


Abstract
Telomerase is a ribonucleoprotein (RNP) particle required for the replication of telomeres. The RNA component, termed hTR, of human telomerase contains a domain structurally and functionally related to box H/ACA small nucleolar RNAs (snoRNAs). Furthermore, hTR is known to be associated with two core components of H/ACA snoRNPs, hGar1p and Dyskerin (the human counterpart of yeast Cbf5p). To assess the functional importance of the association of hTR with H/ACA snoRNP core proteins, we have attempted to express hTR in a genetically tractable system, Saccharomyces cerevisiae. Both mature non-polyadenylated and polyadenylated forms of hTR accumulate in yeast. The former is associated with all yeast H/ACA snoRNP core proteins, unlike TLC1 RNA, the endogenous RNA component of yeast telomerase. We show that the presence of the H/ACA snoRNP proteins Cbf5p, Nhp2p and Nop10p, but not Gar1p, is required for the accumulation of mature non-polyadenylated hTR in yeast, while accumulation of TLC1 RNA is not affected by the absence of any of these proteins. Our results demonstrate that yeast telomerase is unrelated to H/ACA snoRNPs. In addition, they show that the accumulation in yeast of the mature RNA component of human telomerase depends on its association with three of the four core H/ACA snoRNP proteins. It is likely that this is the case in human cells as well.
INTRODUCTION
Telomerase is a ribonucleoprotein (RNP) particle found from yeast to human, that is required for the complete replication of telomeres (1). In all organisms, the RNA component of telomerase contains a small region that is used as a template by the reverse transcriptase subunit of the particle to synthesize telomeric repeats. Until recently, the structure and function of the rest of the RNA component [designated hTR (2)] of human telomerase remained ill-defined. Work by Mitchell et al. (3) and Chen et al. (4) have provided insight into these issues. The former noticed that the 3′ half of the RNA component of mouse and human telomerases can be folded into a structure reminiscent of that adopted by box H/ACA small nucleolar RNAs (snoRNAs) (3). These RNAs are involved in chemical modification and cleavage of the pre-rRNA (5–7). Comparison of the sequences of the RNA component of telomerases from 32 different vertebrate species then led Chen et al. (4) to propose a phylogenetically supported secondary structure model for vertebrate telomerase RNA. This model does include a domain akin to box H/ACA snoRNAs at the 3′-end of the RNA (4). The structure of box H/ACA snoRNAs is characterized by the succession of two irregular stem–loops containing an internal bulge. These stem–loop structures are separated by a single-stranded hinge region containing the conserved H box (consensus 5′-ANANNA-3′) and followed by a single-stranded tail containing the 5′-ACA-3′ motif situated 3 nt upstream from the mature 3′-end of the snoRNA (8,9). Mutations of conserved nucleotides of the H or ACA box impair accumulation and function of box H/ACA snoRNAs (8–10). Moreover, such mutations prevent the nucleolar accumulation of box H/ACA snoRNAs injected into Xenopus oocytes (11). The functional similarities between the H and ACA boxes of hTR and those of the box H/ACA snoRNAs were confirmed by mutational analyses. Mutations that modify the putative H or ACA box of hTR, so that they no longer conform to the consensus, drastically lower the hTR levels in HeLa cells (3). In contrast, mutations that result in an H box that does not deviate from the consensus have no effect on hTR accumulation (3). Similarities between box H/ACA snoRNAs and hTR extend also to their sub-nuclear localization. A small but significant proportion of hTR is found within nucleoli (3). Moreover, hTR injected into Xenopus oocytes accumulates within the fibrillar regions of nucleoli (11). Strikingly, mutating the ACA box of hTR prevents the accumulation of this RNA within nucleoli, as is observed for box H/ACA snoRNAs (11).
Whether the presence of an H/ACA-like domain in telomerase RNA is a feature specific to vertebrates is unclear. However, the phylogenetically supported secondary structure model proposed for the telomerase RNA of ciliates does not contain a domain obviously related to box H/ACA snoRNAs (12–14). No convincing secondary structure model for Saccharomyces cerevisiae telomerase RNA [TLC1 (15)] is yet available but the presence of a match to the consensus binding site of Sm proteins near the 3′-end of TLC1 RNA was noticed by Seto et al. (16). Remarkably, mutations within this putative Sm-binding site drastically lower the levels of the TLC1 RNA (16).
The discovery that hTR RNA contains an H/ACA-like domain crucial for its in vivo accumulation, 3′-end processing and intra-nuclear trafficking prompted the suggestion that hTR does associate with H/ACA snoRNP proteins. Four core H/ACA snoRNP proteins, all essential for viability, have been identified in yeast: Cbf5p (17–20), Gar1p (8,9,21), Nhp2p (20,22) and Nop10p (22). In the absence of Cbf5p, Nhp2p or Nop10p, H/ACA snoRNPs cannot accumulate in yeast cells (19,20,22). Both Gar1p (23) and Nhp2p (A.Henras, C.Dez, J.Noaillac-Depeyre, Y.Henry and M.Caizergues-Ferrer, manuscript submitted) have been proposed to directly contact box H/ACA snoRNAs. The human homologs of yeast Cbf5p, termed Dyskerin (24), and yeast Gar1p, termed hGar1p (25), have been identified. Immunoprecipitation experiments reveal that both Dyskerin and hGar1p are in fact associated with hTR (25,26).
To assess the functional importance of the association of hTR with H/ACA snoRNP proteins, we decided to express hTR in a genetically tractable system, S.cerevisiae. We demonstrate that the mature form of hTR is associated with all four H/ACA snoRNP proteins in yeast and that absence of Cbf5p, Nhp2p and Nop10p prevents the accumulation of the mature form of hTR. In contrast, we find that TLC1 is not associated with H/ACA snoRNP proteins and that depletion of these proteins has no effect on TLC1 levels.
MATERIALS AND METHODS
Strains, media and plasmids
Escherichia coli DH5α strain [F′, endA1, hsdr17 (rk– mk+), supE44, thi-1, recA1, gyrA (Nalr), relA1, Δ(lacIZYA-argF)U169, deoR, (φ80dlacΔ(lacZ)M15)] grown on LB (1% bacto-tryptone, 0.5% bacto-yeast extract, 1% NaCl) liquid or solid media was used for all cloning procedures.
Saccharomyces cerevisiae strains were grown in YNB medium [0.17% yeast nitrogen base, 0.5% (NH4)2SO4] supplemented with 2% glucose or 2% galactose and the required amino acids.
Plasmids allowing expression of hTR in yeast were produced as follows. The entire coding sequence of hTR was released from phTR1 (25) by SfcI and FspI digestions. The 5′-overhangs produced by the SfcI digestion were filled-in by treatment with the Klenow fragment of E.coli DNA polymerase I. The resulting blunt-ended fragment was inserted in plasmid pFL45esno (10), previously digested by KpnI and treated with T4 DNA polymerase to remove the 3′-overhangs left by the KpnI digestion. In the resulting plasmid, phTRTRP1, the sequence encoding hTR is inserted in-between the promoter and terminator regions of SNR5. phTRTRP1 contains the TRP1 selection marker. For transformation into certain strains expressing epitope-tagged proteins (see below), it was necessary to exchange the TRP1 marker in phTRTRP1 for either the LEU2 or URA3 markers. LEU2 and URA3 gene cassettes obtained from pFL46S (27) or pFL38 (27) by BglII digestion were used to replace the TRP1-containing BglII fragment of phTRTRP1, creating plasmids phTRLEU2 and phTRURA3, respectively. To avoid unnecessary details in the main text, the distinction between the different forms of phTR was not made in the Results section.
A strain expressing hTR and Cbf5pZZ was obtained by transforming strain YDL524-18 (19) with phTRTRP1. A strain expressing hTR and Gar1pZZ was obtained by transforming strain YO79 with plasmids pMCGZZ1 (22) and phTRLEU2. YO79 was derived from the GAL::gar1 strain YO24 (21) by disruption of the LEU2 locus by the LYS2 marker gene. A strain expressing hTR and Nhp2pZZ was obtained by transforming strain YO346 (A.Henras, C. Dez, J.Noaillac-Depeyre, Y.Henry and M.Caizergues-Ferrer, manuscript submitted) with plasmid phTRURA3. A strain expressing hTR and Nop10pZZ was obtained by transforming strain YO249 [a GAL::nop10ZZ strain described previously (22)] with plasmids phTRTRP1 and pFH35, a centromeric plasmid containing the LEU2 marker gene and the NOP10ZZ gene under the control of the NOP10 promoter. A strain expressing hTR and ZZNop1p was obtained by transforming strain JG540 (22) with plasmids phTRTRP1 and pUN100Nt (a gift from Dr T. Bergès, Université de Poitiers). Strains that contain the gene encoding hTR and that conditionally transcribe the CBF5, GAR1, NHP2 and NOP10 genes were obtained by transforming the GAL::cbf5 strain YDL521-1 (19), the GAL::gar1 strain YO24 (21), the GAL::nhp2 strain Y0253 (22) and the GAL::nop10 strain YO247 (22) with plasmid phTRTRP1.
RNA extractions, northern hybridizations and purification of polyA+ RNAs
RNA extractions were performed as described previously (28). Polyadenylated RNAs were purified using the ‘PolyA Tract mRNA Isolation System IV’ (Promega) as instructed by the supplier. Electrophoresis and transfer of RNAs to nylon membranes was performed as described previously (22). In most cases, 10 µg of total RNA was analyzed. In experiments presented in Figures 1 and 4, 7% of the polyadenylated and non-polyadenylated RNA fractions obtained from 300 µg of total RNA were used. Specific RNAs were detected by hybridization of membranes with kinased oligonucleotide probes. The following antisense oligonucleotides were used: anti-TLC1, 5′-GATCAATCCGAAATCCGACACTATCTCTTCACCATCGAG-3′; anti-hTR, 5′-CTGGGCAGGCGACCCGCCGCAGGTCCCCGGGAGGGGCGAACGGGCCAGCAGCT-3′; anti-yU3, 5′-ATGGGGCTCATCAACCAAGTTGG-3′; anti-snR37, 5′-GATAGTATTAACCACTACTG-3′. The sequence of the antisense oligonucleotide used to detect snR42 has been reported previously (22). Hybridization and washing conditions used with antisense oligonucleotide probes allowing detection of snoRNAs have been detailed previously (22). The anti-TLC1 oligonucleotide was hybridized to northern membranes overnight at 37°C in 6× SSC, 5× Denhardt’s, 0.5% SDS, 120 µg/ml heat-denatured salmon sperm DNA. Membranes were then washed twice in 2× SSC, 0.1% SDS at 37°C for 15 min and twice in 1× SSC, 0.1% SDS at 50°C for 15 min. The anti-hTR oligonucleotide was hybridized to northern membranes overnight at 37°C in 50% formamide, 5× SSC, 5× Denhardt’s, 1% SDS, 120 µg/ml heat-denatured salmon sperm DNA. Membranes were then washed once in 2× SSC, 0.1% SDS at 37°C for 15 min, once in 1× SSC, 0.1% SDS at 42°C for 15 min and finally twice in 0.1× SSC, 0.1% SDS at 48°C for 15 min each time.
Figure 1.
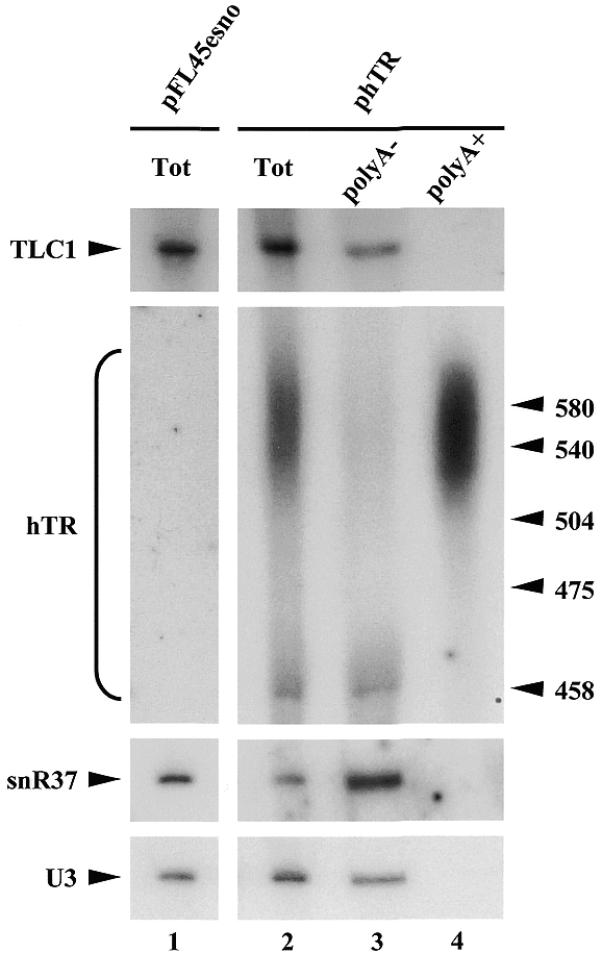
Both mature polyA– hTR RNA and polyA+ forms of this RNA can accumulate in S.cerevisiae. Total RNAs (Tot) were extracted from yeast strains transformed either with pFL45esno expression vector lacking insert (lane 1) or with the phTR plasmid, derived from pFL45esno, that carries the DNA fragment encoding hTR (lane 2). A sample of the total RNA preparation obtained from the strain containing phTR was incubated with biotinylated oligo(dT) and polyadenylated RNAs bound to oligo(dT) were precipitated using streptavidin-coated magnetic beads. RNAs were recovered from the supernatant (polyA–, lane 3) or the pellet (polyA+, lane 4). All RNAs were separated on a 6% polyacrylamide gel, transferred to a nylon membrane and hybridized with antisense oligonucleotide probes detecting the TLC1, hTR, snR37 or U3 RNA. Positions of molecular weight markers are indicated on the right.
Figure 4.
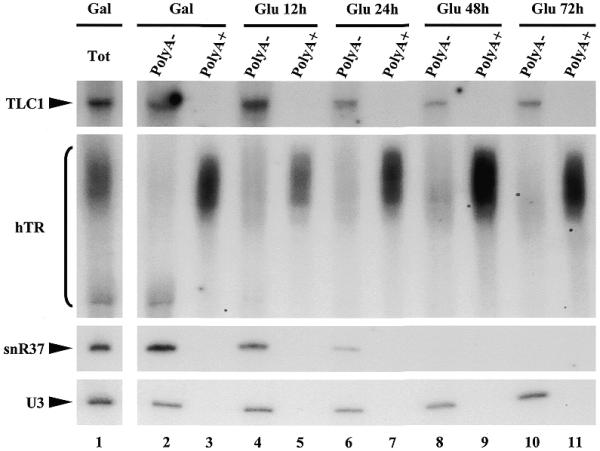
Accumulation in S.cerevisiae of polyadenylated forms of hTR and of polyA– mature TLC1 does not require the presence of Nhp2p. The GAL::nhp2 strain transformed with phTR was grown in galactose-containing medium (lanes 1–3) and was then transferred to glucose-containing medium for 12, 24, 48 and 72 h. Culture samples were collected from which total RNAs were extracted (Tot). Samples of the total RNA preparations were incubated with biotinylated oligo(dT) and polyadenylated RNAs were precipitated using streptavidin-coated magnetic beads. RNAs were recovered from the supernatants (polyA–) or the pellets (polyA+). All RNAs were separated on a 6% polyacrylamide gel, transferred to a nylon membrane and hybridized with antisense oligonucleotide probes detecting TLC1, hTR, snR37 or U3 RNA.
Immunoprecipitations
Immunoprecipitations of ZZ-tagged proteins from yeast extracts were performed as described previously (9).
RESULTS
Both mature non-polyadenylated hTR and a heterogeneous population of polyadenylated hTR species can accumulate in yeast cells
In order to assess the functional importance of the association of H/ACA snoRNP proteins with hTR, we wished to express this RNA in a genetically tractable system. Because it had been previously demonstrated that both human C/D-type and H/ACA-type snoRNAs could be expressed in yeast and were functional in this organism (10,29), we chose S.cerevisiae as our model system. The DNA fragment encoding the whole of hTR was cloned in the yeast expression vector pFL45esno (10), producing a plasmid we termed phTR. In such a construct, hTR expression is controlled by the promoter and terminator sequences of the yeast SNR5 gene that encodes the snR5 H/ACA snoRNA (30). pFL45esno and phTR were transformed in yeast and RNAs extracted from the resulting transformed strains were analyzed by northern hybridization. Blots were incubated with an antisense oligonucleotide probe designed to hybridize to nucleotides 181–233 of hTR. Using this probe, an RNA of the expected size for mature hTR (451 nt) as well as a heterogeneous population of extended species can be detected in the RNA sample extracted from the strain containing phTR (Fig. 1, lane 2). These species are absent in the RNA sample extracted from the strain transformed with pFL45esno (Fig. 1, lane 1). A fraction of certain snoRNAs (31) and yeast telomerase RNA (32) is polyadenylated in S.cerevisiae. Therefore, we tested whether the bulk of the extended species just described consists of polyadenylated hTR RNAs. A sample of total RNAs extracted from the yeast strain containing phTR was incubated with biotinylated oligo(dT) and polyadenylated RNAs bound to these oligonucleotides were precipitated using streptavidin-coated magnetic beads. RNAs recovered from the supernatant (lacking the polyadenylated species) or the pellet (containing the polyadenylated species) were analyzed by northern hybridization. The bulk of the extended species is recovered from the pellet (Fig. 1, lane 4), indicating that these species are polyadenylated. We conclude that in a yeast strain containing phTR, both mature non-polyadenylated hTR and polyadenylated forms of hTR accumulate.
Mature non-polyadenylated hTR is associated in yeast with all four core H/ACA snoRNP proteins, unlike the RNA component of yeast telomerase
To assess whether hTR species expressed in yeast are associated with all four H/ACA snoRNP proteins, immunoprecipitation experiments were performed. For that purpose, phTR was transformed in yeast strains expressing Cbf5p, Gar1p, Nhp2p or Nop10p proteins tagged at their C-terminus with two IgG-binding domains (ZZ domains) derived from Staphylococcus aureus protein A. We also transformed phTR in a strain expressing Nop1p tagged at its N-terminus with the two ZZ domains. Total cellular extracts were prepared from the resulting strains and immunoprecipitation experiments were carried out using IgG–Sepharose to which the ZZ moieties of the tagged proteins avidly bind. Northern analysis of RNAs extracted from the pellet fractions (Fig. 2) reveals that the mature non-polyadenylated form of hTR is co-immunoprecipitated along with the Cbf5pZZ, Gar1pZZ, Nhp2pZZ and Nop10pZZ proteins as efficiently as the endogenous yeast box H/ACA snoRNAs tested (Fig. 2, lanes 1–8). In contrast, only a minor fraction of the extended hTR species is precipitated. No co-precipitation of hTR RNAs with ZZ–Nop1p is observed (Fig. 2, lanes 9 and 10). We were also unable to detect an interaction between TLC1 and any of the core H/ACA snoRNP proteins (Fig. 2).
Figure 2.
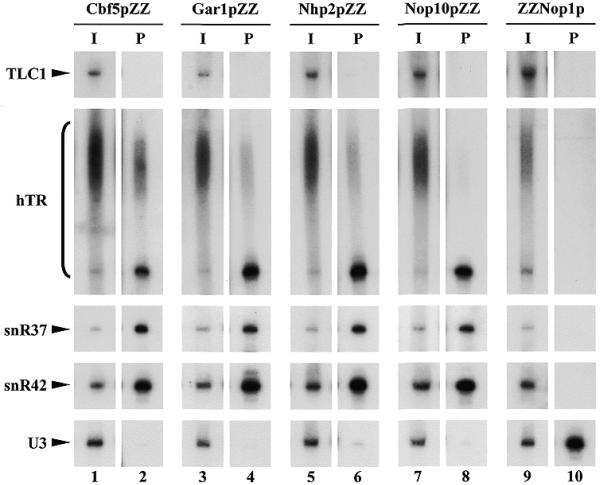
Mature polyA– hTR expressed in yeast interacts with all four core proteins of H/ACA snoRNPs. Immunoprecipitation experiments were carried out using IgG–Sepharose and extracts from strains expressing hTR and either Cbf5pZZ (lanes 1 and 2), Gar1pZZ (lanes 3 and 4), Nhp2pZZ (lanes 5 and 6), Nop10pZZ (lanes 7 and 8) or ZZNop1p (lanes 9 and 10). RNAs extracted from one-tenth of the input extracts (I) or from the pellets following immunoprecipitation (P) were separated on a 6% polyacrylamide gel and transferred to a nylon membrane. RNAs indicated on the left were detected by hybridization with antisense oligonucleotide probes.
Cbf5p, Nhp2p and Nop10p are required for the accumulation of mature non-polyadenylated hTR in yeast
We have previously shown that H/ACA snoRNPs cannot accumulate if cells are deprived of Cbf5p (19), Nhp2p (22) or Nop10p (22). In contrast, H/ACA particles can assemble in the absence of Gar1p, although these remain non-functional (33). To assess the consequences of the depletion of any of the four core H/ACA snoRNP proteins on hTR accumulation, plasmid phTR was transformed into strains GAL::cbf5 (19), GAL::gar1 (21), GAL::nhp2 (22) and GAL::nop10 (22). In these strains, the endogenous promoters of the CBF5, GAR1, NHP2 or NOP10 genes have been replaced by a GAL promoter, to ensure conditional transcription of these genes: on galactose-containing medium, their transcription is induced whereas it is repressed on glucose-containing medium. Strains GAL::cbf5, GAL::gar1, GAL::nhp2 and GAL::nop10 transformed by phTR were propagated on galactose-containing minimal media and were then shifted to glucose-containing minimal media. Samples from galactose-containing cultures and from cultures grown on glucose-containing media for 12, 24, 48 and 72 h were collected. Total proteins and RNAs were extracted from these samples for western (data not shown) and northern analyses (Fig. 3). Western analyses reveal that the steady-state levels of all four H/ACA snoRNP proteins are strongly reduced after 12 h of growth in glucose-containing media and that these proteins are almost undetectable after 24 h of growth in such media (data not shown). Depletion of Cbf5p, Nhp2p and Nop10p is correlated with a drastic decrease in the steady-state levels of the mature non-polyadenylated form of hTR (Fig. 3). In fact, levels of hTR seem to decrease even faster during the depletion time-courses than those of endogenous H/ACA snoRNAs snR37 or snR42. In contrast, the absence of Gar1p does not reduce the levels of the mature non-polyadenylated form of hTR. Interestingly, the steady-state levels of the heterogeneous extended hTR species, that seem to only weakly associate with H/ACA snoRNP proteins (Fig. 2), do not appear to be significantly affected by the depletion of any one of these proteins. Moreover, the steady-state levels of the RNA component of yeast telomerase remain unchanged.
Figure 3.
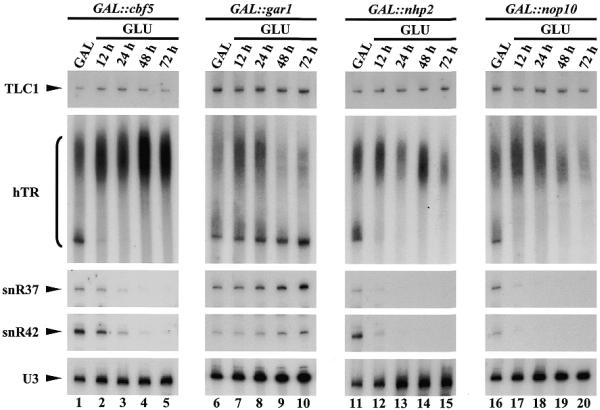
Accumulation in S.cerevisiae of mature polyA– hTR requires the presence of Cbf5p, Nhp2p and Nop10p. GAL::cbf5 (lanes 1–5), GAL::gar1 (lanes 6–10), GAL::nhp2 (lanes 11–15) and GAL::nop10 (lanes 16–20) strains transformed with phTR were grown in galactose-containing medium (lanes 1, 6, 11 and 16) and were then transferred to glucose-containing medium for 12, 24, 48 or 72 h. At each time-point, culture samples were collected from which total RNAs were extracted. These were separated on 6% polyacrylamide gels and transferred to nylon membranes. RNAs indicated on the left were detected by hybridization with specific oligonucleotide probes.
It has been reported that in wild-type yeast cells, a minor fraction (∼5%) of total TLC1 is polyadenylated and that this form is the precursor to mature non-polyadenylated TLC1 (32). The possibility that depletion of H/ACA snoRNP proteins results in over-accumulation of the polyadenylated form of TLC1 that would mask a decrease in the levels of mature non-polyadenylated TLC1 could not be excluded (the polyA+ and polyA– TLC1 species differ only by ∼80 nt and are difficult to distinguish). To rule out this possibility, polyA+ RNAs were purified from RNA samples extracted from the GAL::nhp2 strain containing phTR grown in glucose-containing medium for different times. As shown in Figure 4, the TLC1 species that are detected in RNA samples extracted from Nhp2p depleted cells clearly correspond to deadenylated TLC1. In contrast, the bulk of the extended heterogeneous hTR species that accumulate in Nhp2p depleted cells is polyadenylated.
DISCUSSION
The discovery that hTR RNA contains a domain related to box H/ACA snoRNAs prompted the suggestion that human telomerase contains at least a subset of the H/ACA snoRNP proteins. This was confirmed by the recent findings that hTR RNA is associated with two of these proteins, Dyskerin and hGar1p (25,26). What could be the role(s) of these proteins regarding telomerase assembly and activity? The two most likely hypotheses are that H/ACA snoRNP proteins are involved (i) in the intranuclear trafficking of the telomerase complex and (ii) in the 3′-end processing of the telomerase RNA component and hence are required for its normal accumulation. These two processes are in fact likely to be interconnected. To investigate the second hypothesis, we decided to express hTR in S.cerevisiae, in which the production of the four known H/ACA snoRNP proteins can be easily turned off. We find that the mature form of hTR can accumulate in yeast, an observation also reported by Autexier and coworkers (34,35) while our work was in progress. We provide evidence that mature hTR is associated in yeast with the four known H/ACA snoRNP proteins, including Nhp2p and Nop10p. It is therefore likely that the human counterparts of the yeast Nhp2p and Nop10p proteins are also associated with hTR in human cells.
In addition to the mature form of hTR, a heterogeneous population of extended forms of hTR accumulate in yeast. Most of these species are polyadenylated. Polyadenylation of RNA species other than mRNAs has already been documented in yeast. In particular, 5–10% of TLC1, the RNA component of S.cerevisiae telomerase, is polyadenylated at steady-state (32). It is envisaged, but not proven, that the polyadenylated form of TLC1 is the precursor to the mature form of TLC1 that lacks a polyA tail (32). Interestingly, a small proportion of the yeast snR33 H/ACA snoRNA is also polyadenylated in wild-type cells (31). This polyadenylated form of snR33 may also be the precursor to the mature polyA– snR33 snoRNA. We do not know at this stage whether the polyadenylated forms of hTR that accumulate in yeast correspond to dead-end products or whether they constitute precursors to mature non-polyadenylated hTR. Polyadenylated forms of hTR have not yet been reported in human cells (36), but these forms may represent a very minor fraction of the steady-state hTR population and could therefore have gone undetected (32).
Whatever the nature of the precursor(s) to mature non-polyadenylated hTR in yeast cells, it is clear from our data that accumulation of this RNA species in S.cerevisiae depends on its association with the core H/ACA snoRNP proteins Cbf5p, Nhp2p and Nop10p. This is most likely the case in human cells as well. Indeed, the importance of Dyskerin (the human homolog of Cbf5p) for the accumulation of hTR in human cells is underscored by the following observation. Levels of hTR are found to be lower in primary fibroblasts from patients suffering from the X-linked form of the disease dyskeratosis congenita, that express a mutated form of Dyskerin, than in corresponding cells from healthy individuals (26).
Depletion of Gar1p, the fourth known core component of H/ACA snoRNPs, has no effect on the accumulation of mature hTR in yeast. Remarkably, accumulation of box H/ACA snoRNAs is dependent on Cbf5p (19), Nhp2p (22) and Nop10p (22) but is also unaffected by depletion of Gar1p (21; this work). This strongly suggests that the four core H/ACA snoRNP proteins assemble with the H/ACA domain of hTR and with box H/ACA snoRNAs in a similar fashion.
No secondary structure model supported by phylogenetic comparison is yet available for the very large TLC1 RNA (1.3 kb). Given the scarcity of conserved nucleotides in box H/ACA snoRNAs, it is difficult to rule out the possibility, by mere inspection of the primary structure of TLC1, that this RNA contains an H/ACA-like domain. Our data strongly suggest that mature TLC1 is not associated with any of the four known H/ACA snoRNP proteins, and show that depletion of any of these proteins has no effect on the accumulation of mature TLC1. From these results, we conclude that no functional H/ACA-like motif is present in TLC1. In contrast, convincing evidence has been provided that TLC1 contains a binding site for the Sm proteins, which are common to several UsnRNPs (16). Both the integrity of the Sm site on the TLC1 RNA and the presence of at least two of the Sm proteins are needed for accumulation of the mature form of TLC1 (16). Thus it seems that Sm protein binding to TLC1 RNA is required for its accumulation, just as assembly of H/ACA snoRNP proteins on the H/ACA domain of hTR is required for the accumulation of this RNA.
The following model can be put forward to explain how H/ACA snoRNP proteins participate in hTR 3′-end formation and promote hTR accumulation. The H/ACA snoRNP protein(s) bound to the ACA box (situated just 3 nt upstream from the 3′-end of mature hTR) could stop the advance of 3′→5′ exonucleases, for example components of the human exosome complex (37), during maturation of a 3′-extended hTR precursor. In this model, box H/ACA proteins provide a barrier to further degradation, thus preventing the turnover of hTR and defining the 3′-end of the mature form of this RNA. In addition, the possibility that H/ACA snoRNP proteins also actively promote the digestion of the 3′-extension of hTR precursors by exonucleases cannot be excluded (38). It is less easy to propose a model explaining how Sm proteins promote accumulation of TLC1. It will be interesting to assess whether the Sm protein binding site of TLC1 can be substituted for the H/ACA domain in hTR and vice versa.
Acknowledgments
ACKNOWLEDGEMENTS
We wish to thank Dr T. Kiss (LBME, Toulouse) for the gift of plasmid pFL45esno, Dr T. Bergès (Université de Poitiers) for the gift of plasmid pUN100Nt and Dr F. Dragon (Yale University), Prof. W. Fillipowicz and V. Pogacic (FMI, Basel) for the gift of plasmid phTR1 and communication of results prior to publication. The involvement of S. Arico in initial stages of this work is gratefully acknowledged. We are thankful to members of the Ferrer lab for help and numerous discussions. We thank Y. de Préval for synthesis of oligonucleotides and D. Villa for artwork. C.D. and A.H. are supported by grants from the Ministère de l’Education Nationale, de l’Enseignement Supérieur et de la Recherche and la Ligue Nationale contre le Cancer, respectively. D.L.J.L. is a Chercheur qualifié du F.N.R.S. This work was financed by la Ligue Nationale contre le Cancer, the CNRS and Université Paul Sabatier, Toulouse.
References
- 1.Nugent C.I. and Lundblad,V. (1998) The telomerase reverse transcriptase: components and regulation. Genes Dev., 12, 1073–1085. [DOI] [PubMed] [Google Scholar]
- 2.Feng J., Funk,W.D., Wang,S.S., Weinrich,S.L., Avilion,A.A., Chiu,C.P., Adams,R.R., Chang,E., Allsopp,R.C., Yu,J. et al. (1995) The RNA component of human telomerase. Science, 269, 1236–1241. [DOI] [PubMed] [Google Scholar]
- 3.Mitchell J.R., Cheng,J. and Collins,K. (1999) A box H/ACA small nucleolar RNA-like domain at the human telomerase RNA 3′ end. Mol. Cell. Biol., 19, 567–576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Chen J.L., Blasco,M.A. and Greider,C.W. (2000) Secondary structure of vertebrate telomerase RNA. Cell, 100, 503–514. [DOI] [PubMed] [Google Scholar]
- 5.Ganot P., Bortolin,M.L. and Kiss,T. (1997) Site-specific pseudouridine formation in preribosomal RNA is guided by small nucleolar RNAs. Cell, 89, 799–809. [DOI] [PubMed] [Google Scholar]
- 6.Ni J., Tien,A.L. and Fournier,M.J. (1997) Small nucleolar RNAs direct site-specific synthesis of pseudouridine in ribosomal RNA. Cell, 89, 565–573. [DOI] [PubMed] [Google Scholar]
- 7.Tollervey D. and Kiss,T. (1997) Function and synthesis of small nucleolar RNAs. Curr. Opin. Cell Biol., 9, 337–342. [DOI] [PubMed] [Google Scholar]
- 8.Balakin A.G., Smith,L. and Fournier,M.J. (1996) The RNA world of the nucleolus: two major families of small RNAs defined by different box elements with related functions. Cell, 86, 823–834. [DOI] [PubMed] [Google Scholar]
- 9.Ganot P., Caizergues-Ferrer,M. and Kiss,T. (1997) The family of box ACA small nucleolar RNAs is defined by an evolutionarily conserved secondary structure and ubiquitous sequence elements essential for RNA accumulation. Genes Dev., 11, 941–956. [DOI] [PubMed] [Google Scholar]
- 10.Bortolin M.L., Ganot,P. and Kiss,T. (1999) Elements essential for accumulation and function of small nucleolar RNAs directing site-specific pseudouridylation of ribosomal RNAs. EMBO J., 18, 457–469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Narayanan A., Lukowiak,A., Jady,B.E., Dragon,F., Kiss,T., Terns,R.M. and Terns,M.P. (1999) Nucleolar localization signals of box H/ACA small nucleolar RNAs. EMBO J., 18, 5120–5130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Romero D.P. and Blackburn,E.H. (1991) A conserved secondary structure for telomerase RNA. Cell, 67, 343–353. [DOI] [PubMed] [Google Scholar]
- 13.ten Dam E., van Belkum,A. and Pleij,K. (1991) A conserved pseudoknot in telomerase RNA. Nucleic Acids Res., 19, 6951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lingner J., Hendrick,L.L. and Cech,T.R. (1994) Telomerase RNAs of different ciliates have a common secondary structure and a permuted template. Genes Dev., 8, 1984–1998. [DOI] [PubMed] [Google Scholar]
- 15.Singer M.S. and Gottschling,D.E. (1994) TLC1: template RNA component of Saccharomyces cerevisiae telomerase. Science, 266, 404–409. [DOI] [PubMed] [Google Scholar]
- 16.Seto A.G., Zaug,A.J., Sobel,S.G., Wolin,S.L. and Cech,T.R. (1999) Saccharomyces cerevisiae telomerase is an Sm small nuclear ribonucleoprotein particle. Nature, 401, 177–180. [DOI] [PubMed] [Google Scholar]
- 17.Jiang W., Middleton,K., Yoon,H.J., Fouquet,C. and Carbon,J. (1993) An essential yeast protein, CBF5p, binds in vitro to centromeres and microtubules. Mol. Cell. Biol., 13, 4884–4893. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cadwell C., Yoon,H.J., Zebarjadian,Y. and Carbon,J. (1997) The yeast nucleolar protein Cbf5p is involved in rRNA biosynthesis and interacts genetically with the RNA polymerase I transcription factor RRN3. Mol. Cell. Biol., 17, 6175–6183. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lafontaine D.L.J., Bousquet-Antonelli,C., Henry,Y., Caizergues-Ferrer,M. and Tollervey,D. (1998) The box H + ACA snoRNAs carry Cbf5p, the putative rRNA pseudouridine synthase. Genes Dev., 12, 527–537. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Watkins N.J., Gottschalk,A., Neubauer,G., Kastner,B., Fabrizio,P., Mann,M. and Luhrmann,R. (1998) Cbf5p, a potential pseudouridine synthase, and Nhp2p, a putative RNA-binding protein, are present together with Gar1p in all H BOX/ACA-motif snoRNPs and constitute a common bipartite structure. RNA, 4, 1549–1568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Girard J.-P., Lehtonen,H., Caizergues-Ferrer,M., Amalric,F., Tollervey,D. and Lapeyre,B. (1992) GAR1 is an essential small nucleolar RNP protein required for pre-rRNA processing in yeast. EMBO J., 11, 673–682. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Henras A., Henry,Y., Bousquet-Antonelli,C., Noaillac-Depeyre,J., Gélugne,J.-P. and Caizergues-Ferrer,M. (1998) Nhp2p and Nop10p are essential for the function of H/ACA snoRNPs. EMBO J., 17, 7078–7090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Bagni C. and Lapeyre,B. (1998) Gar1p binds to the small nucleolar RNAs snR10 and snR30 in vitro through a nontypical RNA binding element. J. Biol. Chem., 273, 10868–10873. [DOI] [PubMed] [Google Scholar]
- 24.Heiss N.S., Knight,S.W., Vulliamy,T.J., Klauck,S.M., Wiemann,S., Mason,P.J., Poustka,A. and Dokal,I. (1998) X-linked dyskeratosis congenita is caused by mutations in a highly conserved gene with putative nucleolar functions. Nature Genet., 19, 32–38. [DOI] [PubMed] [Google Scholar]
- 25.Dragon F., Pogacic,V. and Filipowicz,W. (2000) In vitro assembly of human H/ACA small nucleolar RNPs reveals unique features of U17 and telomerase RNAs. Mol. Cell. Biol., 20, 3037–3048. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Mitchell J.R., Wood,E. and Collins,K. (1999) A telomerase component is defective in the human disease dyskeratosis congenita. Nature, 402, 551–555. [DOI] [PubMed] [Google Scholar]
- 27.Bonneaud N., Ozier-Kalogeropoulos,O., Li,G.Y., Labouesse,M., Minvielle-Sebastia,L. and Lacroute,F. (1991) A family of low and high copy replicative, integrative and single-stranded S. cerevisiae/E. coli shuttle vectors. Yeast, 7, 609–615. [DOI] [PubMed] [Google Scholar]
- 28.Tollervey D. and Mattaj,I.W. (1987) Fungal small nuclear ribonucleoproteins share properties with plant and vertebrate U-snRNPs. EMBO J., 6, 469–476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kiss-Laszlo Z., Henry,Y. and Kiss,T. (1998) Sequence and structural elements of methylation guide snoRNAs essential for site-specific ribose methylation of pre-rRNA. EMBO J., 17, 797–807. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Parker R., Simmons,T., Shuster,E.O., Siliciano,P.G. and Guthrie,C. (1988) Genetic analysis of small nuclear RNAs in Saccharomyces cerevisiae: viable sextuple mutant. Mol. Cell. Biol., 8, 3150–3159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.van Hoof A., Lennertz,P. and Parker,R. (2000) Yeast exosome mutants accumulate 3′-extended polyadenylated forms of U4 small nuclear RNA and small nucleolar RNAs. Mol. Cell. Biol., 20, 441–452. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Chapon C., Cech,T.R. and Zaug,A.J. (1997) Polyadenylation of telomerase RNA in budding yeast. RNA, 3, 1337–1351. [PMC free article] [PubMed] [Google Scholar]
- 33.Bousquet-Antonelli C., Henry,Y., Gélugne J.-P., Caizergues-Ferrer,M. and Kiss,T. (1997) A small nucleolar RNP protein is required for pseudouridylation of eukaryotic ribosomal RNAs. EMBO J., 16, 4770–4776. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Bachand F. and Autexier,C. (1999) Functional reconstitution of human telomerase expressed in Saccharomyces cerevisiae. J. Biol. Chem., 274, 38027–38031. [DOI] [PubMed] [Google Scholar]
- 35.Bachand F., Kukolj,G. and Autexier,C. (2000) Expression of hTERT and hTR in cis reconstitutes and active human telomerase ribonucleoprotein. RNA, 6, 778–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Zaug A.J., Linger,J. and Cech,T.R. (1996) Method for determining RNA 3′-ends and application to human telomerase RNA. Nucleic Acids Res., 24, 532–533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Allmang C., Petfalski,E., Podtelejnikov,A., Mann,M., Tollervey,D. and Mitchell,P. (1999) The yeast exosome and human PM-Scl are related complexes of 3′→5′ exonucleases. Genes Dev., 13, 2148–2158. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Kufel J., Allmang,C., Chanfreau,G., Petfalski,E., Lafontaine,D.L. and Tollervey,D. (2000) Precursors to the U3 small nucleolar RNA lack small nucleolar RNP proteins but are stabilized by La binding. Mol. Cell. Biol., 20, 5415–5424. [DOI] [PMC free article] [PubMed] [Google Scholar]