M702522200. [DOI] [PubMed] [Google Scholar]
- Kruft V, Eubel H, Jansch L, Werhahn W, Braun HP. Proteomic approach to identify novel mitochondrial proteins in Arabidopsis. Plant Physiol. 2001;127:1694–1710. doi: 10.1104/pp.010474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lee CP, Eubel H, O’Toole N, Millar AH. Heterogeneity of the mitochondrial proteome for photosynthetic and non-photosynthetic Arabidopsis metabolism. Mol Cell Proteomics. 2008;7:1297–1316. doi: 10.1074/mcp.M700535-MCP200. [DOI] [PubMed] [Google Scholar]
- Logan DC, Millar AH, Sweetlove LJ, Hill SA, Leaver CJ. Mitochondrial biogenesis during germination in maize embryos. Plant Physiol. 2001;125:662–672. doi: 10.1104/pp.125.2.662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marienfeld JR, Reski R, Friese C, Abel WO. Isolation of nuclear, chloroplast and mitochondrial DNA from the moss Physcomitrella patens. Plant Sci. 1989;61:235–244. doi: 10.1016/0168-9452(89)90230-6. [DOI] [Google Scholar]
- Mason CB, Matthews S, Bricker TM, Moroney JV. Simplified procedure for the isolation of intact chloroplasts from Chlamydomonas reinhardtii. Plant Physiol. 1991;97:1576–1580. doi: 10.1104/pp.97.4.1576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Metivier D, Dallaporta B, Zamzami N, Larochette N, Susin SA, Marzo I, Kroemer G. Cytofluorometric detection of mitochondrial alterations in early CD95/Fas/APO-1-triggered apoptosis of Jurkat T lymphoma cells. Comparison of seven mitochondrion-specific fluorochromes. Immunol Lett. 1998;61:157–163. doi: 10.1016/S0165-2478(98)00013-3. [DOI] [PubMed] [Google Scholar]
- Millar AH, Liddell A, Leaver CJ. Isolation and subfractionation of mitochondria from plants. Methods Cell Biol. 2001;65:53–74. doi: 10.1016/S0091-679X(01)65004-0. [DOI] [PubMed] [Google Scholar]
- Millar AH, Sweetlove LJ, Giege P, Leaver CJ. Analysis of the Arabidopsis mitochondrial proteome. Plant Physiol. 2001;127:1711–1727. doi: 10.1104/pp.010387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mitschke J, Fuss J, Blum T, Höglund A, Reski R, Kohlbacher O, Rensing SA. Prediction of dual targeting to plant organelles. New Phytol. 2009;183:224–236. doi: 10.1111/j.1469-8137.2009.02832.x. [DOI] [PubMed] [Google Scholar]
- Molloy MP, Herbert BR, Walsh BJ, Tyler MI, Traini M, Sanchez JC, Hochstrasser DF, Williams KL, Gooley AA. Extraction of membrane proteins by differential solubilization for separation using two-dimensional gel electrophoresis. Electrophoresis. 1998;19:837–844. doi: 10.1002/elps.1150190539. [DOI] [PubMed] [Google Scholar]
- Poot M, Zhang YZ, Kramer JA, Wells KS, Jones LJ, Hanzel DK, Lugade AG, Singer VL, Haugland RP. Analysis of mitochondrial morphology and function with novel fixable fluorescent stains. J Histochem Cytochem. 1996;44:1363–1372. doi: 10.1177/44.12.8985128. [DOI] [PubMed] [Google Scholar]
- Rensing SA, Lang D, Zimmer AD, Terry A, Salamov A, Shapiro H, Nishiyama T, Perroud PF, Lindquist EA, Kamisugi Y, Tanahashi T, Sakakibara K, Fujita T, Oishi K, Shin IT, Kuroki Y, Toyoda A, Suzuki Y, Hashimoto S, Yamaguchi K, Sugano S, Kohara Y, Fujiyama A, Anterola A, Aoki S, Ashton N, Barbazuk WB, Barker E, Bennetzen JL, Blankenship R, Cho SH, Dutcher SK, Estelle M, Fawcett JA, Gundlach H, Hanada K, Heyl A, Hicks KA, Hughes J, Lohr M, Mayer K, Melkozernov A, Murata T, Nelson DR, Pils B, Prigge M, Reiss B, Renner T, Rombauts S, Rushton PJ, Sanderfoot A, Schween G, Shiu SH, Stueber K, Theodoulou FL, Tu H, Van de Peer Y, Verrier PJ, Waters E, Wood A, Yang L, Cove D, Cuming AC, Hasebe M, Lucas S, Mishler BD, Reski R, Grigoriev IV, Quatrano RS, Boore JL. The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science. 2008;319:64–69. doi: 10.1126/science.1150646. [DOI] [PubMed] [Google Scholar]
- Reski R. Development, genetics and molecular biology of mosses. Bot Acta. 1998;111:1–15. [Google Scholar]
- Reski R. Challenges to our current view on chloroplasts. Biol Chem. 2009;390:731–738. doi: 10.1515/BC.2009.089. [DOI] [PubMed] [Google Scholar]
- Reski R, Abel WO. Induction of budding on chloronemata and caulonemata of the moss, Physcomitrella patens, using isopentenyladenine. Planta. 1985;165:354–358. doi: 10.1007/BF00392232. [DOI] [PubMed] [Google Scholar]
- Richardt S, Lang D, Reski R, Frank W, Rensing SA. PlanTAPDB, a phylogeny-based resource of plant transcription-associated proteins. Plant Physiol. 2007;143:1452–1466. doi: 10.1104/pp.107.095760. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Richardt S, Timmerhaus G, Lang D, Qudeimat E, Correa LG, Reski R, Rensing SA, Frank W. Microarray analysis of the moss Physcomitrella patens reveals evolutionarily conserved transcriptional regulation of salt stress and abscisic acid signalling. Plant Mol Biol. 2010;72:27–45. doi: 10.1007/s11103-009-9550-6. [DOI] [PubMed] [Google Scholar]
- Richter U, Kiessling J, Hedtke B, Decker E, Reski R, Börner T, Weihe A. Two RpoT genes of Physcomitrella patens encode phage-type RNA polymerases with dual targeting to mitochondria and plastids. Gene. 2002;290:95–105. doi: 10.1016/S0378-1119(02)00583-8. [DOI] [PubMed] [Google Scholar]
- Roose JL, Wegener KM, Pakrasi HB. The extrinsic proteins of photosystem II. Photosynth Res. 2007;92:369–387. doi: 10.1007/s11120-006-9117-1. [DOI] [PubMed] [Google Scholar]
- Salvi D, Rolland N, Joyard J, Ferro M. Purification and proteomic analysis of chloroplasts and their sub-organellar compartments. Methods Mol Biol. 2008;432:19–36. doi: 10.1007/978-1-59745-028-7_2. [DOI] [PubMed] [Google Scholar]
- Salvi D, Rolland N, Joyard J, Ferro M. Assessment of organelle purity using antibodies and specific assays: the example of the chloroplast envelope. Methods Mol Biol. 2008;432:345–356. doi: 10.1007/978-1-59745-028-7_23. [DOI] [PubMed] [Google Scholar]
- Segui-Simarro JM, Coronado MJ, Staehelin LA. The mitochondrial cycle of Arabidopsis shoot apical meristem and leaf primordium meristematic cells is defined by a perinuclear tentaculate/cage-like mitochondrion. Plant Physiol. 2008;148:1380–1393. doi: 10.1104/pp.108.126953. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Strittmatter P, Soll J, Bolter B. The chloroplast protein import machinery: a review. Methods Mol Biol. 2010;619:307–321. doi: 10.1007/978-1-60327-412-8_18. [DOI] [PubMed] [Google Scholar]
- Stumpe M, Bode J, Göbel C, Wichard T, Schaaf A, Frank W, Frank M, Reski R, Pohnert G, Feussner I. Biosynthesis of C9-aldehydes in the moss Physcomitrella patens. Biochim Biophys Acta. 2006;1761:301–312. doi: 10.1016/j.bbalip.2006.03.008. [DOI] [PubMed] [Google Scholar]
- Sweetlove LJ, Taylor NL, Leaver CJ. Isolation of intact, functional mitochondria from the model plant Arabidopsis thaliana. Methods Mol Biol. 2007;372:125–136. doi: 10.1007/978-1-59745-365-3_9. [DOI] [PubMed] [Google Scholar]
- Wessel D, Flügge UI. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal Biochem. 1984;138:141–143. doi: 10.1016/0003-2697(84)90782-6. [DOI] [PubMed] [Google Scholar]
- Wiedemann G, Hermsen C, Melzer M, Büttner-Mainik A, Renneberg H, Reski R, Kopriva S. Targeted knock-out of a gene encoding sulfite reductase in the moss Physcomitrella patens affects gametophytic and sporophytic development. FEBS Lett. 2010;584:2271–2278. doi: 10.1016/j.febslet.2010.03.034. [DOI] [PubMed] [Google Scholar]
- Yi X, Hargett SR, Liu H, Frankel LK, Bricker TM. The PsbP protein is required for photosystem II complex assembly/stability and photoautotrophy in Arabidopsis thaliana. J Biol Chem. 2007;282:24833–24841. doi: 10.1074/jbc.M705011200. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Silver staining of a SDS-PAGE (12%) loaded with 4 μg of protein extracts; L: 5 μl of Fermentas Prestained PageRuler™ protein ladder, T: total protein, C: chloroplast protein, M2: mitochondrial fraction 2 protein, M1: mitochondrial fraction 1 protein. Protein extracts from organelle fractions contain residual BSA (~66 kDa) present in the isolation buffer (TIFF 201 kb)
Fluorescence microscopy images of stained mitochondria. 200 μl of mitochondrial fractions M1 (A,C) and M2 (B,D) were stained with MitoTracker Green FM (A,B) or MitoTracker Red CMXRos and observed under a fluorescence microscope (Axioplan, Zeiss, Jena, Germany). Scale bar: 50 μm (TIFF 533 kb)