

Abstract
OBJECTIVE
The metabolic syndrome (MetS) is defined as concomitant disorders of lipid and glucose metabolism, central obesity, and high blood pressure, with an increased risk of type 2 diabetes and cardiovascular disease. This study tests whether common genetic variants with pleiotropic effects account for some of the correlated architecture among five metabolic phenotypes that define MetS.
RESEARCH DESIGN AND METHODS
Seven studies of the STAMPEED consortium, comprising 22,161 participants of European ancestry, underwent genome-wide association analyses of metabolic traits using a panel of ∼2.5 million imputed single nucleotide polymorphisms (SNPs). Phenotypes were defined by the National Cholesterol Education Program (NCEP) criteria for MetS in pairwise combinations. Individuals exceeding the NCEP thresholds for both traits of a pair were considered affected.
RESULTS
Twenty-nine common variants were associated with MetS or a pair of traits. Variants in the genes LPL, CETP, APOA5 (and its cluster), GCKR (and its cluster), LIPC, TRIB1, LOC100128354/MTNR1B, ABCB11, and LOC100129150 were further tested for their association with individual qualitative and quantitative traits. None of the 16 top SNPs (one per gene) associated simultaneously with more than two individual traits. Of them 11 variants showed nominal associations with MetS per se. The effects of 16 top SNPs on the quantitative traits were relatively small, together explaining from ∼9% of the variance in triglycerides, 5.8% of high-density lipoprotein cholesterol, 3.6% of fasting glucose, and 1.4% of systolic blood pressure.
CONCLUSIONS
Qualitative and quantitative pleiotropic tests on pairs of traits indicate that a small portion of the covariation in these traits can be explained by the reported common genetic variants.
Metabolic syndrome (MetS) is defined as a combination of any three metabolic abnormalities, including central obesity, dyslipidemia, insulin resistance and/or glucose intolerance, and elevated blood pressure. These abnormalities tend to cluster in an individual and within families. Using the National Cholesterol Education Program (NCEP) MetS definition, the prevalence among adults in the U.S. was reported to be 23.8% in Caucasians, 21.6% in African Americans, and 31.9% in Mexican Americans (1,2) and 30.2% in adult Finns 40–65 years of age (3). MetS is associated with a higher risk of type 2 diabetes and cardiovascular disease and death (4–6). Because MetS risk factors have been reported to be heritable, we hypothesized that genes with pleiotropic effects may be responsible for some of the clustering of metabolic abnormalities observed in MetS.
MetS has been previously investigated using multivariate analysis (7–9) and pairwise combinations of its components (10,11). In an investigation of lipid traits, Kullo et al. (12) concluded that pleiotropy (when a gene influences multiple traits) contributes to the covariation among three lipid traits, high-density lipoprotein cholesterol (HDLC), triglycerides (TG), and low-density lipoprotein particle size, supporting the hypothesis of genetic pleiotropy as a source of correlation among metabolic traits (13). Although several recent genome-wide association studies (GWAS) have been carried out on MetS components individually (obesity/dyslipidemia/blood pressure [BP] or type 2 diabetes) in several populations (14–21), it is unknown whether the identified variants influence combinations of MetS components.
The current study leverages GWAS results from seven studies participating in the single nucleotide polymorphisms (SNP) Typing for Association with Multiple Phenotypes from Existing Epidemiologic Data (STAMPEED) Consortium. Using the NCEP thresholds for a series of five traits including waist circumference (WC), fasting glucose (GLUC), HDLC, TG, and BP, we created an affected status for all pairwise combinations of traits in which individuals exceeding the threshold for both traits were considered affected and all others were considered unaffected. By performing association tests with these 10 binary traits, as well as MetS per se, we sought to identify common genetic variants that affect the correlated architecture of these metabolic traits.
RESEARCH DESIGN AND METHODS
The STAMPEED Consortium, established in 2007, was sponsored by the National Heart, Lung, and Blood Institute (NHLBI) and includes 13 independent studies. In this report, we analyze a total of 22,161 participants of European ancestry, originating from seven studies, in alphabetic order: the Atherosclerotic Disease, Vascular function, and Genetic Epidemiology (ADVANCE) study (cases and controls separately), the Atherosclerosis Risk in Communities Study (ARIC) study, the Cardiovascular Health Study (CHS), the NHLBI Family Heart Study (FHS), the Genetic Epidemiology Network of Arteriopathy (GENOA), the Genetic Study of Aspirin Responsiveness (GeneSTAR), and the Northern Finland Birth Cohort 1966 (NFBC). Additional information regarding the characteristics of the original studies (regardless of any subsetting for GWAS genotyping) is available in the Supplementary Data.
The NCEP defines thresholds for five metabolic traits: WC ≥102 cm for men or WC ≥88 cm for women, HDLC <40 mg/dL for men or HDLC <50 mg/dL for women, TG ≥150 mg/dL, GLUC ≥100 mg/dL, and BP threshold as one of the three, systolic BP (SBP) or diastolic BP (DBP) ≥130/85 mmHg or antihypertensive medication use. We defined 10 bivariate traits, HDLC-WC, HDLC-TG, HDLC-GLUC, BP-HDLC, WC-TG, TG-GLUC, TG-BP, WC-GLUC, BP-GLUC, and WC-BP, in which an individual was considered affected only if s/he exceeded these thresholds for both traits in a pair, otherwise unaffected. For comparison, we also analyzed a MetS per se trait where affected subjects exceed the specified threshold for three or more traits, of the five traits according to NCEP criteria.
In each study, ∼2.5 million SNPs were imputed based on the HapMap (of European origin) CEU panel. Each study provided GWA results for 11 traits to the Data Coordinating Center (Division of Statistical Genomics, Washington University in Saint Louis, MO) for the meta-analysis. SNPs with R2-hat <0.3 (for studies that imputed using MACH), proper info <0.4 (IMPUTE), or variance ratio <0.3 (BIMBAM), as well as those with study-specific minor allele frequency <5% were filtered out from meta-analysis. As a result, these percentages of imputed markers were used in the meta-analyses: ADVANCE case, 81.4%; ADVANCE control, 81.2%; ARIC, 85.2%; CHS, 69.6%; FHS, 85.5%; GeneSTAR, 79.4%; GENOA, 84.1%; and NFBC, 86.1%; representing ∼2 million SNPs for the combined analyses. Based on the Q-Q plots for GWAS results, all studies demonstrated a reasonable compliance to the null expectation. A homogeneity test for each SNP across studies is reported in Supplementary Tables 1–6; the tests were not significant, indicating the data could be combined for the meta-analysis.
Statistical methods.
The 11 traits were tested for association with each SNP assuming additive SNP effects: 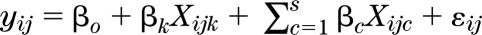 , where the trait yij is a function of βo, the intercept βk represents the k-th β coefficient from the regression analysis on imputed dosage, evaluated for each SNP (k = 1, 2, …, m), βc is the coefficient for covariate effects (c = 1, 2, …, s); and εij represents the residual for subject j (j = 1, 2, …, nID) in pedigree i (i =1, …, nPID). Individual studies used various software packages to carry out the GWAS (specified in Supplementary Data). Model covariates included sex, age, age2, and up to 10 genotypic principal components describing population substructure in the model.
, where the trait yij is a function of βo, the intercept βk represents the k-th β coefficient from the regression analysis on imputed dosage, evaluated for each SNP (k = 1, 2, …, m), βc is the coefficient for covariate effects (c = 1, 2, …, s); and εij represents the residual for subject j (j = 1, 2, …, nID) in pedigree i (i =1, …, nPID). Individual studies used various software packages to carry out the GWAS (specified in Supplementary Data). Model covariates included sex, age, age2, and up to 10 genotypic principal components describing population substructure in the model.
In preparation for meta-analyses, marker panels were aligned with dbSNP, build 36.3 for allele orientation. A meta-analysis of the GWAS results was conducted using a random coefficients model as proposed by DerSimonian and Laird (Supplementary Ref. 18) implemented through the MIXED procedure of SAS. This analysis returns combined estimates of the SNP β coefficients and standard errors, as well as the overall significance level accounting for the variance among estimates across studies. The homogeneity test is conducted to assess whether the β coefficients are combinable across studies (i = 1 to k). This test statistic is 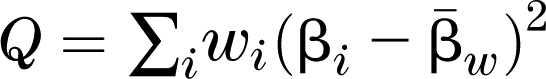 , where βi are β coefficients across studies, and the
, where βi are β coefficients across studies, and the 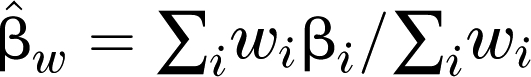 , where wi is inverse of the i-th sampling variance. The Q statistic has an approximate χ2 with k-1 degrees of freedom.
, where wi is inverse of the i-th sampling variance. The Q statistic has an approximate χ2 with k-1 degrees of freedom.
We adopted a genome-wide significance criterion of P ≤ 9.7*10−8, corresponding to a negative log10 P value of 7.01. This threshold corresponds to the Bonferroni corrected level, by calculating the effective number of independent comparisons after accounting for linkage disequilibrium (LD) among SNPs (Supplementary Ref. 19). Post hoc analyses included investigation of the LD structure in gene regions of interest using HaploView v. 4.2, with HapMap data (v3 release 2/ v2 release 22) on subjects of European descent (CEU).
The top SNPs (that is, those meeting genome-wide significance, choosing one SNP per gene with the smallest P value) identified by the meta-analyses were tested in each study population for their association with each of the five dichotomized traits: WC, HDLC, TG, GLUC, and BP. In addition, we carried out a specific test of pleiotropy using the full range of variation in the relevant quantitative traits. For subjects using lipid-lowering and/or antihypertensive medications, we imputed untreated traits values based on the estimated average effect of medications from clinical trials, as follows: TG/(1 − 15.2/100); HDLC/(1 + 6.1/100); and SBP + 14.8 mmHg (Supplementary Ref. 17). An appropriate transformation of the quantitative traits to obtain a good approximation to normality was applied, as needed (e.g., all studies used ln TG). The pleiotropy test on quantitative traits is a test of the simultaneous effect of the SNP on both quantitative traits, while allowing for a residual correlation: 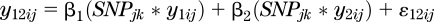 , where y12ij represents a combined vector of traits 1 and 2 for each observation of subject j in family i (if families are present in the data); y1ij and y2ij each represent indicators of 1 s and 0 s if y12ij belong to trait 1 or 2, respectively. The β1 and β2 estimates represent the additive contributions of k-th SNP on traits 1 and 2, respectively. We tested the null hypothesis H0: β1 = 0 and β2 = 0 versus its alternative H1: β1 ≠ 0 and β2 ≠ 0 with degrees of freedom equal to the number of subjects minus 2. In the family-based studies, subjects were nested within their corresponding pedigree and were considered statistically as repeated units. The tests were implemented with the MIXED procedure of SAS, v. 9.2. All the P values from each study were meta-analyzed.
, where y12ij represents a combined vector of traits 1 and 2 for each observation of subject j in family i (if families are present in the data); y1ij and y2ij each represent indicators of 1 s and 0 s if y12ij belong to trait 1 or 2, respectively. The β1 and β2 estimates represent the additive contributions of k-th SNP on traits 1 and 2, respectively. We tested the null hypothesis H0: β1 = 0 and β2 = 0 versus its alternative H1: β1 ≠ 0 and β2 ≠ 0 with degrees of freedom equal to the number of subjects minus 2. In the family-based studies, subjects were nested within their corresponding pedigree and were considered statistically as repeated units. The tests were implemented with the MIXED procedure of SAS, v. 9.2. All the P values from each study were meta-analyzed.
For all studies, informed consent was obtained from all subjects and approval was granted by participating institutional review boards.
RESULTS
The sample characteristics for the subjects with both valid phenotype and genotype information in each respective study are shown in Table 1. The study populations showed large variation in the prevalence of MetS from 9% in the NFBC sample to 55% in GENOA, in part, reflecting different ascertainment strategies and age ranges represented in each study; the average age ranged from 31 years in the NFBC 1966 study to 73 years in the CHS study (Table 1). Of the five MetS components, TG and HDLC had the strongest average correlation of any combination of traits (Supplementary Fig. 1).
TABLE 1.
Characteristics of the participants included in genome-wide association analyses (with both nonmissing genotype and phenotype data)
| Sample characteristic | ADVANCE control (up to 311) | ADVANCE cases (up to 275) | ARIC (up to 8,127) | CHS (up to 3,262) | FHS (up to 2,432) | GeneSTAR (up to 1,789) | GENOA (up to 1,208) | NFBC (up to 4,757) |
|---|---|---|---|---|---|---|---|---|
| % Females | 59 | 59 | 53 | 61 | 55 | 52 | 55 | 52 |
| % Diabetic* | 2.5 | 27 | 8 | 12 | 5 | 9 | 10 | 1 |
| % Smoking† | 32.8 | 63.3 | 25 | 11 | 29 | 23 | 14 | 30 |
| Age (years) | ||||||||
| Men | 40 ± 3 | 42 ± 4 | 55 ± 6 | 73 ± 6 | 50 ± 13 | 47 ± 13 | 56 ± 11 | 31 ± 1 |
| Women | 48 ± 5 | 52 ± 4 | 54 ± 6 | 72 ± 5 | 51 ± 13 | 47 ± 13 | 55 ± 11 | 31 ± 1 |
| BMI (kg/m2) | ||||||||
| Men | 27 ± 5 | 31 ± 6 | 27 ± 4 | 26 ± 4 | 28 ± 4 | 29 ± 5 | 30 ± 5 | 25 ± 4 |
| Women | 26 ± 6 | 31 ± 8 | 27 ± 6 | 26 ± 5 | 27 ± 6 | 29 ± 7 | 31 ± 7 | 24 ± 5 |
| Waist circumference (cm) | ||||||||
| Men | 93 ± 12 | 102 ± 14 | 100 ± 10 | 98 ± 10 | 100 ± 12 | 101 ± 14 | 104 ± 12 | 89 ± 10 |
| Women | 79 ± 13 | 94 ± 19 | 93 ± 15 | 90 ± 14 | 94 ± 16 | 94 ± 17 | 96 ± 17 | 79 ± 12 |
| HDL cholesterol (mg/dL) | ||||||||
| Men | 44 ± 11 | 40 ± 10 | 43 ± 12 | 48 ± 13 | 43 ± 11 | 45 ± 13 | 45 ± 13 | 55 ± 13 |
| Women | 62 ± 14 | 51 ± 16 | 57 ± 17 | 60 ± 16 | 56 ± 15 | 57 ± 15 | 57 ± 17 | 66 ± 15 |
| Triglycerides (mg/dL) | ||||||||
| Men | 151 ± 224 | 184 ± 165 | 147 ± 100 | 138 ± 78 | 163 ± 113 | 160 ± 97 | 194 ± 107 | 119 ± 75 |
| Women | 97 ± 69 | 162 ± 145 | 128 ± 81 | 141 ± 74 | 138 ± 87 | 133 ± 79 | 188 ± 102 | 94 ± 52 |
| Fasting glucose (mg/dL) | ||||||||
| Men | 86 ± 11 | 104 ± 43 | 107 ± 27 | 111 ± 31 | 102 ± 29 | 99 ± 27 | 103 ± 32 | 93 ± 11 |
| Women | 88 ± 11 | 95 ± 18 | 102 ± 28 | 105 ± 27 | 95 ± 22 | 92 ± 21 | 96 ± 27 | 88 ± 11 |
| SBP (mmHg) | ||||||||
| Men | 117 ± 11 | 114 ± 12 | 120 ± 16 | 135 ± 21 | 119 ± 15 | 126 ± 16 | 134 ± 17 | 130 ± 13 |
| Women | 109 ± 13 | 118 ± 20 | 117 ± 18 | 135 ± 21 | 114 ± 18 | 121 ± 17 | 132 ± 17 | 120 ± 12 |
| DBP (mmHg) | ||||||||
| Men | 78 ± 10 | 73 ± 10 | 74 ± 10 | 72 ± 11 | 72 ± 9 | 80 ± 11 | 81 ± 10 | 80 ± 11 |
| Women | 70 ± 9 | 70 ± 10 | 70 ± 10 | 69 ± 11 | 67 ± 10 | 76 ± 10 | 77 ± 9 | 75 ± 11 |
| % Antihyperlipidemic meds. use | 4 | 81 | 3 | 4 | 10 | 15 | 17 | 0 |
| % Antihypertensive meds. use | 7 | 84 | 20 | 35 | 3 | 22 | 65 | 0.6 |
| % Antiglycemic meds. use‡ | 1.3 | 24 | 3.2 | 5 | 0.2 | 4.3 | 6.9 | 0.6 |
| MetS prevalence (%) | 13.7 | 55.2 | 37.3 | 41.7 | 29.7 | 34.3 | 55.4 | 9.1 |
Values are means ± SD for each quantitative variable and where available split by male/female grouping. Meds., medication.
*Definition of type 2 diabetes: fasting blood glucose >126 mg/dL or antiglycemic medication use.
†Smoking: current smoking.
‡Antiglycemic meds. use: oral hypoglycemic agents or insulin.
SNPs in or near 15 genes were significantly associated with at least one of the 11 traits studied (Fig. 1 and Table 2 and details in Supplementary Tables 1–6). MetS per se was associated with several variants in genes including BUD13 (BUD13 homolog [S. cerevisiae], rs10790162, P = 5.4E-09), ZNF259 (zinc finger protein 259, rs2075290, P = 2.1E-09), and APOA5 (apolipoprotein A-V, rs2266788, P = 1.9E-09), all of which map within the APOA-cluster on chromosome 11 (11q23.3-q23, Supplementary Fig. 2). In addition, variants in LPL (lipoprotein lipase, 8p22, rs295, P = 1.7E-09) and CETP (cholesteryl ester transfer protein, plasma, 16q21, rs173539, P = 9.1E-09) were found associated with MetS. Specifically, variants in LPL were associated with BP-HDLC (rs1441756, P = 2.7E-08), TG-BP (rs15285, P = 1.3E-10), TG-GLUC (rs2197089, P = 1.6E-09), HDLC-TG (rs13702, P = 1.0E-16), and HDLC-WC (rs301, P = 3.2E-11). CETP variants were also significantly associated with BP-HDLC (rs3764261, P = 3.3E-13), HDLC-GLUC (rs9939224, P = 6.9E-12), HDLC-TG (rs173539, P = 4.5E-16), and HDLC-WC (rs173539, P = 1.0E-16), and variants in the gene APOA-cluster (APOA5, BUD13, and ZNF259) were significantly associated with TG-BP, TG-GLUC, HDLC-TG, and WC-TG (Table 2).
FIG. 1.

Genome-wide meta-analyses results. Arrow annotated loci represent genes that show significant associations with MetS and/or individual binary bivariate traits. Each subgraph exemplifies results from a binary bivariate meta-analysis results or MetS meta-analysis. A dashed line in each subgraph represents a significance threshold of negative log10 P value of 7.01 corresponding to a P value of 9.7*10−8 (see Supplementary Data). A gene name in parentheses annotates a variant close to that particular gene. (A high-quality color representation of this figure is available in the online issue.)
TABLE 2.
A summary of STAMPEED β-meta-analyses of the most significant results
| Gene | Variant | Trait | Ch | Position | Meta-β | Meta-SE | Meta P | Homog. P value | Coded | Genotype |
|---|---|---|---|---|---|---|---|---|---|---|
| GCKR | rs780093 | TG-BP | 2 | 27596107 | 0.18 | 0.03 | 3.0E-10 | 0.70 | A | A/G |
| GCKR | rs780093 | WC-TG | 2 | 27596107 | 0.19 | 0.03 | 1.9E-12 | 0.64 | A | A/G |
| C2orf16 | rs1919128 | WC-TG | 2 | 27655263 | −0.18 | 0.03 | 2.0E-09 | 0.82 | A | A/G |
| ZNF512 | rs13022873 | WC-TG | 2 | 27669014 | −0.17 | 0.03 | 5.0E-09 | 0.47 | A | A/C |
| CCDC121 | rs3749147 | WC-TG | 2 | 27705422 | −0.18 | 0.03 | 1.4E-09 | 0.79 | C | C/T |
| ABCB11 | rs569805 | HDLC-GLUC | 2 | 169491126 | 0.16 | 0.03 | 8.5E-08 | 0.46 | A | A/T |
| TFAP2B | rs2206277 | WC-GLUC | 6 | 50906485 | 0.17 | 0.03 | 1.3E-07 | 0.75 | A | A/G |
| (LOC100129150) | rs9987289 | HDLC-TG | 8 | 9220768 | 0.25 | 0.04 | 1.1E-08 | 0.54 | A | A/G |
| (LOC100129150) | rs9987289 | HDLC-WC | 8 | 9220768 | 0.24 | 0.04 | 3.7E-08 | 0.58 | A | A/G |
| LPL | rs295 | MetS | 8 | 19860518 | 0.17 | 0.03 | 1.7E-09 | 0.47 | A | A/C |
| LPL | rs301 | HDLC-WC | 8 | 19861214 | −0.22 | 0.03 | 3.2E-11 | 0.58 | C | C/T |
| LPL | rs13702 | HDLC-TG | 8 | 19868772 | 0.29 | 0.03 | 1.0E-16 | 0.67 | A | A/G |
| LPL | rs15285 | TG-BP | 8 | 19868947 | −0.27 | 0.04 | 1.3E-10 | 0.65 | A | A/G |
| (LPL) | rs2197089 | TG-GLUC | 8 | 19870653 | 0.18 | 0.03 | 1.6E-09 | 1.00 | C | C/T |
| (LPL) | rs1441756 | BP-HDLC | 8 | 19912666 | −0.18 | 0.03 | 2.7E-08 | 0.43 | G | G/T |
| (TRIB1) | rs2954026 | HDLC-TG | 8 | 126553708 | −0.16 | 0.03 | 7.9E-09 | 0.46 | G | G/T |
| (TRIB1) | rs2954033 | TG-BP | 8 | 126562928 | 0.17 | 0.03 | 8.5E-09 | 0.55 | A | A/G |
| (LOC100128354) | rs1387153 | BP-GLUC | 11 | 92313476 | −0.19 | 0.03 | 8.1E-09 | 0.48 | C | C/T |
| (LOC100128354) | rs1387153 | HDLC-GLUC | 11 | 92313476 | −0.21 | 0.03 | 2.4E-09 | 0.49 | C | C/T |
| (LOC100128354) | rs10830956 | TG-GLUC | 11 | 92320661 | −0.20 | 0.03 | 4.8E-11 | 0.67 | C | C/T |
| BUD13 | rs11825181 | TG-BP | 11 | 116131468 | 0.32 | 0.05 | 3.0E-09 | 0.98 | A | A/G |
| BUD13 | rs11820589 | TG-GLUC | 11 | 116139072 | 0.32 | 0.06 | 5.5E-09 | 0.83 | A | A/G |
| BUD13 | rs10790162 | HDLC-TG | 11 | 116144314 | 0.38 | 0.05 | 2.8E-15 | 0.44 | A | A/G |
| BUD13 | rs10790162 | MetS | 11 | 116144314 | 0.25 | 0.04 | 5.4E-09 | 0.44 | A | A/G |
| BUD13 | rs10790162 | WC-TG | 11 | 116144314 | 0.39 | 0.05 | 6.6E-16 | 0.79 | A | A/G |
| ZNF259 | rs11823543 | TG-BP | 11 | 116154345 | 0.35 | 0.06 | 2.5E-09 | 1.00 | A | A/G |
| ZNF259 | rs12286037 | TG-GLUC | 11 | 116157417 | −0.32 | 0.06 | 1.1E-08 | 0.86 | C | C/T |
| ZNF259 | rs2075290 | HDLC-TG | 11 | 116158506 | 0.39 | 0.05 | 1.5E-14 | 0.39 | C | C/T |
| ZNF259 | rs2075290 | MetS | 11 | 116158506 | 0.26 | 0.04 | 2.1E-09 | 0.64 | C | C/T |
| ZNF259 | rs2075290 | WC-TG | 11 | 116158506 | 0.41 | 0.05 | 1.1E-16 | 0.94 | C | C/T |
| APOA5 | rs2266788 | HDLC-TG | 11 | 116165896 | 0.39 | 0.05 | 4.6E-13 | 0.36 | C | C/T |
| APOA5 | rs2266788 | MetS | 11 | 116165896 | 0.26 | 0.04 | 1.9E-09 | 0.66 | C | C/T |
| APOA5 | rs2266788 | TG-BP | 11 | 116165896 | 0.37 | 0.07 | 3.5E-08 | 0.18 | C | C/T |
| APOA5 | rs2266788 | WC-TG | 11 | 116165896 | 0.41 | 0.05 | 2.2E-16 | 0.92 | A | A/G |
| (LIPC) | rs10468017 | HDLC-WC | 15 | 56465804 | 0.16 | 0.03 | 5.5E-08 | 0.47 | C | C/T |
| (LIPC) | rs2043085 | HDLC-GLUC | 15 | 56468246 | −0.17 | 0.03 | 1.3E-08 | 0.83 | A | A/G |
| (CETP) | rs173539 | HDLC-TG | 16 | 55545545 | 0.26 | 0.03 | 4.5E-16 | 0.61 | C | C/T |
| (CETP) | rs173539 | HDLC-WC | 16 | 55545545 | 0.29 | 0.03 | 1.0E-16 | 0.65 | C | C/T |
| (CETP) | rs173539 | MetS | 16 | 55545545 | 0.16 | 0.03 | 9.1E-09 | 0.41 | C | C/T |
| (CETP) | rs3764261 | BP-HDLC | 16 | 55550825 | 0.29 | 0.04 | 3.3E-13 | 0.43 | G | G/T |
| CETP | rs9939224 | HDLC-GLUC | 16 | 55560233 | −0.31 | 0.05 | 6.9E-12 | 0.46 | G | G/T |
| LOC100129500 | rs439401 | HDLC-TG | 19 | 50106291 | 0.24 | 0.04 | 1.0E-08 | 0.44 | C | C/T |
Ch, chromosome number; position, position in base pairs; gene-hugo name, gene name is shown in parentheses when a SNP is near a location of the gene; meta-β, meta-β coefficient; meta-SE, meta-analysis β coefficients SE; homog. P value, a P value from a test of homogeneity of β coefficients, which has a high bound to one when all β coefficients included in the β-meta-analysis are very similar and is significant (less than 0.05) when coefficients among studies differ drastically; coded, the allele that was considered as coded allele for combining data of different studies accounting for β coefficient direction.
The GCKR-cluster of genes on chromosome 2p23–2p23.3 (Supplementary Fig. 3), including GCKR (glucokinase [hexokinase 4] regulator, 2p23), ZNF512 (zinc finger protein 512, 2p23), CCDC121 (coiled-coil domain containing 121, 2p23.3), and C2orf16 (chromosome 2 open reading frame 16, 2p23.3), were, respectively, significantly associated with WC-TG (rs780093, P = 1.9E-12; rs13022873, P = 5.0E-09; rs3749147, P = 1.4E-09; and rs1919128, P = 2.0E-09) and also with TG-BP (rs780093, P = 3.0E-10) for GCKR gene. For a specific genotype in the GCKR variant studied, the genetic additive effects were inversely associated between TG and GLUC.
Additional variants were significantly associated with several binary traits, even if not with MetS. A total of 27 unique variants in 16 genes associated to some bivariate combinations of the five metabolic traits. Variants within LIPC (lipase hepatic, 15q21-q23) associated with HDLC-GLUC, (rs2043085, P = 1.3E-08) and with WC-HDLC (rs10468017, P = 5.5E-08); ABCB11 (ATP-binding cassette, subfamily B [MDR/TAP], member 11, 2q24) associated with HDLC-GLUC (rs569805, P = 8.5E-08); TRIB1 (tribbles homolog 1 [Drosophila], 8q24.13) variants were associated, respectively, with HDLC-TG (rs2954026, P = 7.9E-09) and TG-BP (rs2954033, P = 8.5E-09); and TFAP2B (transcription factor AP-2 β [activating enhancer binding protein 2 β], 6p12) was associated at a borderline significance level with WC-GLUC (rs2206277, P = 1.3E-07). Two SNPs located between LOC100128354 (similar to small nuclear ribonucleoprotein polypeptide G, 11q21) and MTNR1B (melatonin receptor 1B) were significantly associated with BP-GLUC (rs1387153, P = 8.1E-09), with HDLC-GLUC (rs1387153, P = 2.4E-09), and with TG-GLUC (rs10830956, P = 4.8E-11) (Supplementary Fig. 4). In addition, rs439401 of LOC100129500 (protein coding hypothetical LOC100129500, 19q13.2) was associated with HDLC-TG (P = 1.0E-08), and LOC100129150 (protein coding LP5624) variants with HDLC-TG (rs9987289, P = 1.1E-08) and HDLC-WC (rs9987289, P = 3.7E-08) (Fig. 1, Table 2, and details in Supplementary Tables 1–6).
Twenty-nine unique SNPs in association with binary traits and MetS per se were further tested with each of the individual dichotomized traits WC, HDLC, TG, GLUC, and BP (Fig. 2). None of the SNPs demonstrated a significant (P < 0.05) association with three or more traits and none of them with BP, even though some of the SNPs were in fact identified based on their association with MetS. Thus individual dichotomized trait associations do not necessarily reflect what one can discover with the combination of traits. Several variants were associated with two individual dichotomized traits: rs2266788 of APOA5, rs2075290 of ZNF259, rs11820589 of BUD13, and rs13702 of LPL associated with TG and HDLC and rs780093 of GCKR with TG and GLUC. We chose a single SNP with the strongest significance per gene to follow-up with a formal test of pleiotropy using the respective quantitative traits. Figure 3 shows a depiction of the pleiotropic relationships among traits based on the meta-analysis results for the pleiotropy test.
FIG. 2.

Top significant SNPs from meta-analyses of MetS and bivariate traits associations, tested now for their association with dichotomized risk traits (WC, HDLC, TG, GLUC, and BP) as defined in the MetS NCEP definition. The results shown in the graph are the sample weighted mean of negative log10 P values (blue bars) per trait association, for all studies combined. On the top of blue bars (mean) added are the corresponding standard errors (red bars) of these negative log10 P values. The minimal threshold of negative log10 (0.05) P value is shown with a vertical red dashed line. A gene name in parentheses means the corresponding SNP is located in a region near the gene. (A high-quality color representation of this figure is available in the online issue.)
FIG. 3.

A summary of meta-analysis on pleiotropy effects for selected SNPs on pair combinations of quantitative traits. Each study performed a pleiotropy test for selected SNPs with corresponding quantitative trait combinations. The identified meta-significant results show that variants associated more with two lipid measures and fasting glucose. (A high-quality color representation of this figure is available in the online issue.)
Finally, 11 of the top SNPs in 16 genes exhibited association P values with MetS meeting nominal significance levels (meta P < 0.05) (Table 3). Somewhat surprisingly, none of these was associated with fasting insulin levels (results not shown). On average across studies, the top 16 SNPs together accounted for the following proportions of the total variance for each trait: TG, 9%; HDLC, 5.8%; GLUC, 3.6%; WC, 2.3%; and SBP, 1.4%. To gain some insight as to the possible metabolic relationships among these genes, we used the KEGG database (http://www.kegg.jp/) to determine the pathways in which these genes participate. LPL and APOA5 are classified as part of hsa03320 (PPAR signaling) pathway, and LPL and LIPC are part of hsa00561 (Glycerolipid metabolism). In addition, LPL was classified in hsa05010 (Alzheimer’s disease) and LIPC is a member of hsa01100 (Metabolic pathways); MTNR1B is a member of hsa04080 (Neuroactive ligand receptor interaction), and ABCB11 is classified as a member of hsa02010 (ABC transporters) pathway. The rest of reported genes are not classified in pathways at present.
TABLE 3.
Meta-analysis among studies of 16 top SNPs (one per gene) for their association with MetS
| Number | Hugo | SNP | Chrom. | Position (bps) | Meta-β coefficient | Meta-β SE | χ2 value | P value | Homogeneity P value | Number of samples |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | GCKR | rs780093 | 2 | 27596107 | 0.042 | 0.030 | 1.94 | 1.6E-01 | 0.222 | 8 |
| 2 | C2orf16 | rs1919128 | 2 | 27655263 | −0.055 | 0.035 | 2.51 | 1.1E-01 | 0.186 | 8 |
| 3 | ZNF512 | rs13022873 | 2 | 27669014 | −0.054 | 0.036 | 2.24 | 1.3E-01 | 0.154 | 8 |
| 4 | CCDC121 | rs3749147 | 2 | 27705422 | −0.050 | 0.039 | 1.62 | 2.0E-01 | 0.110 | 8 |
| 5 | ABCB11 | rs569805 | 2 | 169491126 | 0.059 | 0.024 | 5.91 | 1.5E-02 | 0.411 | 8 |
| 6 | TFAP2B | rs2206277 | 6 | 50906485 | 0.134 | 0.046 | 8.57 | 3.4E-03 | 0.073 | 6 |
| 7 | LOC100129150 | rs9987289 | 8 | 9220768 | 0.186 | 0.038 | 23.83 | 1.1E-06 | 0.703 | 8 |
| 8 | LPL | rs13702 | 8 | 19868772 | 0.152 | 0.029 | 27.45 | 1.6E-07 | 0.343 | 8 |
| 9 | TRIB1 | rs2954026 | 8 | 126562928 | −0.084 | 0.035 | 5.74 | 1.7E-02 | 0.132 | 8 |
| 10 | LIPC | rs2043085 | 15 | 56468246 | −0.059 | 0.023 | 6.40 | 1.1E-02 | 0.734 | 8 |
| 11 | LOC100128354 | rs10830956 | 11 | 92320661 | −0.070 | 0.031 | 5.03 | 2.5E-02 | 0.254 | 8 |
| 12 | BUD13 | rs10790162 | 11 | 116144314 | 0.243 | 0.060 | 16.65 | 4.5E-05 | 0.172 | 8 |
| 13 | ZNF259 | rs2075290 | 11 | 116158506 | 0.259 | 0.059 | 19.52 | 1.0E-05 | 0.196 | 7 |
| 14 | APOA5 | rs2266788 | 11 | 116165896 | 0.265 | 0.044 | 36.07 | 1.9E-09 | 0.663 | 6 |
| 15 | CETP | rs9939224 | 16 | 55560233 | −0.168 | 0.036 | 21.84 | 3.0E-06 | 0.810 | 8 |
| 16 | LOC100129500 | rs439401 | 19 | 50106291 | 0.053 | 0.056 | 0.89 | 3.5E-01 | 0.071 | 6 |
Number of samples represents study results that were included in the meta-analysis for these SNPs. A few of them were not included in the analysis if they had MAF less than 5%, or Hardy-Weinberg Equilibrium P value less than 10−6, or if the quality of imputation was less than the required thresholds (see research design and methods and Supplementary Data).
DISCUSSION
We used GWAS of pairs of metabolic traits to discover genetic determinants contributing to the correlated architecture of several metabolic traits that define MetS. Angers and Biswas (22), who studied mathematical and statistical aspects of bivariate trait combination versus univariate ordinal categorical data, have shown that univariate analysis fails to detect features of the data found by the bivariate analysis. Therefore, this valid approach has the potential to uncover novel determinants not detectable with usual single phenotype-based analyses. Conversely, it is possible that by bivariate subsampling this study identified top SNPs that may further reduce the association power for any other subsample of three- or four- or five-trait combinations.
A meta-analysis approach was used to augment the power to detect such determinants. DerSimonian and Laird’s method (Supplementary Ref. 18) has the considerable advantage of dynamically accounting for study differences by including the variance of the SNP-wise β parameter estimates in the calculation of the meta-analysis P value. This is particularly valuable in the context of the present analysis, with the variety of ascertainment strategies and ages across individual studies.
We identified 29 unique variants in or near 15 genes associated with binary pairwise traits or with MetS per se at the genome-wide significance level (P < 9.7 × 10−8), and an additional one at borderline significance. Interestingly, all but two of these results included a lipid abnormality, suggesting that genetic effects on lipid levels are more pronounced than for other traits. However, this observation is also consistent with the proposal of McGarry (23) that dyslipidemia is a fundamental component in the development of MetS. Moreover, the predominant pleiotropic pattern was for HDLC and TG, consistent with their well-known inverse correlation (Supplementary Fig. 1). The most influential variants in the correlation among traits were in or near LPL, CETP, APOA5, ZNF259, BUD13, TRIB1, LOC100129500, and LOC100128154 (Table 2). Genes with variants influencing MetS per se included LPL, CETP, and the APOA-cluster (APOA5, ZNF259, and BUD13), which are known to play an important role in lipid metabolism (24–37).
Variants in LPL were significantly associated with MetS and also with binary traits HDLC-BP, TG-BP, TG-GLUC, TG-HDLC, and WC-HDLC. LPL encodes lipoprotein lipase, which hydrolyzes TG in circulating very low-density lipoprotein cholesterol and chylomicrons, providing free fatty acids and monoacylglycerol for utilization by the surrounding target tissues, particularly in skeletal and cardiac muscle and adipose tissue. Previously LPL variants have been associated with individual components of the MetS (14,24,25), as well as with insulin resistance (26) and CHD (27). Our results indicate that LPL has pleiotropic effects on TG and HDLC validated by pleiotropy tests on the quantitative traits (Fig. 3). In Supplementary Fig. 5 we have simulated TG distributions for each study, based on the number of subjects, mean, and standard deviations of TG reported from each study. The LPL variant rs13702 (3′ UTR) had the lowest P value in the association tests within LPL gene, and the prevalent genotype (‘AA’) was associated with increased levels of TG and lower levels of HDLC. We provide detailed directions of association tests for each study in Supplementary Tables 1–6.
CETP is an independent gene that enables the transfer of cholesteryl esters in HDLC toward TG-rich lipoproteins, thereby contributing to lower HDLC. Variants of CETP were significantly associated with MetS, TG-HDLC, HDLC-GLUC, HDLC-BP, and WC-HDLC. Variants in CETP have been associated with HDLC levels (28,29). In our study rs9939224 ‘TT’ rare genotype was associated with higher TG and lower HDLC levels. Although CETP genotypes have been reported extensively in association with CVD, its exact role in disease pathogenesis is unclear (30).
APOA5 is a member of the cluster of genes APOA1–C3–A4–A5, located on 11q23. APOA5 encodes for apolipoprotein A5, a protein found in chylomicrons, VLDL, and HDL particles (31). APOA5 has functional role in the endocytosis of TG-rich particles through its binding to the LDL receptor (32). Variants of the APOA5 gene associate with elevated TG levels and may increase CHD risk (33). APOA5 variants also have been reported to be associated with MetS (34,35). The ‘CC’ genotype of rs2266788 (3′ UTR) was observed to have pleiotropic effects on (increasing) TG and (lowering) HDLC levels. By contrast, the functions of nearby genes ZNF259 and BUD13 are currently unknown, although they may reflect common haplotype associations (Supplementary Fig. 2).
TG-BP and WC-TG were both associated with an intronic SNP, rs780093 in GCKR. The ‘AA’ genotype for rs780093 was associated with higher TG levels and, in most of the studies, with lower levels of GLUC. GCKR also has been previously implicated as a susceptibility gene for type 2 diabetes (36). Functional studies suggest that rs1260326, a nonsynonymous SNP, has an effect on plasma glucose and triglyceride levels through increased glucokinase activity in liver (37). However, this coding SNP is in a different LD block than is rs780093 identified in our study (Supplementary Fig. 3). Furthermore, rs1260326 and the intronic rs780094 within GCKR have been reproducibly associated with multiple quantitative metabolic traits, but in the study of Sparsø et al. (38) with reduced risk for type 2 diabetes. It also is associated with TG (38–40), LDL cholesterol (41), C-reactive protein (42), and WC (43). These reports together with our evidence support the broad pleiotropic effects of GCKR.
LIPC variants demonstrated pleiotropic effects, specifically rs10468017 on WC-HDLC and rs2043085 on HDLC-GLUC. LIPC, which is expressed in liver, has the capacity to catalyze hydrolysis of phospholipids; mono, di-, and triglycerides; and acyl-CoA thioesters and is considered an important enzyme in HDLC metabolism (28,44–46). Our results confirm those of Kathiresan et al. (15), who showed that the minor ‘T’ allele at rs10468017 was associated with lower LIPC expression and increased HDLC levels. The ‘AA’ genotype of rs2043085 also was associated with higher HDLC levels. Variants near TRIB1 (8q24.13) (rs2954026 and rs2954033) associated with TG-BP and TG-HDLC. The rs2954026 and rs2954033 SNPs locate 34 and 43 Kb downstream of TRIB1, respectively, a gene that has been associated with hyperlipoproteinemia (47). TRIB1 and TRIB2 genes also have been found to be upregulated in regions of human atherosclerotic plaque.
Other interesting variants are rs10830956 and rs13887153, which map 10 and 3 Kb downstream of LOC100128354, respectively, and about 22 and 29 Kb upstream of MTNR1B. These were associated with TG-GLUC and BP-GLUC, HDLC-GLUC. These variants also showed highly significant associations with fasting glucose levels (Fig. 2). The less common ‘TT’ genotype of rs10830956 showed higher levels of TG and GLUC. These variants are in LD with the MTNR1B gene, although they reside in an LD block physically closer to the LOC1001128354 gene (Supplementary Fig. 4). A recent large study with more than 36,000 participants of European descent showed that rs13887153 associated with glucose. They reported that each copy of the rs13887153 ‘G’ allele was associated with increased levels of glucose, reduced β-cell function measured by HOMA-B, and an increased risk for type 2 diabetes (48). Similar findings for fasting glucose were described by others (49). Our findings support the hypothesis that variants rs10830956 and rs13887153 may serve as proxies for variants in nearby genes LOC100128354/MTNR1B (melatonin receptor) involved in insulin/glucose metabolism.
Several variants were associated with only a single bivariate trait. The rs569805 in the ABCB11 significantly associated with HDLC-GLUC. This intronic SNP is located within 20 Kb and is in LD (r2 = 0.67) with intronic SNP rs560887 in G6PC2, the most significant variant in association with fasting GLUC (P = 4E-75) in a meta-analysis conducted by the MAGIC consortium (36). By contrast, rs560887 was not associated with HDLC levels in the ENGAGE consortium (21). Another significant association for TG-HDLC was identified on chromosome 19, with rs439401, in an intron of the hypothetical protein LOC100129500. This SNP demonstrated association (P = 2E-9) with TG levels and nominal association (P = 3E-3) with HDLC levels in a recent meta-analysis of GWA studies in 16 European cohorts (21). This SNP is in APOC1 (with relative position of −3.7 Kb) located within the larger APOE/C1/C4/C2 gene cluster, known to have pleiotropic effects on lipid metabolism (50). A number of our lipid findings are also reported by Teslovich et al. (20) in a GWAS meta-analysis for plasma lipid traits with more than 123,000 subjects. In that study, 26 out of the 95 loci associated with more than one lipid trait at genome-wide significance.
We set out to test the hypothesis that common variants explain the correlated architecture among MetS traits, at least in part. These common top SNPs (16 variants selected one per gene) together explained a total average variance ranging from 1.4% for SBP to 9.0% for TG. The weak effects on SBP are consistent with a large GWAS on BP with more than 84,000 participants, able to explain less than 2% of BP variation (17). Explicit tests of pleiotropy of our study’s top SNPs on the relevant quantitative traits validated 11 pleiotropic relationships summarized in Fig. 3. Because they explained a small fraction of the correlation among metabolic syndrome traits, other factors must also be at play: variants with rare alleles; nonadditive effects, such as dominance and epistasis; furthermore, cascade effects of a phenotype, e.g., obesity directly influencing glycemic, lipid and blood pressure traits; as well as common environmental factors in defining the correlated architecture of these traits. For tests of pleiotropy carried out on the quantitative phenotypes, we imputed lipid- and SBP trait values for treated subjects based on summaries of a large number of clinical trials. However, we acknowledge that this approach is not ideal and may have limited our ability to detect true pleiotropic effects.
We explored also the relationships among the genes identified in this study. A hypothesized network of interactions among identified genes was constructed using GeneGO software (Fig. 4). Five of the selected genes interact with many genes, represented in Fig. 4 as hidden links. Even this consideration suggests the possibility that many other genes may act in the context of their respective pathways, rather than independently. For example, LPL interacts with INS, APOE, APOB, APOA1, APOA4, APOC3, APOC4, LRP1, and NETO1; and CETP has at least 35 interactions, based on Sigma-Aldrich and Ingenuity database (http://www.sigma-aldrich.com/yfg). It is possible that cross-talk among these pathways via intermediate activator/suppressor molecules contributes to the clustering of disorders in MetS. Thus one possible fruitful direction of study is to examine MetS and its correlated structure in a systems biology context. In summary, none of these variants associated simultaneously with most or all the traits, supporting the concept that MetS is not a consequence of any single pathway or factor but rather a consequence of interactions among different pathways.
FIG. 4.

One of the simplified networks of the genes LPL, CETP, APOA5, GCKR, LIPC, and ABCB11 among 16 genes reported in this article with variants significantly associated to MetS and/or its bivariate traits. This network was built from curated publications based on GeneGO database. (Green arrows show activation; red arrows show suppression.) (A high-quality color representation of this figure is available in the online issue.)
ACKNOWLEDGMENTS
This collaborative work was supported in part by the National Institutes of Health (NIH) grants STAMPEED HL-087700, ADVANCE 5R01-HL-087647, ARIC U01-HL-075572, CHS HL-087652, FHS HL-0877700, GeneSTAR 5R01-HL-087698, GENOA HL-087660, and NFBC1RL1-MH-083268-01.
The ARIC Study is carried out as a collaborative study supported by NHLBI contracts N01-HC-55015, N01-HC-55016, N01-HC-55018, N01-HC-55019, N01-HC-55020, N01-HC-55021, N01-HC-55022, R01-HL-087641, R01-HL-59367, and R01-HL-086694; National Human Genome Research Institute contract U01HG004402; and NIH contract HHSN268200625226C. Infrastructure was partly supported by Grant Number UL1-RR-025005, a component of the NIH and NIH Roadmap for Medical Research.
The CHS research reported in this article was supported by contract numbers N01-HC-85079 through N01-HC-85086, N01-HC-35129, N01-HC-15103, N01-HC-55222, N01-HC-75150, N01-HC-45133, grant numbers U01-HL-080295 and R01-HL-087652 from the NHLBI, with additional contribution from the National Institute of Neurologic Disorders and Stroke. A full list of principal CHS investigators and institutions can be found at http://www.chs-nhlbi.org/pi.htm. DNA handling and genotyping was supported in part by National Center for Research Resources Grant M01RR00425 to the Cedars-Sinai General Clinical Research Center Genotyping core and National Institute of Diabetes and Digestive and Kidney Diseases Grant DK-063491 to the Southern California Diabetes Endocrinology Research Center.
The NFBC 1966 study received financial support from the Academy of Finland (project grants 104781, 120315, and Center of Excellence in Complex Disease Genetics); University Hospital Oulu, Biocenter; University of Oulu, Finland; the European Commission (EURO-BLCS, Framework 5 award QLG1-CT-2000-01643); NHLBI Grant 5R01-HL-087679-02 through the STAMPEED program (1RL1MH083268-01); NIH/NIMH (5R01-MH-63706:02); ENGAGE project and grant agreement HEALTH-F4-2007-201413; and the Medical Research Council (studentship grant G0500539). The DNA extractions, sample quality controls, biobank upkeeping, and aliquoting were performed in the National Public Health Institute, Biomedicum Helsinki, Finland and supported financially by the Academy of Finland and Biocentrum Helsinki.
No potential conflicts of interest relevant to this article were reported.
A.T.K., D.V., and J.S.P. wrote the article, researched data, and contributed to discussion. M.O.G. wrote the article and contributed to discussion. T.L.A. wrote the article, researched data, and contributed to discussion. I.J.K. wrote the article and contributed to discussion. U.S., R.A.M., Y.V.S., and N.F. researched data and contributed to discussion. D.A., G.L., and Q.Z. researched data. M.F.F. researched data and contributed to discussion. N.L.G., T.H., and A.-L.H. researched data. J.W.K. researched data and contributed to discussion. K.E.N., C.I., B.K., and L.Y. researched data. P.F.O., M.I.M., C.J., D.J.C., A.C., B.M.P., L.C.B., M.A.P., E.B., and T.Q. contributed to discussion. L.P. researched data. M.-R.J. wrote the article and contributed to discussion. D.M.B. and S.L.R.K. contributed to discussion. J.I.R., Y.-D.I.C., and I.B.B. wrote the article and contributed to discussion. All authors reviewed and edited the article.
The authors thank the staff and participants of each study for important contributions. Recently the authors lost a distinguished colleague and coauthor Leena Palotie. The authors acknowledge her major contributions in human genetics.
Footnotes
D.V., J.I.R., Y.-D.I.C., and I.B.B. are from the STAMPEED MetS Steering Committee.
Deceased.
This article contains Supplementary Data online at http://diabetes.diabetesjournals.org/lookup/suppl/doi:10.2337/db10-1011/-/DC1.
REFERENCES
- 1.Third report of the National Cholesterol Education Program (NCEP) expert panel on detection, evaluation and treatment of high blood cholesterol in adults (Adult Treatment Panel III). Final report. Circulation 2002;106:3144–3421 [PubMed] [Google Scholar]
- 2.Ford ES, Giles WH, Dietz WH. Prevalence of the metabolic syndrome among US adults: findings from the third National Health and Nutrition Examination Survey. JAMA 2002;287:356–359 [DOI] [PubMed] [Google Scholar]
- 3.Ilanne-Parikka P, Eriksson JG, Lindström J, et al. Finnish Diabetes Prevention Study Group Prevalence of the metabolic syndrome and its components: findings from a Finnish general population sample and the Diabetes Prevention Study cohort. Diabetes Care 2004;27:2135–2140 [DOI] [PubMed] [Google Scholar]
- 4.Iribarren C, Go AS, Husson G, et al. Metabolic syndrome and early-onset coronary artery disease: is the whole greater than its parts? J Am Coll Cardiol 2006;48:1800–1807 [DOI] [PubMed] [Google Scholar]
- 5.Kraja AT, Province MA, Huang P, et al. Trends in metabolic syndrome and gene networks in human and rodent models. Endocr Metab Immune Disord Drug Targets 2008;8:198–207 [DOI] [PubMed] [Google Scholar]
- 6.Martins D, Tareen N, Ogedegbe G, Pan D, Norris K. The relative risk of cardiovascular death among racial and ethnic minorities with metabolic syndrome: data from the NHANES-II mortality follow-up. J Natl Med Assoc 2008;100:565–571 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hanley AJ, Karter AJ, Festa A, et al. Insulin Resistance Atherosclerosis Study Factor analysis of metabolic syndrome using directly measured insulin sensitivity: The Insulin Resistance Atherosclerosis Study. Diabetes 2002;51:2642–2647 [DOI] [PubMed] [Google Scholar]
- 8.Kraja AT, Rao DC, Weder AB, et al. Two major QTLs and several others relate to factors of metabolic syndrome in the family blood pressure program. Hypertension 2005;46:751–757 [DOI] [PubMed] [Google Scholar]
- 9.Vaidya D, Szklo M, Liu K, Schreiner PJ, Bertoni AG, Ouyang P. Defining the metabolic syndrome construct: Multi-Ethnic Study of Atherosclerosis (MESA) cross-sectional analysis. Diabetes Care 2007;30:2086–2090 [DOI] [PubMed] [Google Scholar]
- 10.Lehman DM, Arya R, Blangero J, et al. Bivariate linkage analysis of the insulin resistance syndrome phenotypes on chromosome 7q. Hum Biol 2005;77:231–246 [DOI] [PubMed] [Google Scholar]
- 11.Chiu YF, Chuang LM, Kao HY, et al. Bivariate genome-wide scan for metabolic phenotypes in non-diabetic Chinese individuals from the Stanford, Asia and Pacific Program of Hypertension and Insulin Resistance Family Study. Diabetologia 2007;50:1631–1640 [DOI] [PubMed] [Google Scholar]
- 12.Kullo IJ, de Andrade M, Boerwinkle E, McConnell JP, Kardia SL, Turner ST. Pleiotropic genetic effects contribute to the correlation between HDL cholesterol, triglycerides, and LDL particle size in hypertensive sibships. Am J Hypertens 2005;18:99–103 [DOI] [PubMed] [Google Scholar]
- 13.Edwards KL, Mahaney MC, Motulsky AG, Austin MA. Pleiotropic genetic effects on LDL size, plasma triglyceride, and HDL cholesterol in families. Arterioscler Thromb Vasc Biol 1999;19:2456–2464 [DOI] [PubMed] [Google Scholar]
- 14.Bauer F, Elbers CC, Adan RA, et al. Obesity genes identified in genome-wide association studies are associated with adiposity measures and potentially with nutrient-specific food preference. Am J Clin Nutr 2009;90:951–959 [DOI] [PubMed] [Google Scholar]
- 15.Kathiresan S, Willer CJ, Peloso GM, et al. Common variants at 30 loci contribute to polygenic dyslipidemia. Nat Genet 2009;41:56–65 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Lindgren CM, Heid IM, Randall JC, et al. Wellcome Trust Case Control Consortium. Procardis Consortia. Giant Consortium Genome-wide association scan meta-analysis identifies three Loci influencing adiposity and fat distribution. PLoS Genet 2009;5:e1000508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Newton-Cheh C, Johnson T, Gateva V, et al. Wellcome Trust Case Control Consortium Genome-wide association study identifies eight loci associated with blood pressure. Nat Genet 2009;41:666–676 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sabatti C, Service SK, Hartikainen AL, et al. Genome-wide association analysis of metabolic traits in a birth cohort from a founder population. Nat Genet 2009;41:35–46 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Willer CJ, Speliotes EK, Loos RJ, et al. Wellcome Trust Case Control Consortium. Genetic Investigation of Anthropometric Traits Consortium Six new loci associated with body mass index highlight a neuronal influence on body weight regulation. Nat Genet 2009;41:25–34 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Teslovich TM, Musunuru K, Smith AV, et al. Biological, clinical and population relevance of 95 loci for blood lipids. Nature 2010;466:707–713 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Aulchenko YS, Ripatti S, Lindqvist I, et al. ENGAGE Consortium Loci influencing lipid levels and coronary heart disease risk in 16 European population cohorts. Nat Genet 2009;41:47–55 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Angers J, Biswas A. Bivariate versus univariate ordinal categorical data with reference to an ophthalmologic study. J Stat Comput Simul 2008;78:489–502 [Google Scholar]
- 23.McGarry JD. Banting lecture 2001: dysregulation of fatty acid metabolism in the etiology of type 2 diabetes. Diabetes 2002;51:7–18 [DOI] [PubMed] [Google Scholar]
- 24.Mead JR, Ramji DP. The pivotal role of lipoprotein lipase in atherosclerosis. Cardiovasc Res 2002;55:261–269 [DOI] [PubMed] [Google Scholar]
- 25.Goodarzi MO, Wong H, Quiñones MJ, et al. The 3′ untranslated region of the lipoprotein lipase gene: haplotype structure and association with post-heparin plasma lipase activity. J Clin Endocrinol Metab 2005;90:4816–4823 [DOI] [PubMed] [Google Scholar]
- 26.Goodarzi MO, Guo X, Taylor KD, et al. Lipoprotein lipase is a gene for insulin resistance in Mexican Americans. Diabetes 2004;53:214–220 [DOI] [PubMed] [Google Scholar]
- 27.Goodarzi MO, Guo X, Taylor KD, et al. Determination and use of haplotypes: ethnic comparison and association of the lipoprotein lipase gene and coronary artery disease in Mexican-Americans. Genet Med 2003;5:322–327 [DOI] [PubMed] [Google Scholar]
- 28.Chasman DI, Paré G, Zee RY, et al. Genetic loci associated with plasma concentration of LDL-C, HDL-C, triglycerides, ApoA1, and ApoB among 6382 Caucasian women in genome-wide analysis with replication. Circ Cardiovasc Genet 2008;1:21–30 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Agerholm-Larsen B, Tybjaerg-Hansen A, Schnohr P, Steffensen R, Nordestgaard BG. Common cholesteryl ester transfer protein mutations, decreased HDL cholesterol, and possible decreased risk of ischemic heart disease: The Copenhagen City Heart Study. Circulation 2000;102:2197–2203 [DOI] [PubMed] [Google Scholar]
- 30.Cannon CP, Shah S, Dansky HM, et al. the DEFINE Investigators Safety of Anacetrapib in patients with or at high risk for coronary heart disease. N Engl J Med 2010;363:2406–2415 [DOI] [PubMed] [Google Scholar]
- 31.O’Brien PJ, Alborn WE, Sloan JH, et al. The novel apolipoprotein A5 is present in human serum, is associated with VLDL, HDL, and chylomicrons, and circulates at very low concentrations compared with other apolipoproteins. Clin Chem 2005;51:351–359 [DOI] [PubMed] [Google Scholar]
- 32.Nilsson SK, Christensen S, Raarup MK, Ryan RO, Nielsen MS, Olivecrona G. Endocytosis of apolipoprotein A-V by members of the low density lipoprotein receptor and the VPS10p domain receptor families. J Biol Chem 2008;283:25920–25927 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Smith CE, Tucker KL, Lai CQ, Parnell LD, Lee YC, Ordovás JM. Apolipoprotein A5 and lipoprotein lipase interact to modulate anthropometric measures in Hispanics of Caribbean origin. Obesity (Silver Spring) 2010;18:327–332 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Grallert H, Sedlmeier EM, Huth C, et al. APOA5 variants and metabolic syndrome in Caucasians. J Lipid Res 2007;48:2614–2621 [DOI] [PubMed] [Google Scholar]
- 35.Hsu LA, Ko YL, Chang CJ, Teng MS, Wu S, Hu CF. Apolipoprotein A5 gene -1131T/C polymorphism is associated with the risk of metabolic syndrome in ethnic Chinese in Taiwan. Clin Chem Lab Med 2008;46:1714–1719 [DOI] [PubMed] [Google Scholar]
- 36.Dupuis J, Langenberg C, Prokopenko I, et al. DIAGRAM Consortium. GIANT Consortium. Global BPgen Consortium. Anders Hamsten on behalf of Procardis Consortium. MAGIC investigators New genetic loci implicated in fasting glucose homeostasis and their impact on type 2 diabetes risk. Nat Genet 2010;42:105–116 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Beer NL, Tribble ND, McCulloch LJ, et al. The P446L variant in GCKR associated with fasting plasma glucose and triglyceride levels exerts its effect through increased glucokinase activity in liver. Hum Mol Genet 2009;18:4081–4088 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Sparsø T, Andersen G, Nielsen T, et al. The GCKR rs780094 polymorphism is associated with elevated fasting serum triacylglycerol, reduced fasting and OGTT-related insulinaemia, and reduced risk of type 2 diabetes. Diabetologia 2008;51:70–75 [DOI] [PubMed] [Google Scholar]
- 39.Willer CJ, Sanna S, Jackson AU, et al. Newly identified loci that influence lipid concentrations and risk of coronary artery disease. Nat Genet 2008;40:161–169 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Wallace C, Newhouse SJ, Braund P, et al. Genome-wide association study identifies genes for biomarkers of cardiovascular disease: serum urate and dyslipidemia. Am J Hum Genet 2008;82:139–149 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Kathiresan S, Melander O, Guiducci C, et al. Six new loci associated with blood low-density lipoprotein cholesterol, high-density lipoprotein cholesterol or triglycerides in humans. Nat Genet 2008;40:189–197 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Ridker PM, Pare G, Parker A, et al. Loci related to metabolic-syndrome pathways including LEPR,HNF1A, IL6R, and GCKR associate with plasma C-reactive protein: the Women’s Genome Health Study. Am J Hum Genet 2008;82:1185–1192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.van der Harst P, Bakker SJ, de Boer RA, et al. Replication of the five novel loci for uric acid concentrations and potential mediating mechanisms. Hum Mol Genet 2010;19:387–395 [DOI] [PubMed] [Google Scholar]
- 44.Rufibach LE, Duncan SA, Battle M, Deeb SS. Transcriptional regulation of the human hepatic lipase (LIPC) gene promoter. J Lipid Res 2006l;47:1463–1477 [DOI] [PubMed] [Google Scholar]
- 45.Feitosa MF, Myers RH, Pankow JS, Province MA, Borecki IB. LIPC variants in the promoter and intron 1 modify HDL-C levels in a sex-specific fashion. Atherosclerosis 2009;204:171–177 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Freeman L, Amar MJ, Shamburek R, et al. Lipolytic and ligand-binding functions of hepatic lipase protect against atherosclerosis in LDL receptor-deficient mice. J Lipid Res 2007;48:104–113 [DOI] [PubMed] [Google Scholar]
- 47.Deng J, James CH, Patel L, et al. Human tribbles homologue 2 is expressed in unstable regions of carotid plaques and regulates macrophage IL-10 in vitro. Clin Sci (Lond) 2009;116:241–248 [DOI] [PubMed] [Google Scholar]
- 48.Bouatia-Naji N, Bonnefond A, Cavalcanti-Proença C, et al. A variant near MTNR1B is associated with increased fasting plasma glucose levels and type 2 diabetes risk. Nat Genet 2009;41:89–94 [DOI] [PubMed] [Google Scholar]
- 49.Lyssenko V, Nagorny CL, Erdos MR, et al. Common variant in MTNR1B associated with increased risk of type 2 diabetes and impaired early insulin secretion. Nat Genet 2009;41:82–88 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Klos K, Shimmin L, Ballantyne C, et al. APOE/C1/C4/C2 hepatic control region polymorphism influences plasma apoE and LDL cholesterol levels. Hum Mol Genet 2008;17:2039–2046 [DOI] [PMC free article] [PubMed] [Google Scholar]